
Abstract
The concomitant infections of Canine distemper virus (CDV), Canine adenovirus A types 1 (CAdV-1) and 2 (CAdV-2), Canine parvovirus type 2 (CPV-2), and Toxoplasma gondii are described in a 43-day-old mixed-breed puppy. Clinically, there were convulsions and blindness with spontaneous death; 14 siblings of this puppy, born to a 10-month-old dam, which was seropositive (titer: 1,024) for T. gondii, also died. Necropsy revealed unilateral corneal edema (blue eye), depletion of intestinal lymphoid tissue, non-collapsible lungs, congestion of meningeal vessels, and a pale area in the myocardium. Histopathology demonstrated necrotizing myocarditis associated with intralesional apicomplexan protozoa; necrotizing and chronic hepatitis associated with rare intranuclear inclusion bodies within hepatocytes; necrotizing bronchitis and bronchiolitis; interstitial pneumonia associated with eosinophilic intracytoplasmic inclusion bodies within epithelial cells; atrophy and fusion of intestinal villi with cryptal necrosis; and white matter demyelination of the cerebrum and cerebellum associated with intranuclear inclusion bodies within astrocytes. Polymerase chain reaction (PCR) amplified the partial fragments (bp) of the CDV N gene (290 bp), CPV-2c VP2 capsid protein gene (583 bp), and CAdV-1 (508 bp) and CAdV-2 (1,030 bp) E gene from urine and tissue samples. The PCR assays demonstrated that the apicomplexan protozoa observed within several organs contained DNA specific for T. gondii; genotyping revealed T. gondii type III. The findings support the characterization of concomitant infections of CDV, CAdV-1, CAdV-2, CPV-2, and T. gondii in this puppy. Further, seroreactivity to T. gondii of the dam in association with the systemic disease observed in the puppy described herein is suggestive of congenital toxoplasmosis.
Canine distemper virus (CDV; order Mononegavirales, family Paramyxoviridae, subfamily Paramyxovirinae, genus Morbillivirus) causes systemic, respiratory, cutaneous, bone, and/or neurological manifestations in a wide range of mammalian hosts. 14 Although the occurrence of canine distemper (CD) is significantly reduced in domestic dog populations in developed countries due to the use of vaccination, 14 this disease is endemic and is a major cause of mortality in Brazil, impacting the local economy with an estimated US$147.5–$160.3 million annually due to the therapy associated with the systemic effects of CDV. 16 Infectious canine hepatitis (ICH), caused by Canine adenovirus A (family Adenoviridae, genus Mastadenovirus) type 1 (CAdV-1), occurs worldwide, is more frequently observed in dogs less than 1 year of age, and is clinically characterized by corneal edema. 5 Canine adenovirus A type 2 (CAdV-2) has been consistently recovered from dogs that have suffered from the clinical entity known as canine infectious respiratory disease (CIRD), kennel cough, or canine infectious tracheobronchitis. 10 Canine parvoviral enteritis is a fatal gastrointestinal disease that is caused by Canine parvovirus type 2 (CPV-2) and predominantly affects puppies that are between 6 weeks and 6 months of age. 13 Toxoplasmosis is caused by the protozoa Toxoplasma gondii, which is frequently associated with secondary infections in dogs. 9
Coinfections of CDV with CAdV-1, 8 CAdV-2, 7 Canine parainfluenza virus (CPIV), 8 and CPV-2 enteritis have been described, and there are reports of triple viral infections (CDV, CPIV, and CAdV-2). 8 The current study describes the findings of systemic canine distemper, infectious tracheobronchitis, and toxoplasmosis with simultaneous ICH and canine parvoviral enteritis in a puppy.
A 43-day-old, male, mixed-breed dog was submitted for routine necropsy to the Laboratory of Veterinary Pathology, Universidade Norte do Paraná, southern Brazil. The puppy was part of a litter of 15 (7 males and 8 females); the puppies were born from a 10-month-old, unvaccinated, female Pit Bull mix and were kept in a country house within the city of Londrina, southern Brazil. The dam originally belonged to a colony of dogs, and reportedly demonstrated clinical manifestations of distemper at approximately 60 days of age. Additionally, during the last phase of pregnancy, the dam had sporadic contacts with 2 Lhasa Apso dogs that had ocular-nasal secretions. According to the referring veterinarian, 13 of the puppies died suddenly during a period of 30 days. The first puppy died 15 days after birth, demonstrating ocular discharge, with death of the other 14 puppies occurring in intervals of 3–4 days; 7 puppies had similar ocular lesions. The last of the 14 puppies had episodes of convulsion before spontaneous death. The puppy submitted for necropsy had reportedly demonstrated convulsive crises and manifestations of blindness; dexamethasone, penicillin, vitamin B1, and poly-vitamin complex were prescribed, and the puppy apparently recovered from the illness. However, convulsions, stumbling into objects, and discrete icterus of the ocular mucosa were observed 2 days later before the animal died spontaneously, after which a routine necropsy was done. At necropsy, the puppy was afebrile, without ocular or nasal secretions.
Selected tissue samples (brain, lung, liver, kidneys, palatine tonsils, heart, urinary bladder, intestine, spleen, and lymph nodes) were fixed by immersion in 10% buffered formalin solution and routinely processed for histopathological evaluation. Fresh fecal samples, collected during necropsy, were submitted for parasitological analyses by the zinc sulfate flotation technique.
Serum samples (collected in ethylenediamine tetra-acetic acid tubes) from the dam were used for the detection of anti–T. gondii antibodies by indirect fluorescent antibody test (IFAT) 11 ; a reaction was considered positive when a titer measuring ≥16 was obtained. Selected formalin-fixed, paraffin-embedded (FFPE) tissue sections of the liver and lung were processed for the detection of CAdV-1 and CAdV-2 nucleic acid, using in situ hybridization (ISH). 3
Urine and selected tissue samples (brain, liver, spleen, lung, and cervical lymph node) were processed for extraction of RNA and subsequent reverse transcription polymerase chain reaction (RT-PCR) assay targeting a 287–base pair (bp) fragment of the N gene of CDV. 15 The partial nucleotide sequences of the CDV N gene obtained were initially compared with those deposited in GenBank by using BLAST (http://www.ncbi.nlm.nih.gov/BLAST). Phylogenetic trees and sequence alignments were then created by using MEGA 5, 24 constructed by the Maximum Likelihood method, based on 1,000 bootstrapped data sets. Sequences of Phocine distemper virus were included as an out-group to provide stability to the generated tree. The GenBank accession numbers of the nucleotide sequences used for phylogenetic analyses are given in Figure 1.
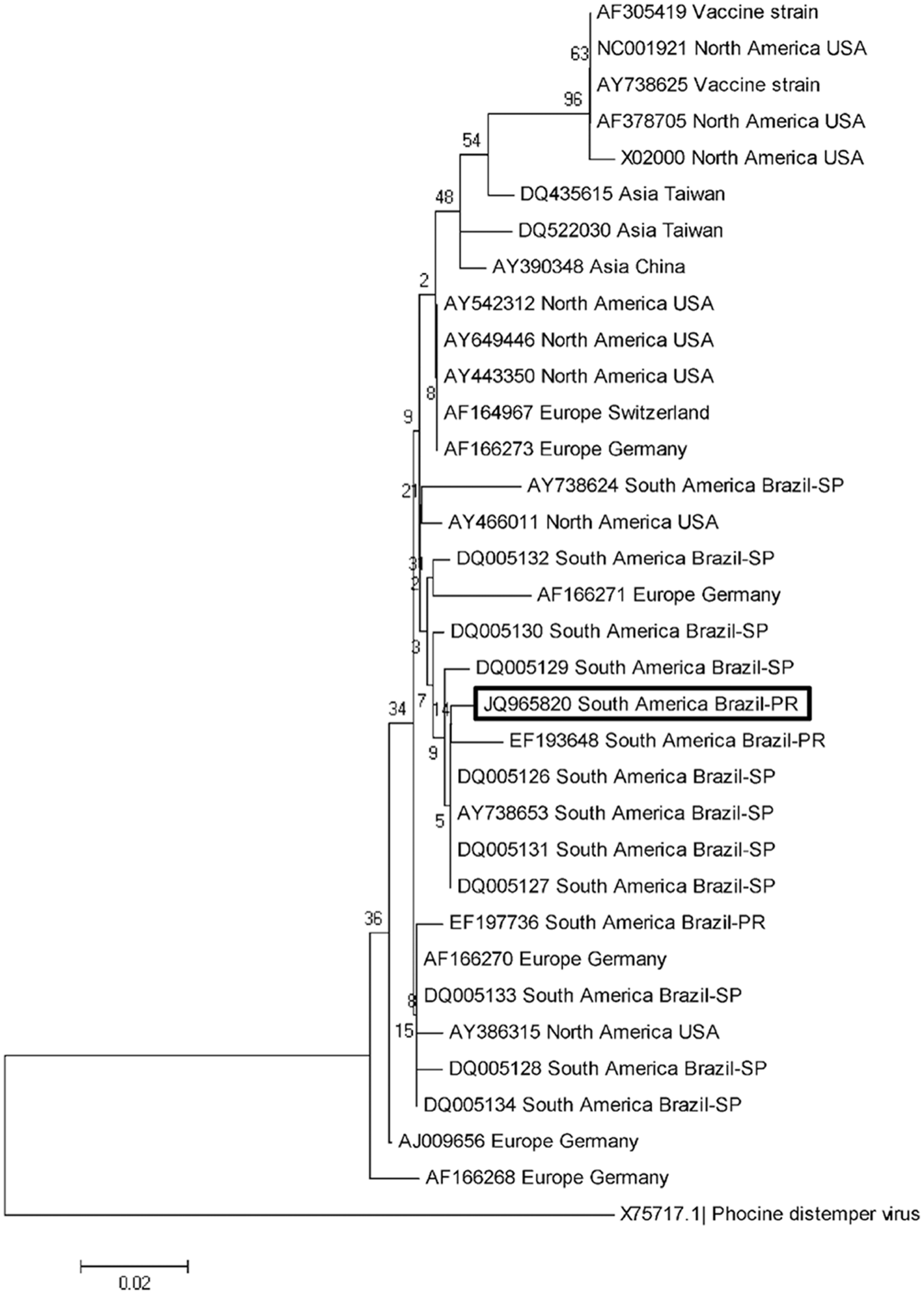
The phylogenetic relationship of selected strains of the Canine distemper virus N gene generated by MEGA 5. 24 The bootstrap consensus tree was constructed by the Maximum Likelihood method using nucleotide sequences with 1,000 bootstrap replicates and was based on the Tamura–Nei model. The GenBank accession numbers and the geographical origin of the sequences used are given. The sequence derived from the current study is highlighted within the box. Phocine distemper virus was used as the out-group. PR = Paraná; SP = São Paulo.
The amplification of CAdV-1 and -2 viral DNA was performed from freshly collected tissue samples (liver and kidney) and urine by using a PCR assay designed to detect the 508-bp and 1,030-bp fragments of the E gene of CAdV-1 and -2, respectively. 19 DNA extracted from the thymus, cervical lymph node, and intestine was used to amplify the 583-bp fragment of the VP-2 gene of CPV-2c. 18 DNA from fresh tissue fragments of the liver, kidney, and lung was used for the amplification of the 450-bp fragment of the glycoprotein B gene of Canid herpesvirus 1 (CaHV-1). 22 Positive controls consisted of aliquots from commercial vaccines (CAdV-1, 2, and CPV-2), and from previously described (CDV) 15 and unpublished (CaHV-1; Headley SA, et al., manuscript in preparation) cases; nuclease-free water a served as negative control. All obtained PCR products were separated by electrophoresis in 2% agarose gels, stained with ethidium bromide, and examined under ultraviolet light. The products of the CDV RT-PCR assay were purified b and submitted for direct sequencing using the forward and reverse primers.
Selected FFPE tissue blocks (liver and myocardium [with and without white foci] and lungs) were used to identify the presence of apicomplexan protozoa by extracting protozoan DNA using a commercial kit. c The extracted DNA was then used in a PCR assay that targets the 18S ribosomal DNA (rDNA) gene that is common between apicomplexan protozoa, by using specific primers (Tg18s58F: CTAAGTATAA GCTTTTATACGGC; Tg18s348R: TGCCACGGTAGTCCA ATAC) that amplify a 291-bp fragment of T. gondii. 23 Genetic typing of the apicomplexan protozoa was then achieved by restriction enzymatic digestion to differentiate between T. gondii, Neospora caninum, Hammondia hammondi, and Sarcocystis neurona by conventional PCR–restriction fragment length polymorphism (PCR-RFLP). 23 Positive controls consisted of protozoan DNA from previous cases; ultrapure water a was used as negative control. Positive and negative controls were included in each assay. Genotyping was performed by using multilocus PCR-RFLP analyses of 13 markers: SAG2-3’, SAG2-5’, SAG3, GRA6, SAG1, BTUB, L358, c22-8, c29-2, PK1, Apico, SAG2-alt, and CS3. 23
Necropsy revealed discrete icterus of visible mucous membranes and the subcutaneous tissue and corneal edema of the left eye (Fig. 2A). The lungs were non-collapsible, wet, and heavy, with discrete accumulation of edematous fluid at the distal extremity of the trachea. There was a focal (2 mm × 4 mm) white streak at the myocardium, close to the apex (Fig. 2B). Lymph nodes (cervical, axillary, prescapular, inguinal, and mesenteric) were moderately enlarged and edematous; the palatine tonsils were also enlarged. The serosal surface of the intestine was slightly granular, and Peyer patches were easily identified; the intestinal lumen was filled with a predominantly watery content, and there was marked depletion of Peyer patches. The liver was pale and friable, the lobular pattern evident with pale foci randomly distributed throughout the capsular surface. Additionally, there was moderate congestion of meningeal vessels.
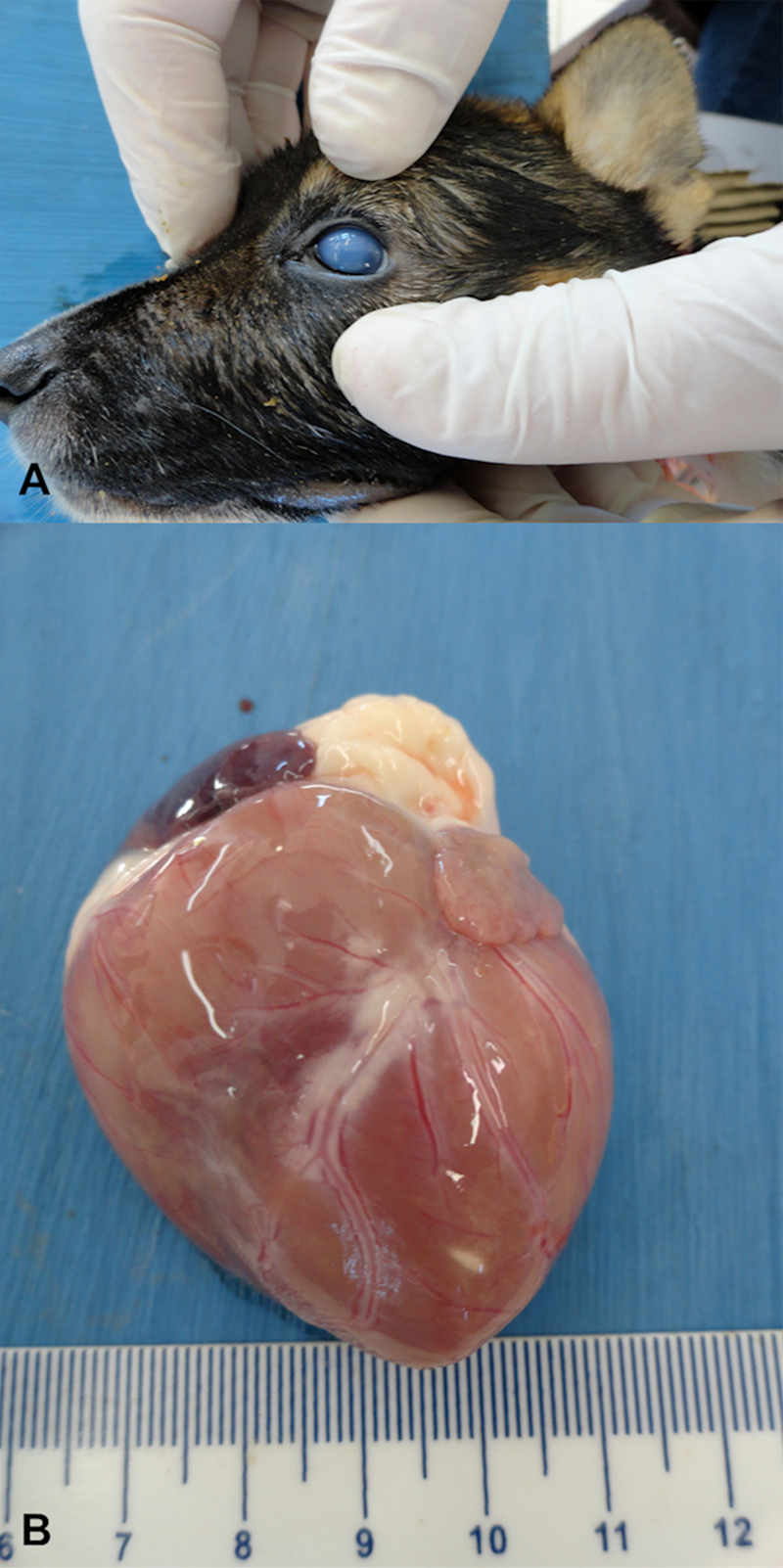
Gross images; 43-day-old, mixed-breed dog. Observe the blue eye feature that is characteristic of canine infectious hepatitis (
Significant histopathological alterations were observed within most tissues collected during necropsy. Pulmonary alterations included interstitial pneumonia associated with intracytoplasmic eosinophilic inclusion bodies within bronchial and bronchiolar epithelial cells (Fig. 3A). In some areas of interstitial pneumonia, there was destruction of the alveolar, bronchial, and bronchiolar epithelium and vasculitis associated with intralesional protozoa (Fig. 3B). There was necrohemorrhagic hepatitis associated with large intranuclear amphophilic inclusion bodies predominantly within Kupffer cells, and more sporadically in viable hepatocytes (Fig. 3C). Additionally, there were areas of random coagulative hepatocellular necrosis associated with intralesional apicomplexan protozoan (Fig. 3D). The white foci observed grossly at the myocardium demonstrated severe necrotizing myocarditis with dystrophic mineralization associated with large quantities of intralesional protozoa (Fig. 3E); these were also observed in areas of myocardial degeneration and nonsuppurative myocarditis. There was discrete white matter demyelination and nonsuppurative meningoencephalitis of the cerebrum and cerebellum associated with few eosinophilic inclusion bodies within astrocytes. Within the intestine, there was atrophy and fusion of intestinal villi with cryptal necrosis (Fig. 3F), depletion of lymphoid tissue, regeneration of cryptal epithelial cells, and discrete hemorrhage. Further, there was nonsuppurative cortical interstitial nephritis with ballooning degeneration of epithelial cells of the renal pelvis, ballooning degeneration of the transitional epithelium of the urinary bladder, multifocal necrosis of the squamous epithelium of the palatine tonsils with depletion and necrosis of lymphoid tissue, and depletion and necrosis of lymphatic nodules of the spleen.
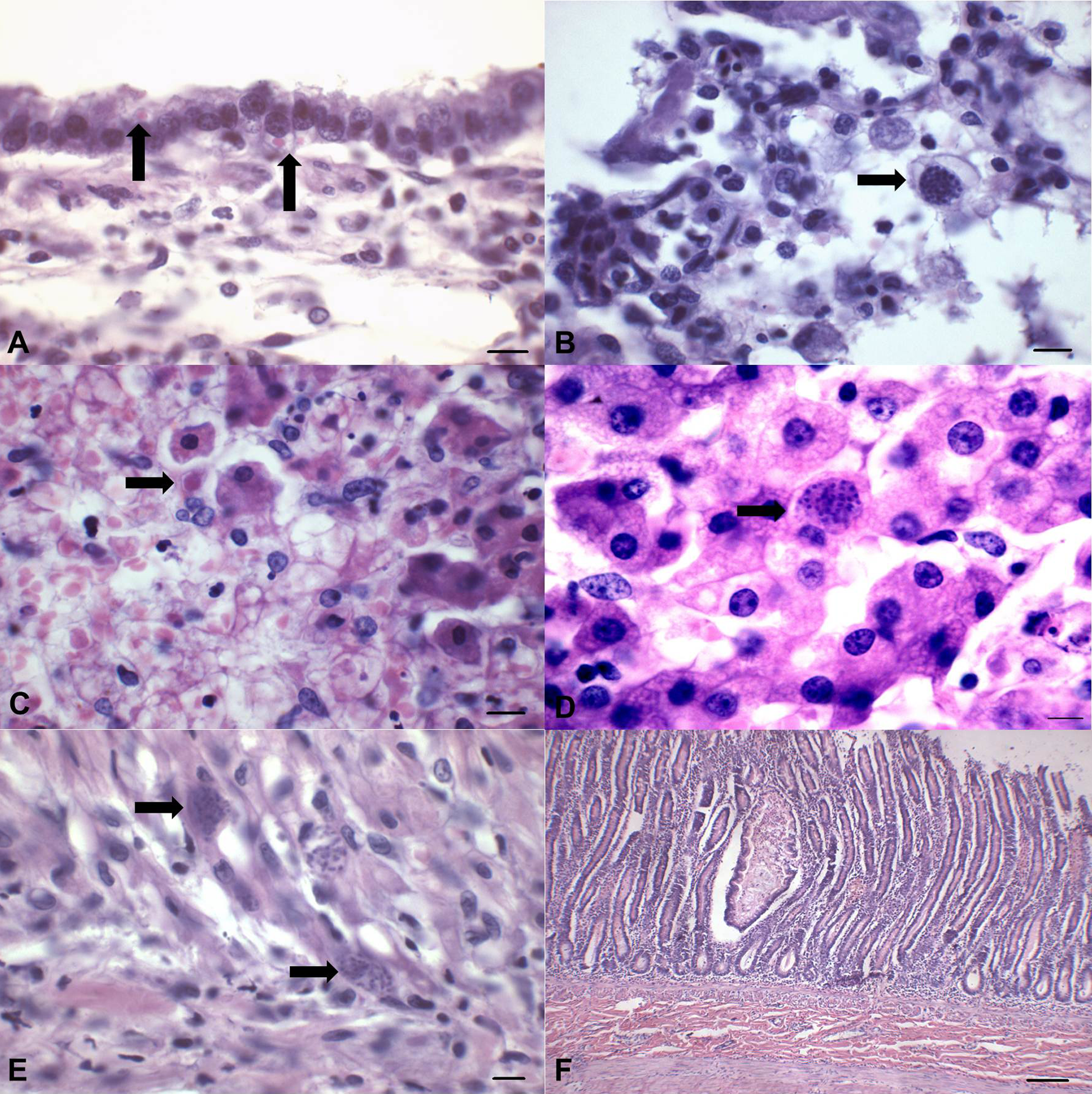
Histopathological characterization of concomitant Canine distemper virus (CDV), Canine adenovirus A type 1 (CAdV-1) and 2 (CPV-2), and Toxoplasma gondii infections in a puppy.
Gastrointestinal parasites were not observed in the samples evaluated by the zinc sulfate flotation technique. The IFAT based on the serum sample derived from the dam revealed a titer of 1,024 for anti–T. gondii antibodies.
The RT-PCR assay amplified the desired 290-bp fragment of the N gene of CDV derived from the urine, lung, brain, liver, spleen, and cervical lymph node samples of the puppy submitted for necropsy. The partial CDV N gene sequences were deposited in GenBank (accession no. JQ965820). Initial analysis using the BLAST program demonstrated that the sequences have 95–99% homology with other similar sequences deposited in GenBank. Phylogenetic analysis has shown that strains from Brazil based on this gene are divided into 2 clusters, one having more members relative to the other. When the sequences derived from the present study were compared with other similar sequences obtained from GenBank, it was demonstrated that these sequences are more closely related to other wild-type strains of CDV circulating in Brazil, particularly to a strain from the state of Paraná and others from São Paulo (Fig. 1). Further, these strains were grouped into the larger cluster of Brazilian wild-type strains of CDV. The PCR assays amplified the 508-bp and 1,030-bp fragments of the E gene of CAdV-1 and -2 from renal and hepatic tissue and from the urine sample, and the 583-bp fragment of CPV-2c VP2 gene from the cervical lymph node. However, CaHV-1 DNA was not amplified from any of the tissue samples analyzed, and positive CAdV-1 and -2 signaling was not observed by ISH.
The PCR assay amplified the 291-bp fragment of the 18S rDNA gene that is common to apicomplexan protozoa, while enzymatic digestion by PCR-RFLP confirmed that the samples derived from the myocardium (with and without mineralization and necrosis), liver (with and without pale foci), and lungs contained apicomplexan DNA specific for T. gondii. The PCR-RFLP multilocus genotyping was only achieved with the SAG3 marker, which revealed that the isolates of the current study are T. gondii genotype III.
The results of these investigations have confirmed the participation of CDV, CPV-2, CAdV-1 and -2, and Toxoplasma gondii in lesions by the combination of characteristic clinical signs and histopathological findings aided with molecular techniques in the manifestations of systemic CD, infectious tracheobronchitis, and toxoplasmosis with concomitant ICH and canine parvoviral enteritis in this puppy.6,9,12,13 The possible participation of CaHV-1 in the lesions observed in the puppy was excluded due to the negative results of the PCR assay that was designed to amplify the partial fragments of the CaHV-1 glycoprotein B gene. 22
A diagnosis of systemic CD was obtained due to histopathological alterations within the lungs and brain associated with eosinophilic inclusion bodies within epithelial cells; such findings are diagnostic for CDV,6,14 and have been described in other cases of CD.15,17 The amplification of viral RNA from the urine, brain, liver, spleen, and cervical lymph node extended the dissemination of this infection. Although inclusion bodies were not identifiable within the urinary bladder or the renal pelvis of the necropsied puppy, the ballooning degeneration observed in these organs is a frequent manifestation of CDV. 6
The sequences derived from the current study were more closely related to other wild-type strains of CDV circulating in Brazil, particularly from the state where the puppy resided. However, these sequences were quite distant phylogenetically from those (GenBank accession no. EF197736) derived previously from a dog (Fig. 1) from the same geographical location that was diagnosed with old dog encephalitis. 15 Hence, the initial phylogenetic results suggest that there might be at least 2 somewhat distant but related wild-type strains of CDV circulating in southern Brazil. A 2012 study demonstrated by phylogenetic analyses of the H gene that the strains of CDV in Brazil might represent part of a larger clade of CDV strains circulating in South America. 21 However, it is suggested that those strains in Brazil that were classified on the basis of nucleic acid sequence from the viral H gene should be correctly referred to as the southern Brazilian, or more specifically northern Paraná, wild-type strains of CDV. This is because these strains were obtained from 2 closely related geographical locations of continental Brazil and might represent 2 different wild-type strains of CDV circulating within these geographical regions of the country. 20
The apicomplexan protozoa observed within several tissues of the necropsied puppy by histopathology were confirmed as T. gondii by a combination of PCR and genetic typing; this molecular approach is diagnostic for T. gondii and is recommended for the differentiation of apicomplexan protozoa within tissue samples. 23 The elevated titer (1,024) of anti–T. gondii antibodies seen in the dam could be indicative of acute toxoplasmosis, 11 while the lesions (pulmonary, hepatic, and cardiac) observed in the puppy are consistent with systemic toxoplasmosis.9,11 Collective analyses of the results would suggest that the puppy was most likely infected congenitally; the dam was only 10 months of age at the time of parturition and might have been infected during pregnancy. Although congenital toxoplasmosis in dogs is rare, 9 a 2009 study described suspected spontaneous transplacental transmission confirmed by PCR-RFLP in 6 puppies born to an 18-month-old, female cattle dog from Australia. 1
The initial diagnosis of CPV-2 was based on characteristic pathological findings that are consistent with this disease4,13; the participation of CPV-2 in these lesions was confirmed by amplifying the desired 583-bp fragment of the VP2 gene by using primers that are specific for CPV-2c. 18 The intestinal lesions (cryptal necrosis, atrophy and fusion of intestinal villi, and lymphoid depletion) observed in the necropsied puppy are highly suggestive of canine parvoviral enteritis.4,13
Infectious canine hepatitis was presumptively diagnosed by the characteristic clinical manifestation of CAdV-1–induced corneal edema and necrotizing hepatitis associated with large intranuclear inclusion bodies predominantly within Kupffer cells and in few hepatocytes; such findings are diagnostic for ICH.4,12 Confirmation of infection was obtained from the PCR assay that successfully amplified the 508-bp fragment of CAdV-1; similar findings have been described. 19
Although characteristic CAdV-2 intranuclear inclusion bodies were not observed within the lung of the puppy, the necrotizing lesions seen within the pulmonary airways can be partially attributed to this pathogen; similar findings have been described.7,10 The amplification of viral DNA from the kidney and urine of this dog would suggest that CAdV-2 is excreted via the urinary system, and clinical urinal samples can be used to identify this agent by PCR. As far as the authors of the current study are aware, this method of viral excretion was never proposed for CAdV-2. However, urinary excretion occurs with CAdV-1, where viral identification within the kidney can be demonstrated long after the virus has been cleared from the liver. 12
Although there are several reports of coinfections with the participation of CDV in dogs,7,8,17 the uniqueness of the current report lies in the concomitant characterization of 5 important canine diseases in the same puppy. In the current case, the immunosuppressive effects of CDV resulting in the destruction of signaling lymphocytic activation molecule (SLAM)-producing cells, 2 coupled with the underdeveloped immune system of the puppy, would have facilitated the simultaneous occurrence and maintenance of these infectious disease agents. Additionally, concomitant systemic CD and toxoplasmosis, as observed in the puppy, are common in immunologically immature neonates 4 ; while the age of the puppy makes the animal highly susceptible to all infections herein described.6,9,12-14 Further, the effects of undernourishment, considering the large number of littermates (14), cannot be overlooked as a contributory factor to the development of these diseases. Moreover, the current study highlights the importance of infections induced by CDV and its related consequences to the urban canine populations of Brazil. 16 Furthermore, because CDV coupled with age and undernourishment might have been the predisposing factors to the other infectious agents, the acute hepatitis due to CAdV-1 with the concomitant hepatic and pulmonary toxoplasmosis and the necrotizing protozoan myocarditis most likely would have resulted in multiple organ failure and consequently death. In conclusion, the present study is an excellent demonstration of the effects of multifactorial conditions (age of the dog, infectious agents, immunosuppression, and probably starvation or at least undernutrition) in the development of disease.
Footnotes
Acknowledgements
The authors thank Karin Fragner, University of Veterinary Medicine, Vienna, Austria, for technical expertise in the realization of in situ hybridization, and Dr. Paulo C. Werner, for submitting the puppy for necropsy.
a.
Invitrogen Corp., Carlsbad, CA.
b.
illustra GFX PCR DNA and Gel Band Purification Kit, GE Healthcare, Little Chalfont, Buckinghamshire, UK.
c.
QIAamp DNA FFPE Tissue Kit, Qiagen Inc., Valencia, CA.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
Drs. SA Headley, AF Alfieri, JL Garcia, and AA Alfieri are recipients of the National Council for Scientific and Technological Development (CNPq; Brazil) fellowships.