
Abstract
Morphological lesions in kidneys and brain are all too often considered diagnostic for confirmation of encephalitozoonosis in rabbits. The current study evaluated the diagnostic value of histology versus other etiological tests, including immunohistochemistry and real-time polymerase chain reaction (PCR) for Encephalitozoon cuniculi infection diagnosis. Samples of brain, heart, lungs, intestine, liver, and kidneys from 81 rabbits were examined for morphological lesions attributed to E. cuniculi infection as well as for the presence of spores and E. cuniculi antigen. Of these, 55 rabbits were tested for E. cuniculi DNA. Histological changes consistent with E. cuniculi infection were seen in 33 rabbits (41%, 33/81) representing 87% (33/38) of all rabbits with confirmed E. cuniculi infection. Brains of these rabbits displayed 6 different types of focal lesions corresponding to the stage of infection and specific tissue response. In 5 rabbits that were tested positive, histology was either inconclusive or inconspicuous. Etiological diagnosis was based on histological spore detection in 16% (6/38) of infected rabbits. Immunohistochemistry was more sensitive (42%, 16/38) than histological spore detection, and real-time PCR proved to be the most sensitive of all investigated methods (30/35, 86% of the examined rabbits with E. cuniculi infection). Encephalitozoon cuniculi infection rarely occurs without characteristic kidney and brain lesions. However, the spectrum of brain changes is wider than previously reported. Based on these findings, confirmation of pathogenic E. cuniculi infection should include standard histology of the predilection sites and a specific etiological assay, preferably real-time PCR.
Introduction
Encephalitozoonosis is a common disease of rabbits caused by Encephalitozoon cuniculi. 23 The disease was first reported in 1922. 35 Rabbits affected with encephalitozoonosis most frequently exhibit multiple neurologic signs (head tilt, ataxia, circling, nystagmus, rotational movements around the body length axis, seizures, paresis, head tremors, swaying or nodding at rest, and behavior changes), kidney dysfunction (azotemia), and phacoclastic uveitis. These 3 manifestations may occur in combination or individually.18,22 A clinical tentative diagnosis is usually based on the combination of neurologic deficits and high serum antibody titers or detection of spores in affected tissues. To date, histopathology is still considered the standard for definitive diagnosis. However, a variety of techniques, including special staining methods, immunohistochemistry (IHC), and polymerase chain reaction (PCR) are used for postmortem diagnosis and exclusion of other microorganisms such as Toxoplasma gondii.1,2,4,7,10,17
The most characteristic morphological feature is the presence of granulomatous encephalitis together with chronic interstitial nephritis.4,10,21 Although, encephalitozoonosis is a common disease in rabbits, information on histopathology of the disease, including description of the changes in visceral organs of a larger number of pet rabbits with a natural infection, is scant. Furthermore, most published reports describe lesions in rabbits younger than 1 year. 6 Likewise, most reports on immunohistochemical assays focus on a small number of rabbits and a limited range of investigated organs.15,27
Previously, the detection of E. cuniculi DNA in tissues or body fluids, including urine and cerebrospinal fluid, employed conventional PCR or nested PCR.1,7,22,23,31 To detect E. cuniculi in tissue samples, conventional PCR proved sufficient only for the identification of E. cuniculi from lens tissue. Nested PCR has been reported to achieve a higher sensitivity, 7 but in contrast with real-time PCR, nested PCR results in a higher risk of sample contamination. Real-time PCR has been previously used for identification of E. cuniculi, but primarily for the identification of the microorganism in environmental samples or human feces and body fluids. 14
An aim of the current study was to elucidate the histopathological changes in various organs of rabbits affected by encephalitozoonosis and to investigate the presence of this microorganism in the tissue. Thus, a large population of rabbits was investigated for possible infection with E. cuniculi using histopathology (the current standard method), IHC, and real-time PCR. The present study further assessed the comparative diagnostic feasibilities of the applied procedures.
Material and methods
Confirmative diagnosis—a definition
A rabbit was defined as being infected with E. cuniculi when typical histopathological lesions (granulomatous encephalitis and interstitial nephritis) were detected and/or E. cuniculi was identified with at least 1 of the following methods: histology, IHC, and real-time PCR. The term encephalitozoonosis is used for E. cuniculi infection associated with tissue damage.
Rabbits
Tissues from 81 randomly selected pet rabbits (38 males, 43 females; aged 3 weeks to 9 years, mean: 2.6 years) were acquired from necropsy cases delivered to the Institute of Veterinary Pathology of the Ludwig-Maximilians-University (Munich, Germany). All rabbits were presented for diagnostic purposes unrelated to the current study. A complete necropsy was performed, and tissue samples were collected as described below. Two samples each of brain, kidneys, lungs, liver, heart, and intestine were immediately frozen and stored at −70°C until further processing for real-time PCR. Two additional samples of each of these organs were placed in 10% neutral buffered formalin for histopathological examination and immunohistochemical staining. In 24 rabbits, the clinical history of neurological deficits suggested that an infection with E. cuniculi was very likely. In addition to the tissues mentioned above, both eyes underwent histological examination if they exhibited gross abnormal changes.
Histopathological analysis
Histopathological examination was performed on brain, heart, lungs, intestine, liver, and kidneys. Fixed tissues were routinely processed, embedded in both paraffin and plastic, 19 and stained with hematoxylin and eosin as well as Giemsa. The tissues were evaluated according to standard histopathological algorithms. All pathologic changes were recorded, and particular attention was directed at granulomatous and lymphoplasmacytic inflammation in the brain and renal tissue. In accordance with previous studies,10,21 granulomatous encephalitis and lymphoplasmacytic meningoencephalitis in combination with multifocal interstitial nephritis was accepted as being characteristic of E. cuniculi infection. After bihemispheric transverse sectioning of the full brain at 3 standard levels (middle cerebellar peduncles, infundibulum, and rostral border of optic chiasm), the following brain areas were separately assessed to elucidate the spatial distribution of possible lesions: brainstem, cerebellum, hippocampus, thalamus, basal nuclei, and neopallium. Cortical affection was subcategorized into lesions of the inner, middle, or outer layers. Focal brain lesions were characterized with regard to the different types and stages of tissue alterations. Major determinants for the subtyping of these focal lesions were destruction of the neuroparenchyma, inflammatory cell infiltrates, as well as reactive proliferation of astroglia and microglia. The number of focal lesions were counted within the different regions per animal (lesions per region per animal) and then summed to obtain the incidence of foci per region.
Immunohistochemical analysis
Immunohistochemistry was performed on dewaxed 4-μm thick sections of all tissues harvested. The staining was carried out at room temperature and employed a horseradish peroxidase–diaminobenzidine/HistoGreen detection system with avidin–biotin complex amplifier.
Mouse monoclonal antibodies directed against 56-, 45-, and 43-kDa proteins of E. cuniculi were used as primary antibodiesa,25 in 1:1,000 dilution. In a previous study, 25 these monoclonal antibodies reacted with E. cuniculi but did not react with Encephalitozoon intestinalis, Encephalitozoon hellem, Escherichia coli, Enterococcus faecalis, Pseudomonas aeruginosa, Klebsiella pneumoniae, Shigella dysenteriae, Proteus vulgaris, Salmonella enterica, or Aspergillus fumigatus.
Before labeling, tissue sections were treated with 3% hydrogen peroxide to quench endogenous peroxidase activity and then washed in a bath of Tris buffered saline (TBS; 0.5 M, pH 7.6). Sections were preincubated with normal goat serum b for 30 min, followed by 60-min incubation with the respective mouse anti–E. cuniculi antiserum. Sections were then washed in baths of TBS, incubated with the secondary antibody (biotinylated goat anti-mouse immunoglobulin) c for 1 hr, and washed in a bath of TBS again. Sections were incubated with a commercial peroxidase-labeled avidin–biotin complex. d The antigen–antibody complex was visualized by the reaction with H2O2 and 3′,3′-diaminobenzidine (DAB) e or HistoGreen f as chromogen. Slides were counterstained with Mayer hematoxylin or kernechtrot. HistoGreen was used to avoid color confusion in some cases because detection of antigen using DAB as chromogen was difficult due to the presence of brownish pigment, particularly in the kidneys.
Nucleic acid isolation and real-time PCR
Following the manufacturer’s instructions, a DNA blood and tissue kit g was used to extract total DNA from frozen tissue samples of intestine, brain, heart, liver, lungs, and kidneys collected at necropsy from 55 rabbits. For E. cuniculi identification, the extracted DNA was subjected to duplex real-time PCR analysis using the primers E. cuniculi forward primer (MSP-3: 5′-TTGCGATGAAGGACGAAGG-3′), E. hellem forward primer (MSP-4: 5′-TGATGAAGGACGAAGG-3′), a shared reverse primer (MSP-5: 5′-TCTTGCGAGCG TACTATCC-3′), and the molecular beacon fluorescence probes E. cuniculi (MSP-S3: 5′-CGCGATCGACTGGACGG GACNGTGTGTGTTGTCCATGAGAAAGATCGCG-3′) and E. hellem (MSP-S4: 5′-CGCGATCGACTGGACGGG ACTGTTTTAGTGTTGTCCGAGAGAAAGATCGCG-3′). The molecular beacon probes MSP-S3 and MSP-S4 were labeled with 5-carboxyfluorescein (FAM) and hexachloro-6-carboxyfluorescein (HEX), respectively, as the 5′-fluorescent reporter and with 4-(4′-dimethylaminophenylazo)-benzene (Dabcyl) as the 3′-quencher. Primers and probes were commercially manufactured. h Real-time PCR was performed using commercially available reagent sets. i Five microliters of DNA from tissue samples was added to the master mix in a reaction (50 μl total volume) containing 7 mM MgCl2, i 1 μM each of forward and reverse primers, 0.5 μM each of the molecular beacon probes MSP-S3 and MSP-S4, 5 units Taq polymerase, i 5 μg of bovine serum albumin, j 0.25 mM deoxyribonucleotide triphosphates k each, and 1× PCR buffer. i
Thermocycling conditions included one 10-min cycle at 95°C, fifty 30-sec cycles at 94°C, and one 60-sec cycle at 53°C. No amplification was detected in positive control samples of E. hellem, E. intestinalis, and Enterocytozoon bieneusi.
Results
Histopathological analysis
Of the 81 rabbits, characteristic histopathological lesions (granulomatous encephalitis in combination with an interstitial nephritis4,10,21) were found in 33 of the rabbits. Thirty-two rabbits did not have lesions suggestive of E. cuniculi infection. Infection was considered possible in 5 rabbits with chronic lymphoplasmacytic interstitial nephritis and multifocal lymphoplasmacytic meningoencephalitis, 10 rabbits with chronic lymphoplasmacytic interstitial nephritis only, and 1 rabbit with focal microglial activation and nerve cell necrosis. Eleven of the 16 rabbits with possible E. cuniculi–associated pathology and 14 rabbits without characteristic histopathological lesions underwent further etiological tests using real-time PCR. A detailed description of the 38 rabbits with confirmed E. cuniculi infections, according to the above-mentioned definition for confirmative diagnosis, is provided in the following sections.
Lesions in the kidneys
In 2 of the examined rabbits, spores were observed in the kidneys. Spores were localized within parasitophorous vacuoles in tubular epithelial cells, most frequently in the medulla (Fig. 1). Parasitophorous vacuoles were found in all segments of the nephron. The most commonly affected segment of the nephron was the inner medullary collecting duct. Lesions were subclassified according to stage into acute, chronic, or both. Acute lesions were characterized by focal tubular necrosis with inflammatory infiltrates consisting of heterophils, macrophages, lymphocytes, and plasma cells (Fig. 2). Such lesions were detectable in 19 of the 38 (50%) rabbits with an E. cuniculi infection. In addition, dilated tubules, often filled with detritus, and regeneration of the tubular epithelial cells were observed. Acute lesions were most frequently found of mild severity in the medulla in the form of round foci.
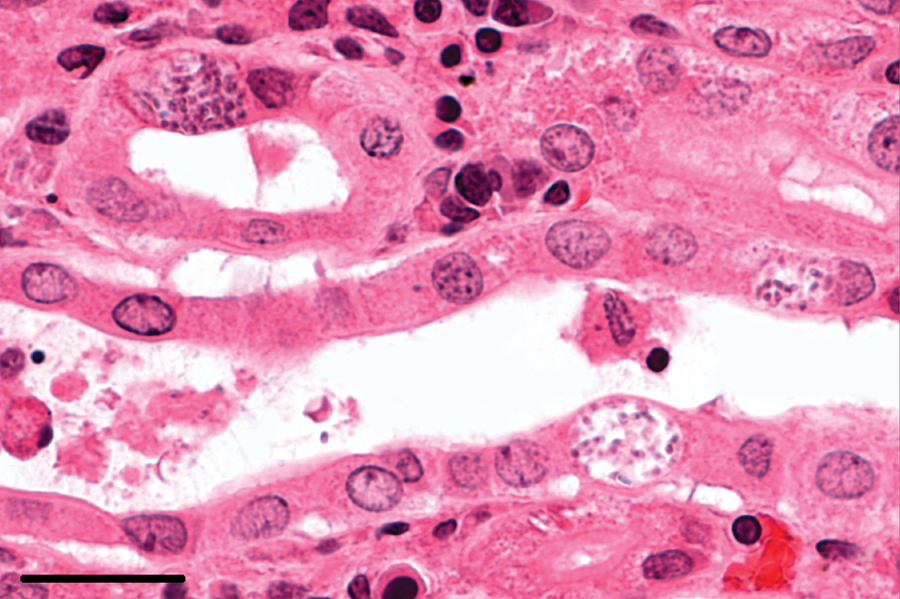
Kidney; rabbit no. 80. Spores within parasitophorous vacuoles in the tubular epithelium. Hematoxylin and eosin. Bar = 25 μm.
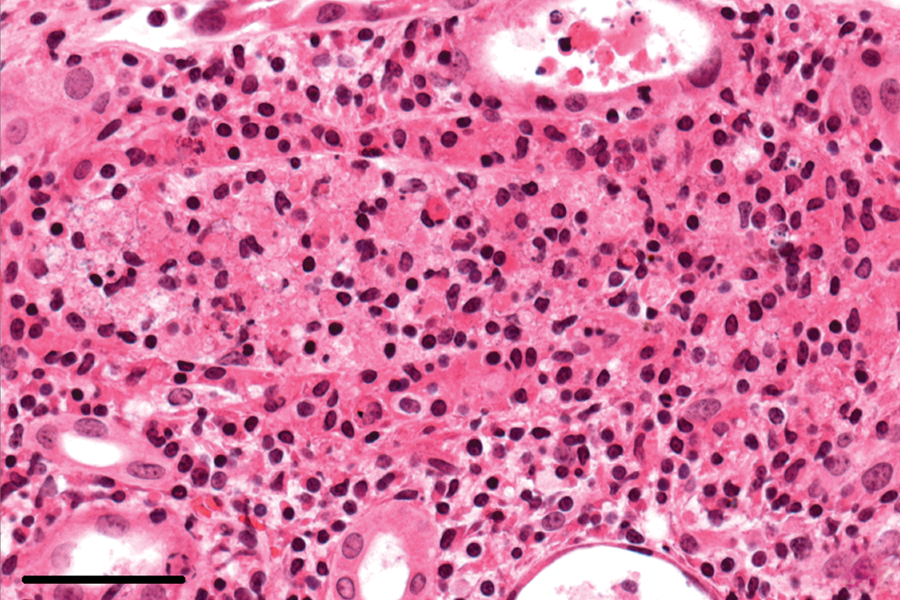
Kidney; rabbit no. 80. Acute inflammation with necrosis. Hematoxylin and eosin. Bar = 50 μm.
Chronic lesions in the kidneys, characterized by interstitial mononuclear cell infiltration, were found in 34 of the 38 (89%) rabbits with an E. cuniculi infection and occurred primarily in the cortex (Fig. 3). The chronic renal lesions were accompanied by interstitial fibrosis, which varied from small subcapsular foci to large areas reaching into the medulla, in 32 of the 34 (94%) rabbits (Fig. 4). In addition to these focal changes, diffuse interstitial fibrosis in the cortex–medulla junction was observed in areas adjacent to chronic inflammation (Fig. 5). End-stage kidneys were identified in 3 out of the 38 (8%) rabbits with an E. cuniculi infection. In kidneys with predominantly chronic lesions, no spores were detectable.
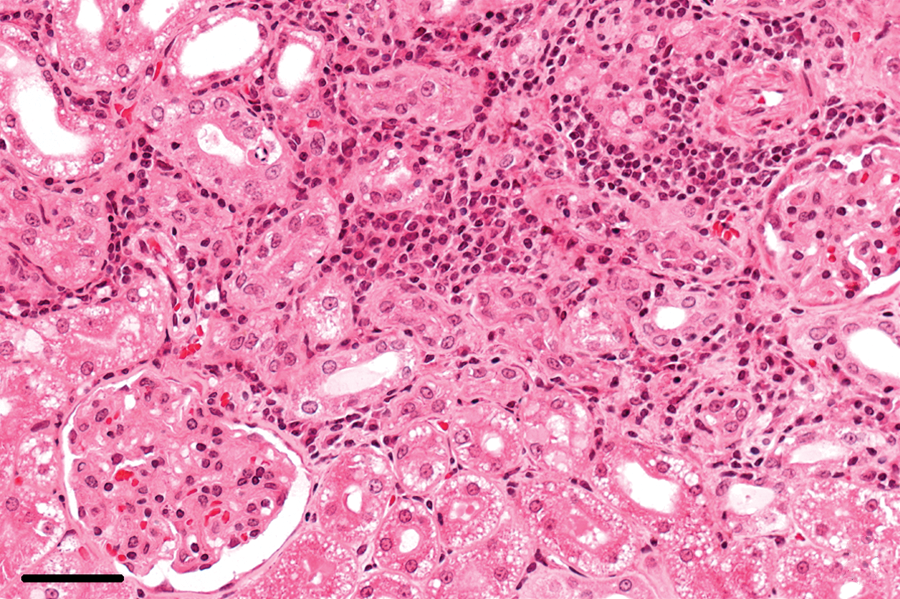
Kidney; rabbit no. 62. Chronic interstitial lymphoplasmacytic nephritis. Hematoxylin and eosin. Bar = 50 μm.
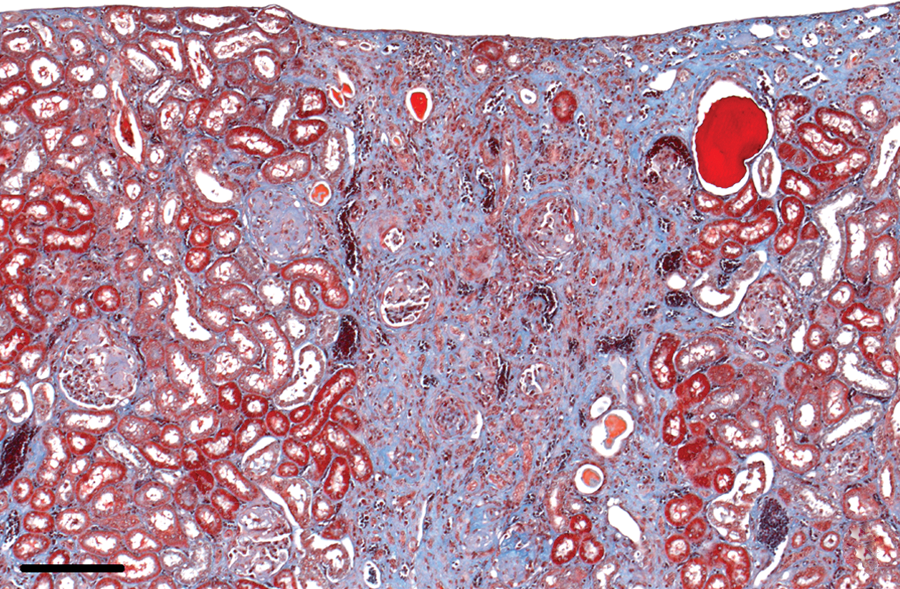
Kidney; rabbit no. 9. Cuneiform fibrosis in the renal cortex. Masson trichrome stain. Bar = 200 μm.
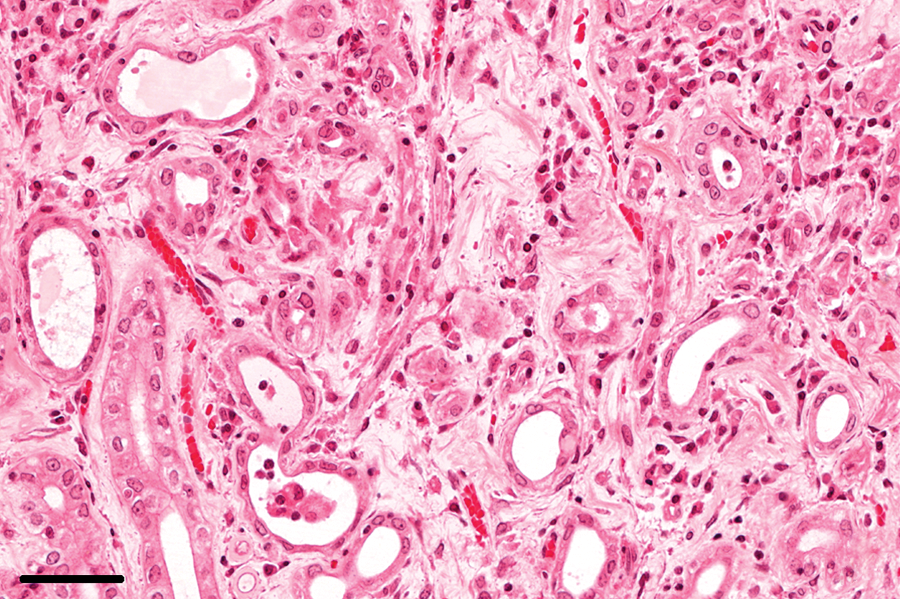
Kidney; rabbit no. 62. Severe interstitial diffuse fibrosis with tubule atrophy and inflammatory infiltration. Hematoxylin and eosin. Bar = 50 μm.
Lesions in the brain
In 6 of the 38 (16%) infected rabbits, spores were detected in the brain. Spores were located in parasitophorous vacuoles in the parenchyma of the brain, either without inflammation (Fig. 6, left panel) or close to focal inflammatory lesions. Host cells infected by the microorganism were not identified in neuropil, but occasionally parasitophorous vacuoles were observed in the endothelial cells of cerebral and meningeal microvasculature (Fig. 6, right panel). In 1 case, a ruptured parasitophorous vacuole was associated with early inflammatory infiltrates (Fig. 7). In another case, free spores were present in the necrotic center of a granuloma.
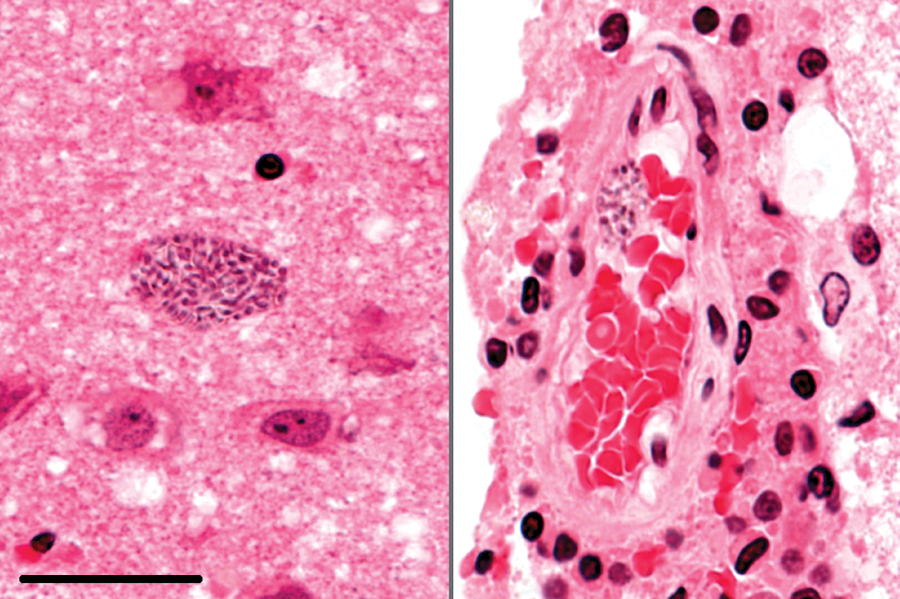
Brain; rabbit no. 6. Spores within parasitophorous vacuoles in the neuropil (left) and in the endothelium of the leptomeninx (right). Hematoxylin and eosin. Bar = 25 μm.
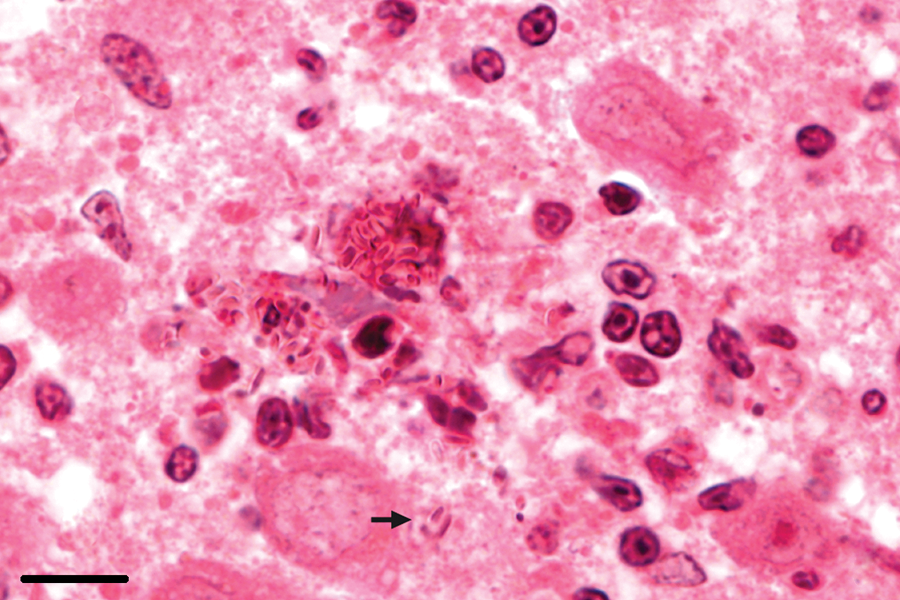
Brain; rabbit no. 46. Ruptured parasitophorous vacuole with free spores in the neuropil (arrow) and beginning inflammatory reaction. Hematoxylin and eosin. Bar = 10 μm.
Focal brain lesions were identified in 33 of the 38 (87%) rabbits with confirmed infection. These lesions were categorized into 6 histopathological subtypes (I–VI).
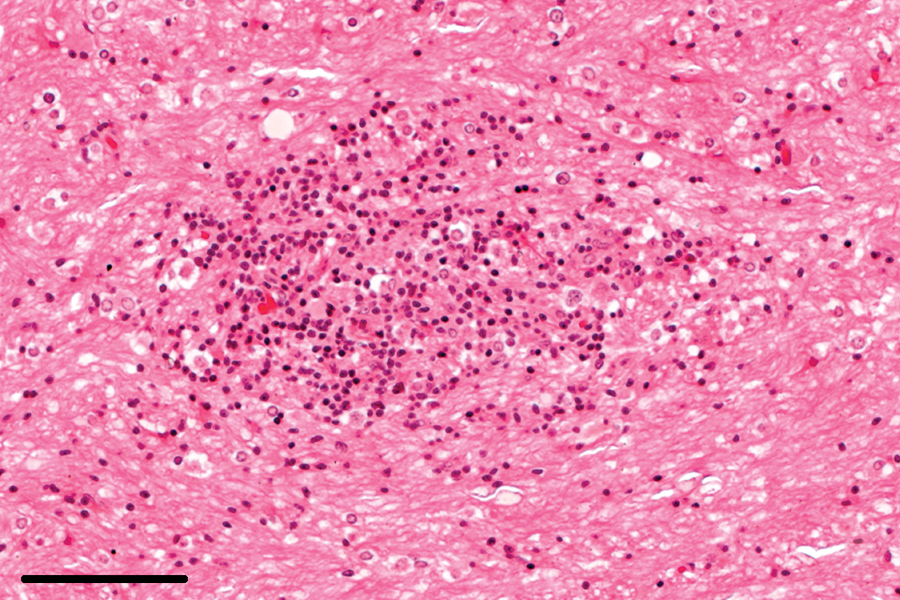
Type I lesions consisted of small, poorly circumscribed aggregates of lymphocytes, frequently intermingling with heterophils, and early astrogliosis. In type II lesions, lymphocytic and heterophilic infiltration and the astrocytic reaction became more advanced and were accompanied by invading plasma cells (Fig. 8). In type III lesions, the inflammatory gliotic foci exhibited necrotic centers (Fig. 9). In type IV lesions, significant involvement of macrophages and protoplasmic, gemistocytic astrocytes was noted, while lymphocytic infiltration was attenuated. These lesions frequently presented with epithelioid histiocytes and occasional multinucleated giant cells (Fig. 10), and their centers quite often appeared necrotic, with or without dystrophic mineralization. Type V lesions were characterized by significant fibrillary astrogliosis (prominent astrocyte processes) and astrocytosis (an increased number of astrocytes), demarcating the granulomatous foci from the surrounding brain tissue. Active inflammation of epithelioid cells and scattered lymphocytes and plasma cells were still evident (Fig. 11). Lesions of type VI were restricted to a glial scar, partly with a central hypocellular area containing very rare postresorptive macrophages (Fig. 12).
Brain; rabbit no. 46. Early lesions with poorly circumscribed inflammatory infiltrate and astrocytosis. Hematoxylin and eosin. Bar = 100 μm.
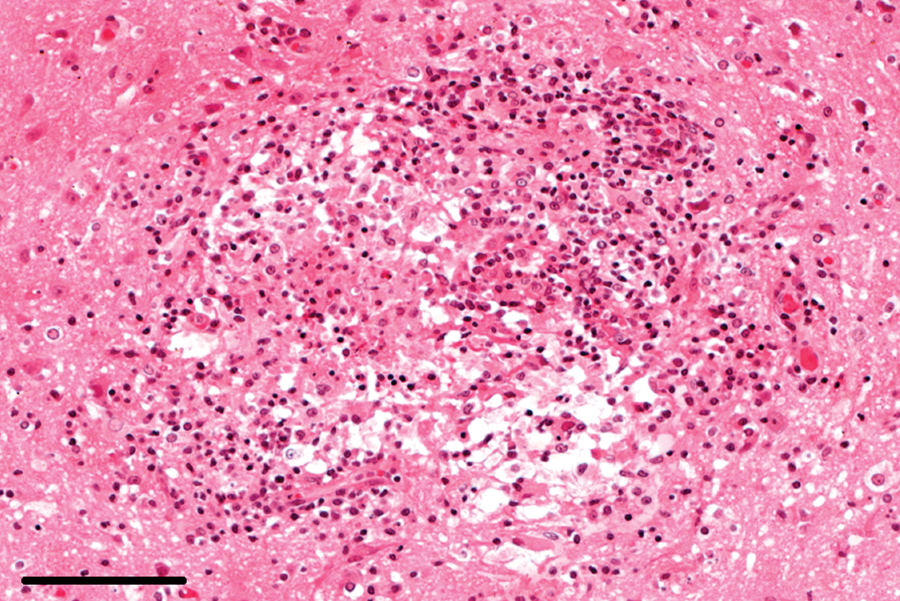
Brain; rabbit no. 34. Inflammatory-gliotic foci with central necrosis. Hematoxylin and eosin. Bar = 100 μm.
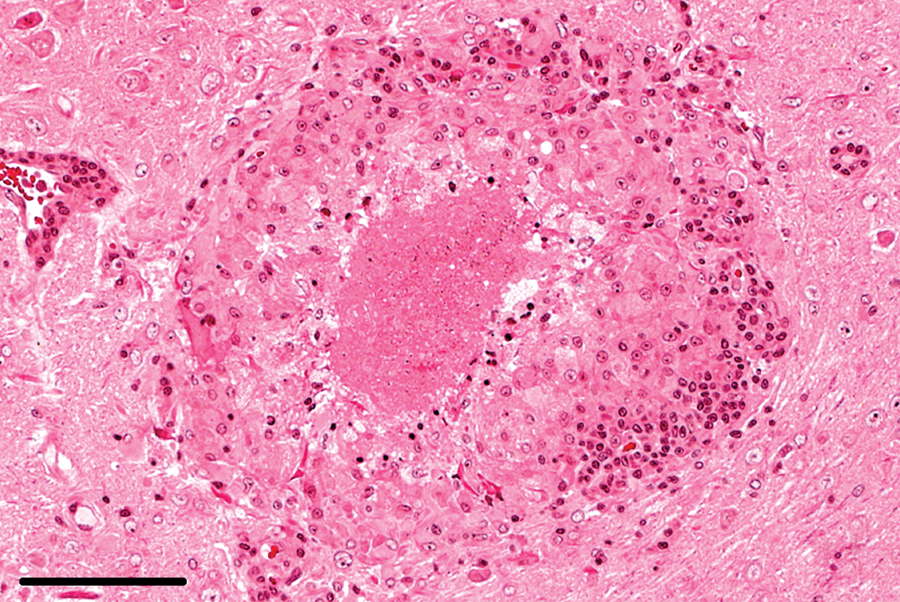
Brain; rabbit no. 3. Granuloma with central necrosis, significant involvement of macrophages and lymphocytic encephalitis in adjacent tissue. Hematoxylin and eosin. Bar = 100 μm.
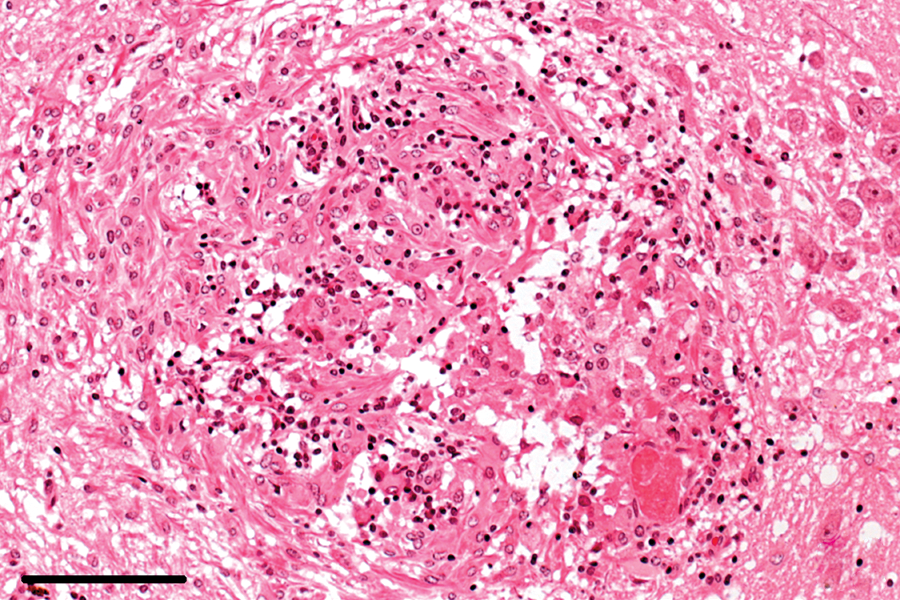
Brain; rabbit no. 46. Late granuloma, demarcated by fibrillary astrocytes and a minor degree of acute inflammation. Hematoxylin and eosin. Bar = 100 μm.
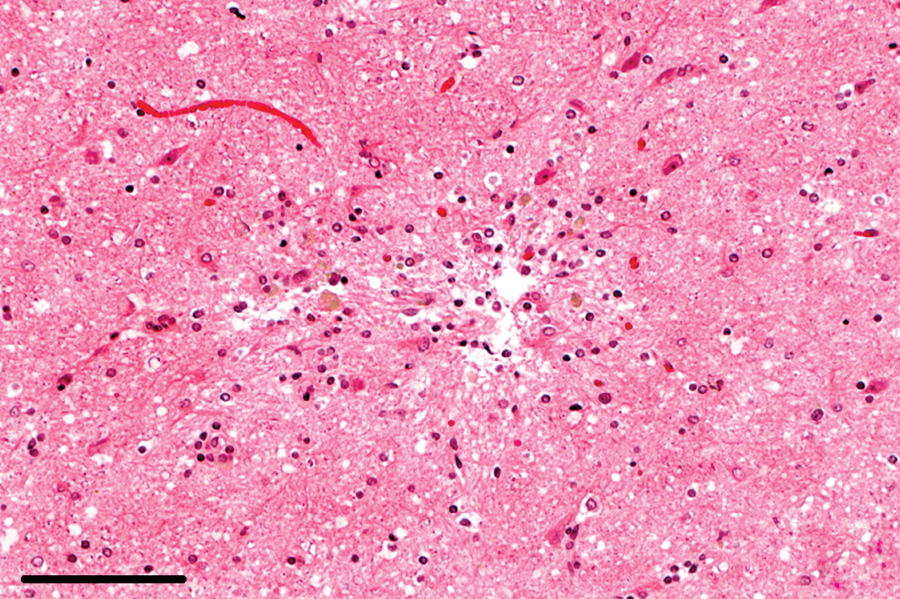
Brain; rabbit no. 44. Glial scar with a hypocellular area and postresorptive macrophages. Hematoxylin and eosin. Bar = 100 μm.
The incidence of different types of focal brain lesions is indicated in Tables 1 and 2. Type I lesions were observed in 17 rabbits, ranging from 1 to 20 foci per brain region throughout all affected rabbits (mean: 7). The lesions most frequently involved the neocortex (20/56 foci throughout the rabbits), thalamus (14/56), and hippocampus (10/56), followed by other areas of gray matter in the cerebrum. Type II lesions were observed in 24 rabbits (57 foci throughout the rabbits, 1–20 foci per region, mean: 7.1). The lesions were as frequently observed as type I lesions and had an identical distribution pattern. Type III lesions were slightly less frequent than types I and II (19 rabbits, 50 foci throughout the rabbits, 1–22 foci per region, mean: 6.25). The basal nuclei were affected second most often, following the neocortex. Type IV lesions were the most commonly observed focal lesions (22 rabbits, 64 foci throughout the rabbits, 0–33 foci per region, mean: 8). The predominance in the cerebral cortex was also evident in type IV lesions. Similar to type II and III lesions, the innermost layers of the neocortex were the most commonly affected. The second most common site of manifestation was in the thalamus. Type V lesions were observed in 12 rabbits (20 foci throughout the rabbits, 0–10 foci per region, mean: 2.5), and type VI lesions were observed in 13 rabbits (20 foci throughout the rabbits, 0–15 foci per region, mean: 2.5). Both of these lesion types (V and VI) were most frequent in the neocortex.
Incidence of different types of focal brain lesions (types I–VI), according to location, in Encephalitozoon cuniculi encephalitis–affected rabbits.*
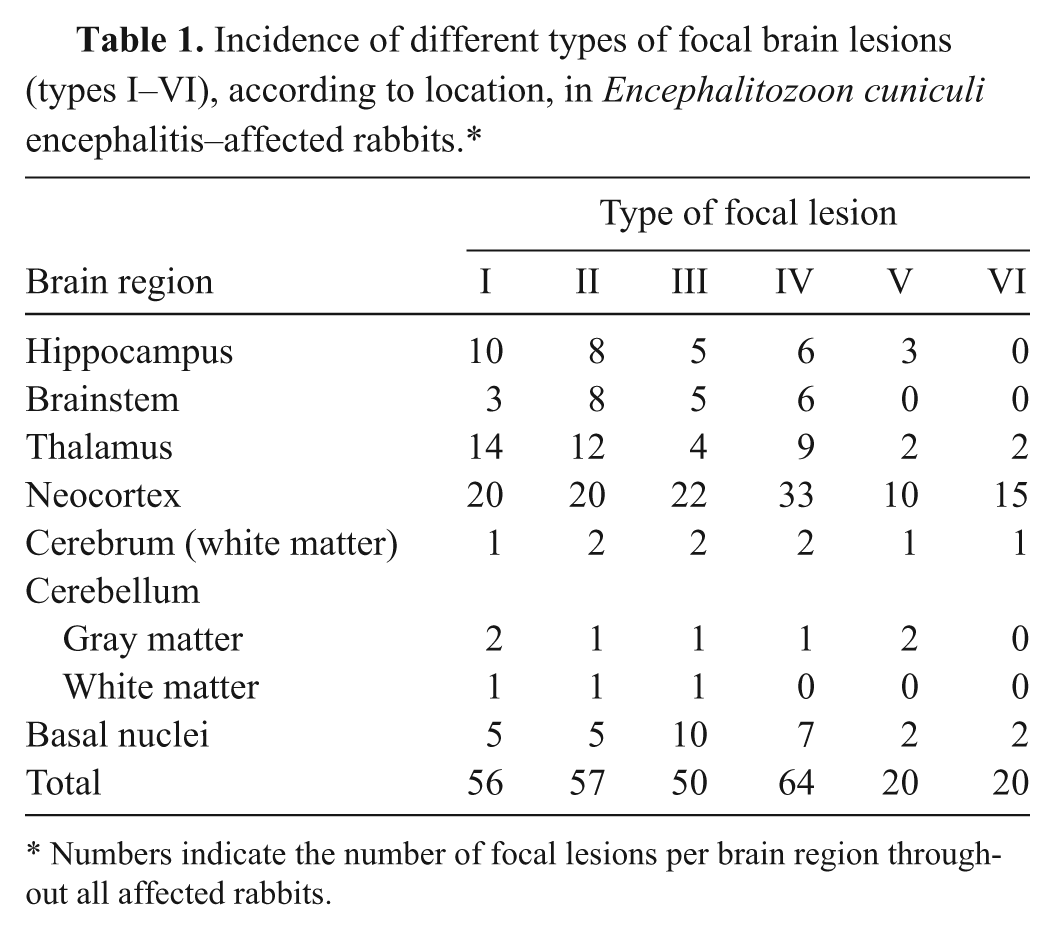
Numbers indicate the number of focal lesions per brain region throughout all affected rabbits.
Incidence of different types of focal lesions in the neocortices of Encephalitozoon cuniculi encephalitis–affected rabbits.

Granulomas were most frequently found in the neopallium (31/33 affected rabbits), followed by the thalamus (17/33), the hippocampus (16/33), and the basal nuclei (16/33). The brain stem (15/33) was less frequently affected. Granulomatous lesions were observed in the cerebellum at a lower frequency than in the other regions (9/33). In summary, focal lesions were largely restricted to the gray matter and primarily spared the white matter (Table 1). In the cerebral cortex, the inner layers were more commonly affected than the middle or the outer layers (Table 2).
In addition to focal parenchymal lesions with tissue destruction and remodeling, all rabbits with granulomatous lesions also exhibited lymphoplasmacytic meningoencephalitis, which was characterized by perivascular cuffs and leptomeningeal infiltrates consisting primarily of lymphocytes and plasma cells (Fig. 13). These inflammatory changes were also observed in 5 rabbits lacking other brain lesions and in 1 rabbit with focal neuronal degeneration.
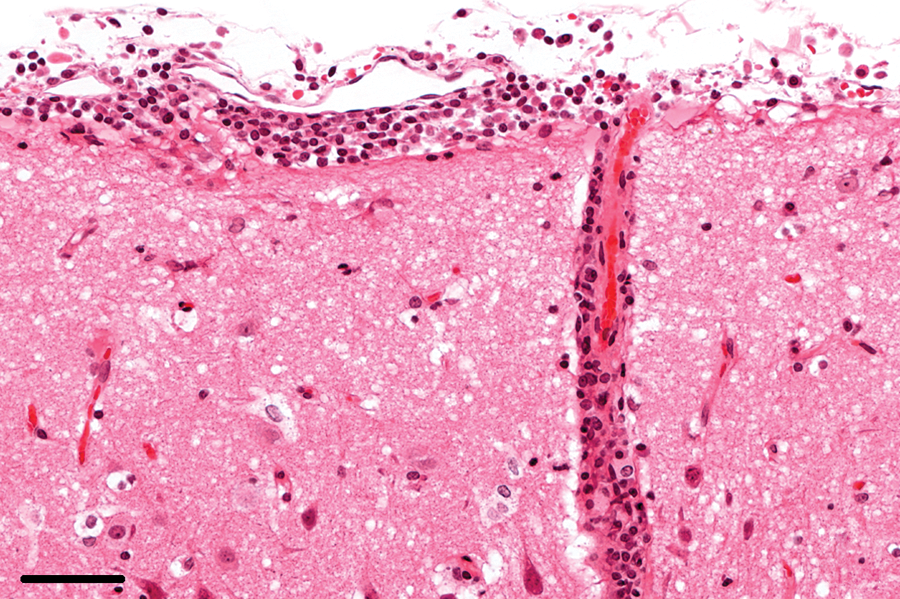
Brain; rabbit no. 81. Severe lymphoplasmacytic infiltration in the leptomeninx. Hematoxylin and eosin. Bar = 50 μm.
Lymphoplasmacytic meningoencephalitis most frequently affected the neopallium (33/33), followed by the hippocampus, the basal nuclei, and the thalamus (26/33 each). The brain stem (23/33) and cerebellum (22/33) were less frequently involved.
Lesions in other organs
In the setting of an active E. cuniculi infection, focal granulomatous infiltrates were observed in the lungs of 37% (14/38), the livers of 16% (6/38), and the hearts of 5% (2/38) of the infected rabbits (Figs. 14, 15). Lymphoplasmacytic periportal infiltrates in the liver and lymphoplasmacytic myocarditis were noted in 95% (36/38) and 58% (22/38), respectively, of the infected rabbits. In 58% (22/38) of the infected rabbits, interstitial, lymphoplasmacytic infiltrates were found in the lungs, partly with hyperplasia of bronchiolus-associated lymphoid tissue. In 1 case, phacoclastic uveitis with spores was observed in the lens. In the remaining rabbits without confirmed E. cuniculi infection (n = 43), focal granulomatous infiltrates were not found in the liver. Granulomatous inflammation was observed in the lungs, caused by lungworms, of 5% (2/43) and in the heart of 2% (1/43) of the noninfected rabbits. Lymphoplasmacytic infiltrates were noted in the liver of 19% (8/43), in the lungs of 7% (3/43), and in the heart of 7% (3/43) of the rabbits without confirmed E. cuniculi infections.
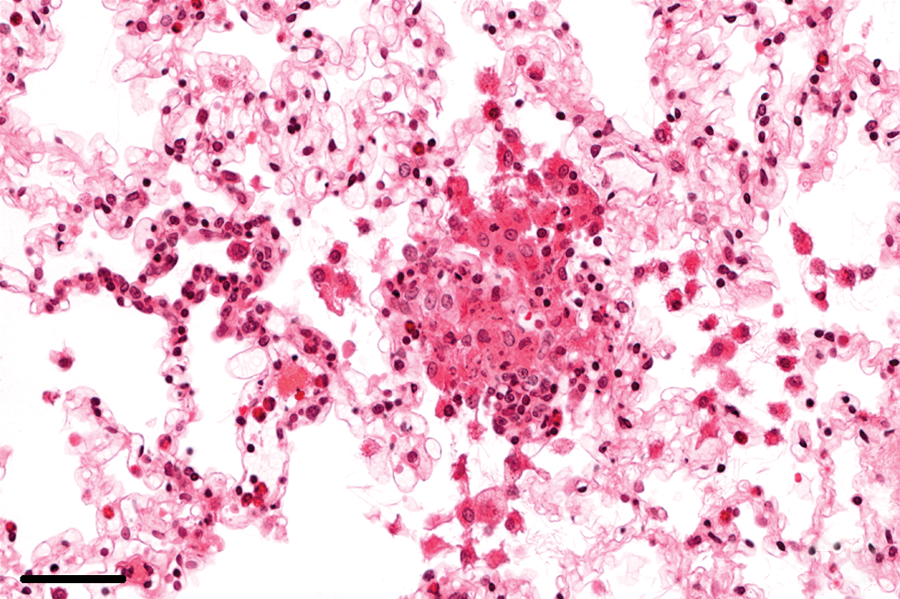
Lung; rabbit no. 36. Focal, inflammatory, predominantly histiocytic infiltrates in the lung parenchyma. Hematoxylin and eosin. Bar = 50 μm.
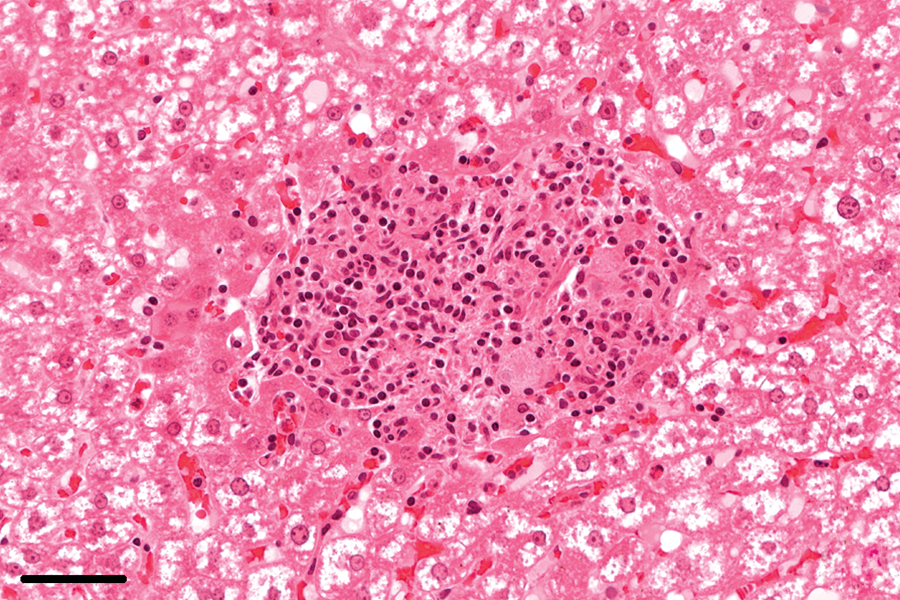
Liver; rabbit no. 46. Focal, inflammatory, predominantly histiocytic infiltrates in the parenchyma of the liver. Hematoxylin and eosin. Bar = 50 μm.
Immunohistochemical analysis
Encephalitozoon cuniculi antigen was detected in at least 1 organ in 42% (16/38) of the infected rabbits. The distribution of immunopositivity among organs is shown in Table 3. Antigen was most frequently detected in the brain and kidneys (11 rabbits each), followed by heart and lungs (1 rabbit each). In 6 rabbits, staining was observed in both brain and kidneys, whereas the other 5 rabbits proved immunopositive in only 1 of these 2 organs. Encephalitozoon cuniculi antigen was observed in 1 case each in heart and lungs, within a parasitophorous vacuole. In both cases, E. cuniculi antigen was also observed in brain and kidneys. The kidneys of 2 of the 11 rabbits exhibited widespread immunopositivity. In the other 9 out of 11 rabbits, the kidneys exhibited very sparse immunostaining. Both rabbits with antigen reactivity in the heart and lungs exhibited weakly positive, punctate staining. Three different staining patterns were observed by IHC. The first pattern stained organisms within parasitophorous vacuoles with distinguishable spore morphology (Fig. 16, left). The second pattern exhibited granular or clumped chromogen, extracellular or intracellular to necrotic cells within the lumen of the renal tubules (Fig. 16, middle). The third pattern of staining was granular within the necrotic center of the inflammation (Fig. 16, right).
Detection rates of Encephalitozoon cuniculi in the examined organs using different methods.*
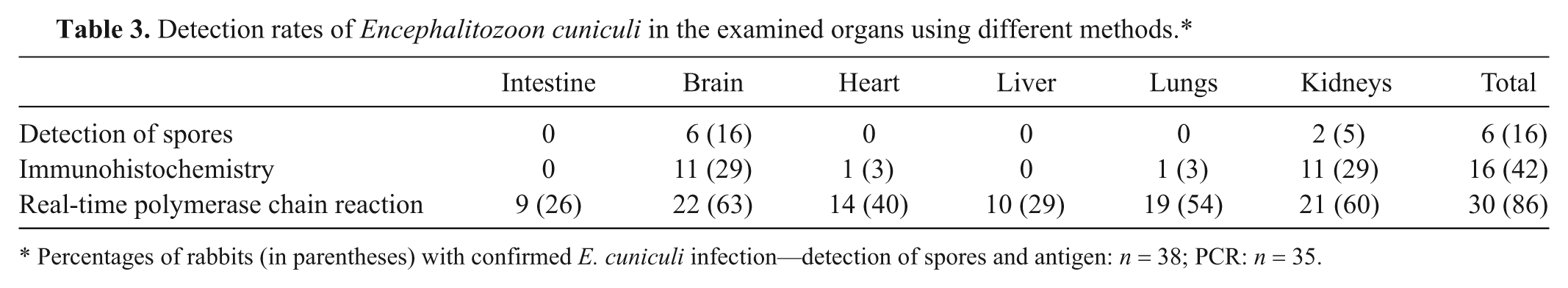
Percentages of rabbits (in parentheses) with confirmed E. cuniculi infection—detection of spores and antigen: n = 38; PCR: n = 35.
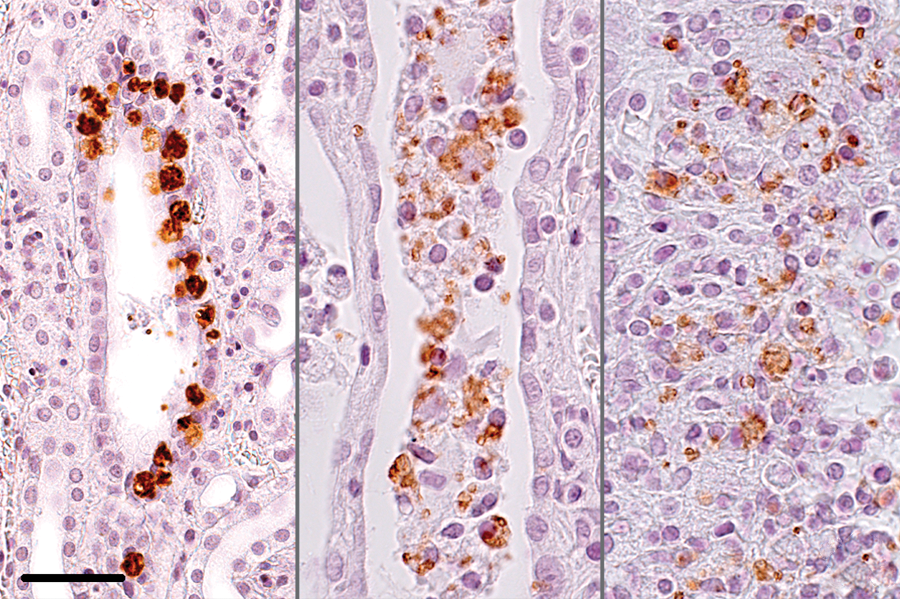
Kidney; rabbit no. 80. Immunoreactivity to Encephalitozoon cuniculi antigen. Left panel: in parasitophorous vacuoles within the tubular epithelium. Bar = 50 μm. Middle panel: free or intracellularly in the tubular lumen. Bar = 25 μm. Right panel: in a center of inflammation. Hematoxylin counterstain. Bar = 25 μm.
Real-time PCR
Tissue samples from 55 rabbits were tested using real-time PCR. These rabbits included 30 that were histopathologically diagnosed with an E. cuniculi infection (in 3 of those cases there was no appropriate material available), 14 without lesions attributable to an E. cuniculi infection, and 11 with possible E. cuniculi infection. In 30 out of 55 rabbits, E. cuniculi DNA was identified in at least 1 of 6 examined organs. The distribution of DNA in examined organs is shown in Table 3. DNA was detected in the brains of 22 rabbits and in the kidneys of 21 rabbits. The lungs were positive in 19 rabbits. In 14 rabbits, the heart was positive, and in 10 rabbits, E. cuniculi DNA was retrieved from the liver. The intestine was positive in 9 rabbits. In 8 rabbits, the kidneys were positive without detectable infection in the brain. In another 8 rabbits, the brain was positive without involvement of the kidneys. In 5 rabbits, DNA was exclusively identified in the brain and kidney. In 1 case, DNA was obtained from the lungs but not from the brain or kidneys.
Comparison of the methods
The results of the different detection methods are provided in Table 4. According to the aforementioned definition, an infection with E. cuniculi was proven in 47% (38/81) of the examined rabbits. In 50% (12/24) of rabbits with clinically presumed encephalitozoonosis, an infection was confirmed. In 41% (33/81) of the rabbits, histological changes associated with E. cuniculi infection were observed, which equated with 87% (33/38) of rabbits with proven E. cuniculi infection. Histology was either inconclusive or inconspicuous in 5 rabbits that subsequently tested positive with real-time PCR. In all 6 cases of histological spore detection, antigen was also detected by IHC. All 16 cases (42%, 16/38) with a positive immunohistochemical result exhibited typical histopathological lesions (granulomatous encephalitis and interstitial nephritis).
Comparison of positive results in the different detection methods used to detect Encephalitozoon cuniculi in rabbits.*
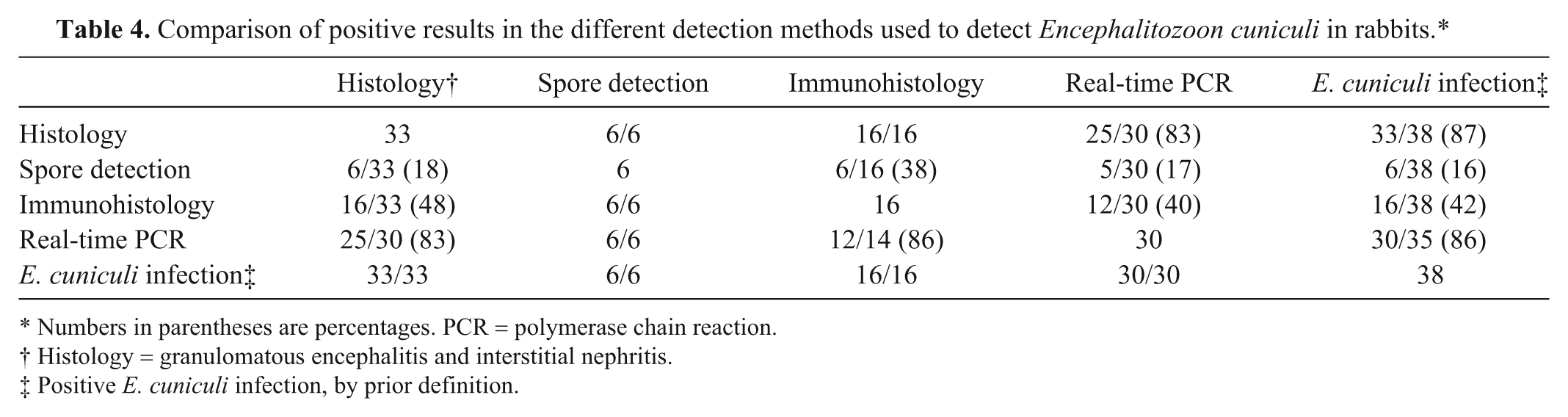
Numbers in parentheses are percentages. PCR = polymerase chain reaction.
Histology = granulomatous encephalitis and interstitial nephritis.
Positive E. cuniculi infection, by prior definition.
In 3 of the 38 infected rabbits, tissue preservation was insufficient to allow PCR testing. In 30 (86%) of the remaining 35 confirmed cases, real-time PCR demonstrated E. cuniculi DNA.
In 83% (25/30) of the rabbits with typical histopathological lesions, tested by PCR, E. cuniculi DNA was detected. In 40% (12/30) of the rabbits with positive PCR results, E. cuniculi antigen was also detected immunohistochemically. Fourteen rabbits with no histopathological indication of E. cuniculi infection were examined by real-time PCR. The PCR was negative in 79% (11/14) of these cases, and PCR was negative in 2 rabbits with a positive immunohistochemical result. One of these rabbits (rabbit no. 24), with 1 single immunopositive spot, exhibited mild inflammatory lesions in the brain. Furthermore, PCR was positive in 2 of the 11 tested rabbits with histologically questionable infection.
Discussion
Encephalitozoon cuniculi is a common cause of neurological disease in pet rabbits. 23 To facilitate the establishment of a gold standard in the diagnosis of E. cuniculi infection, the value of real-time PCR was compared to IHC and conventional morphology. Furthermore, the present study includes a detailed description of histopathological changes in various organs of rabbits infected with E. cuniculi.
The sensitivity of histological detection of E. cuniculi spores, even when using specialized stains, has been perceived differently.7,23,32 Numerous special staining methods to improve the identification of spores in formalin-fixed, paraffin-embedded tissue samples have been described.3,13 In the current study, microscopic evidence of E. cuniculi spores was present in just 16% (6/38) of infected rabbits, despite an extensive search. This finding contrasts with the 87% (33/38) of rabbits exhibiting conclusive histopathological lesions at the tissue level. In a 2009 study, spores were found in all but 1 brain of rabbits with nonsuppurative or granulomatous encephalitis. 6 In the present study, characteristic brain and kidney changes were supported by evidence of spores in 18% of rabbits (6/33), with an emphasis on extralesional localization. As reported in previous studies, the number of parasitophorous vacuoles does not correlate with the severity of histological changes. 7 Intralesional spores were observed quite rarely, which had been explained earlier by extrusion and the possible elimination of the microorganism.6,10
Encephalitozoon cuniculi detection was essentially improved, up to 42% (16/38) in infected rabbits and 48% (16/33) in rabbits with typical histopathological lesions, using antibodies directed against 56-, 45-, and 43-kDa protein epitopes. These results corroborate the findings of a previous study in which antigen was identified in 37.7% of rabbits suspected of E. cuniculi infection. 17 Immunohistochemistry appears to be of particular advantage compared to histological spore detection when there is a low tissue load of the microorganism. 27 Moreover, IHC is favorable for the detection of E. cuniculi in the kidneys, where parasitophorous vacuoles and spores are less prevalent than in the central nervous system. Similar to the histological identification of spores, the immunohistochemical signal is rarely colocalized with histopathological changes. Enceph-alitozoon cuniculi spores and antigen were frequently observed without segregating degenerative or inflammatory tissue changes.5,29
Encephalitozoon cuniculi was detected most frequently in brain and kidneys by both histology and IHC. Other organisms were immunohistochemically detected in the heart and the lungs in a single case each. Spores have also been reported previously to affect liver, spleen, intestine, pancreas, and bone marrow.5,27,29,32 It has been assumed that those localizations are related to an early stage of infection, preceding the elimination of the organism from the respective tissue. 5 It remains unknown why these tissues are apparently spared in recurrent stages of parasitemia, in contrast to kidney and brain. One possible explanation would be cell-bound parasitemia by monocytes. Otherwise, a far more widespread infection of endothelial cells would be postulated.
The brain has been shown to be the most promising organ for the detection of E. cuniculi using morphological and immunohistochemical methods as well as real-time PCR. Accordingly, in a 2009 study using nested PCR, positive detection was more common in the brain than in the kidneys. The heart, lungs, liver, and spleen were only sporadically positive. It was hypothesized that the brain contains the highest concentration of spores because it is the main target organ of E. cuniculi, beside the kidneys. 7 In the present study, E. cuniculi DNA was detected in the brain and in the kidneys at approximately identical frequencies (73% and 70% of rabbits, respectively, with positive PCR results), followed by the lungs (63%). Compared to previous investigations, 7 E. cuniculi DNA was less frequently detected (91.9% vs. 73%) in the brain, and the kidneys were more frequently positive (54.1% vs. 70%). The investigation of the triad brain, kidney, and lung is proposed as a gold standard to obtain the most sensitive PCR results.
Even though the real-time PCR clearly surpassed other methods in the etiologic confirmation of infection (86% of the examined rabbits with E. cuniculi infection compared to 42% for IHC and 16% for histology), 5 PCR-negative rabbits were found with characteristic histopathological lesions in brain and kidneys. Two of these rabbits stained immunohistochemically positive. This discrepancy may be attributed to a low concentration and/or nonuniform distribution of spores in the examined tissue. 7
Notably, E. cuniculi DNA was retrieved from kidneys with predominantly chronic lesions in which spores were not identified or localized by histology or IHC. These results contrast with previously published findings stating that the sensitivity of DNA detection (using nested PCR) correlates with the sensitivity of IHC.7,17 This discrepancy may be explained by the advantages of real-time PCR, including higher specificity, a reduced risk of contamination, and higher sensitivity compared to gel-based PCR. 9
The present findings document the causative organisms’ persistence throughout chronic stages of infection and explain the recurrent, discontinuous shedding of E. cuniculi in urine upon reactivation following immunosuppression.3,28 Likewise, the type of tissue pathology in the kidneys and brain indicate a persistent infection due to the same type of lesions, or recurrent waves of infection, as indicated by concurrent evidence of acute and chronic lesions.
Traditionally, the coexistence of granulomatous encephalitis and interstitial nephritis has been considered diagnostic for E. cuniculi infection because of its consistent and characteristic pattern.10,21 In the current study, the histological manifestation of encephalitozoonosis was similar to previously reported lesions in rabbits with natural or experimental E. cuniculi infections.5,6,21,29
Extensive acute multifocal necrosis of tubular epithelial cells and inflammatory lesions in the kidneys, as reported in early stages of infection, were rare in the investigated samples.10,29 Instead, most rabbits presented with chronic renal lesions with chronic inflammation and fibrous scarring resulting from tubular epithelium necrosis that resulted from the rupture of parasitophorous vacuoles, as described by others.3,10
According to experimental infection studies, following cerebral inoculation of the agent, the type and severity of brain lesions depend on the stage. After 6 weeks, lesions consisted of focal gliosis intermingled with a few scattered lymphocytes. At 8 and 10 weeks, the histological picture was composed of early granulomas with gliosis, lymphocytes, plasma cells, and epithelioid cells. 29 Other studies have described mature granulomas at advanced stages composed of epithelioid cells, lymphocytes, and plasma cells, occasionally accompanied by multinucleated giant cells and centralized necrosis.20,21
The occurrence of a variety of histological lesions coexisting in the brain at different maturation stages was confirmed by the current study. The panel of focal lesions was quite consistent and allowed for the distinction of 6 subtypes (I–VI), distinguished by the level of parenchymal destruction and the reaction pattern of inflammatory cells and astroglia. Type I–III lesions correspond to rather early inflammatory changes. In lesion type IV, the granulomatous character of the inflammation becomes obvious, while in type V the acute inflammatory reaction disappears completely. The end stage of the inflammatory process is reflected in type VI, which is dominated by astroglial scarring. This longitudinal lesion development is consistent with the impact of the E. cuniculi life cycle on the host’s cell integrity, with immunogenic consequences. Evidence of the different lesion types side by side in the brain is indicative of recurrent infection in brain cells, possibly due to recurring parasitemia (e.g., as a result of ruptured parasitophorous vacuoles in the endothelium). Alternatively, the dynamics of the parasite’s intracellular life cycle, and the rupture of parasitophorous vacuoles, may progress at different velocities depending on the host cell’s metabolism and size. In addition, macrophageal uptake may lead to the failed elimination of infective spores and cell-bound spread within brain tissue.3,8,34 Finally, quiescent lesions of types V and VI strongly indicate the possibility of local elimination of the microorganism, but the ability to remove all spores from body is to be questioned.
Previous studies on the spatiotemporal characteristics of central nervous system lesions caused by E. cuniculi have reported minor variations in their results, although there is general agreement that the cerebrum is the most commonly affected region.6,21,28 The current study was intended to elucidate further sublocalizations. Indeed, a regional preference for the innermost layers of the neocortex in E. cuniculi brain infection was demonstrated more concisely than in previous reports. This distribution corresponds to differences in perfusion rates and the density of brain capillaries that the organism may use to enter the brain directly, via their endotheliotropism, or carried by monocytes capable of traversing the blood brain barrier.11,16,33 In addition to the kidneys and brain, nonsuppurative and infrequently granulomatous lesions were reported in other organs, such as liver, lungs, and heart, in connection with E. cuniculi infection.5,12,21,30 Previous reports were confirmed by the present findings. The occurrence of histopathological lesions and the detection of E. cuniculi DNA by real-time PCR in these organs confirm the association of the lesions with this organism. However, compared to kidneys and brain, the other tissues were less frequently lesioned, supporting the stated predilection for the brain and kidneys in encephalitozoonosis.
In experimental infections with E. cuniculi, lungs, kidneys, and liver were found to be involved during acute infection. Brain and gut were unaffected, and the heart was rarely involved in this stage of infection. At chronic stages, the infection mainly affected the brain and kidneys, whereas lesions in heart, lungs, and liver were rarely found and were of minor severity. 5
Other authors 32 have suggested that encephalitozoonosis is a systemic disease, on account of the detection of E. cuniculi in numerous organs. The lymphoplasmacytic infiltrates in the lungs, the liver, and the heart of infected rabbits, as described in the present study, are interpreted as an unspecific immunoreaction due to infection with E. cuniculi. 28 Other authors24,26 have suggested that these infiltrates represent a response to direct damage of the endothelium by the microorganism.
In conclusion, real-time PCR was the most sensitive method for the confirmation of E. cuniculi infection, followed by immunohistology and histological spore detection. Histological observation of the organism in routine sections is most likely in the brain and rare in kidneys. It was not possible to identify E. cuniculi in sections from other tissues. Similarly, brain and kidneys were of the highest diagnostic value for detection of E. cuniculi by IHC. Brain, kidneys, and lungs were all are adequate for E. cuniculi detection using real-time PCR. According to the results of the current study, the gold standard for confirmation of the pathogenic role of E. cuniculi infection would be a combination of histological investigation of predilection sites with real-time PCR performed on the above-mentioned tissues.
Footnotes
Acknowledgements
The authors would like to thank Didier Raoult and Michel Drancourt, Méditerranée Infection, Marseille, France for providing the monoclonal antibodies against E. cuniculi.
a.
Anti-E. cuniculi monoclonal antibodies EC8C12, EC11C5 and EC10G4, Unité des Rickettsies, Faculté de Médecine, Marseille Cedex, France.
b.
Normal goat serum, MP Biomedicals S.A.S., Illkirch, France.
c.
Biotinylated goat anti-mouse Ig, Dako Denmark A/S, Glostrup, Denmark.
d.
Avidin-biotin-complex, Vector Laboratories Ltd., Orton Southgate, Peterborough, UK.
e.
DAB buffer tablets, Merck KGaA, Darmstadt, Germany.
f.
HistoGreen, Linaris GmBH, Wertheim-Bettingen, Germany.
g.
QIAamp DNA Blood and Tissue Kit, Qiagen GmBH, Hilden, Germany.
h.
Thermo Electron GmBH, Ulm, Germany.
i.
HotStarTaq Plus DNA Polymerase Kit, Qiagen GmBH, Hilden, Germany.
j.
Bovine serum albumin, New England Biolabs GmBH, Frankfurt am Main, Germany.
k.
dNTPs, Carl Roth GmbH + Co. KG, Karlsruhe, Germany.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.