
Abstract
Researchers have proposed the adoption of 3 distinct genetic taxa among bacteria previously classified as Edwardsiella tarda; namely E. tarda, E. piscicida, and a taxon presently termed E. piscicida–like. Individual real-time polymerase chain reaction (qPCR) assays were developed, based on published primers, for E. tarda, E. piscicida, and E. piscicida–like sp. to provide rapid quantitative confirmatory tests for these phenotypically ambiguous bacteria. The qPCR assays were shown to be repeatable and reproducible, with high degrees of sensitivity and specificity. Each assay showed a linear dynamic range covering 8 orders of magnitude and a sensitivity limit of 5 copies of target DNA in a 15-µL reaction. In addition, each assay was found specific to their respective targets with no observed amplification from nontarget organisms, including the closely related E. ictaluri and E. hoshinae. Under the conditions used in this study, the 3 assays had a quantifiable limit ranging from 103 (E. piscicida) to 102 (E. piscicida–like and E. tarda) colony forming units in kidney tissue biopsies (approximately 25 mg), pond water samples (35 mL), and broth culture (20 μL). In experimental challenges, the assays were able to detect their respective targets in both clinically and subclinically infected channel catfish (Ictalurus punctatus) fingerlings. In addition to quantifying target bacteria from various substrates, the assays provide rapid identification, differentiation, and confirmation of the phenotypically indistinguishable E. tarda, E. piscicida, and E. piscicida–like sp., a valuable tool for diagnostic assessments.
Introduction
First recognized in the late 1960s, 7 Edwardsiella are a diverse group of enteric Gram-negative bacteria, infecting a wide range of piscine, avian, reptilian, and mammalian hosts.16,20,21 Until 2013, the genus consisted of only 3 taxa: E. tarda, E. ictaluri, and E. hoshinae. 2 There is a dearth of information regarding E. hoshinae, which is primarily considered a pathogen of birds and reptiles.14,29 By comparison, E. ictaluri is well studied. Principally considered a pathogen of cultured channel catfish (Ictalurus punctatus), E. ictaluri can also infect other fish species.8,17,22 Recent reports have implicated E. ictaluri in mortality events outside the farm-raised catfish industry of the southeastern United States, suggesting a more cosmopolitan distribution than previously thought.15,28,31
Edwardsiella tarda is the most widespread and diverse member of the genus, infecting a wide range of hosts from a variety of ecological niches. 21 Primarily thought of as a pathogen of marine and freshwater fish, E. tarda has been demonstrated to have extensive genetic and phenotypic diversity. In 2012, a comparative phylogenomic study of Edwardsiella isolates identified 2 genetically distinct groups (EdwI and EdwII) among organisms phenotypically classified as E. tarda. 30 The work was supported by multilocus sequence analysis of 8 different housekeeping genes, which identified 2 distinct genetic taxa (E. tarda–like and ET883-like) among E. tarda isolates from Europe and Asia. 1 Concurrently, researchers in the United States proposed the existence of 2 genetic taxa within E. tarda, based on the existence of 2 distinct genetic groups (DNA group I and DNA group II) from fishes in the southeastern United States. 12 In 2013, based on comparative phylogenomics, 30 multilocus sequence analysis 1 and DNA–DNA hybridization experiments, the taxon E. piscicida was adopted. 2
A 2014 survey of Edwardsiella isolates from diseased catfish in Mississippi demonstrated that E. piscicida was more commonly associated with disease case submissions of farm-raised catfish than E. tarda or E. piscicida–like sp. 13 Species-specific polymerase chain reaction (PCR) assays targeting the fimbrial gene cluster were developed for each individual taxa and were demonstrated specific to their respective target organisms.13,26 In the current study, real-time PCR (qPCR) assays were developed using these established primer sets and were validated for the detection and quantification of E. tarda, E. piscicida, and E. piscicida–like sp. from catfish kidney tissues, pond water, and broth culture.
Materials and methods
Bacterial cultures and isolation of genomic DNA
The Edwardsiella strains used in the validation of the assays in the current study were characterized as part of an earlier study 13 and identified by gyrB sequencing and species-specific PCR (Table 1). In addition, an Edwardsiella hoshinae strain (ATCC 35051), an Escherichia coli strain (ATCC 25922), 2 Flavobacterium columnare strains (94-081 and ATCC 49512), and 2 Aeromonas hydrophila strains (ML 09-119 and TN 97-08), including a highly virulent strain (ML 09-119) attributed to disease outbreaks in farm-raised catfish 18 were also included in the validation process. Bacteria had been maintained at −80°C in brain–heart infusion (BHI) broth a supplemented with 20% (v/v) glycerol. Frozen cultures were streaked onto Mueller–Hinton agar a plates supplemented with 5% sheep blood (blood agar plates) and allowed to incubate for 24 hr at 37°C (E. coli, E. piscicida, E. piscicida–like, and E. tarda), 24 hr at 28°C (A. hydrophila), or 48 hr at 28°C (E. hoshinae, E. ictaluri, and F. columnare). Individual colonies were picked for each isolate and expanded overnight in BHI broth a at 28°C (A. hydrophila, E. hoshinae, E. ictaluri, F. columnare) or 37°C (E. coli, E. piscicida, E. piscicida–like, E. tarda), respectively. Genomic DNA (gDNA) was isolated from the cultured bacteria using a commercial kit, b following the manufacturer’s suggested protocol for Gram-negative bacteria, resuspended in 100 µL of a commercial DNA hydration solution c (DHS), and quantified spectrophotometrically. d
Specificity of each Edwardsiella real-time polymerase chain reaction (qPCR) assay for the respective target.*
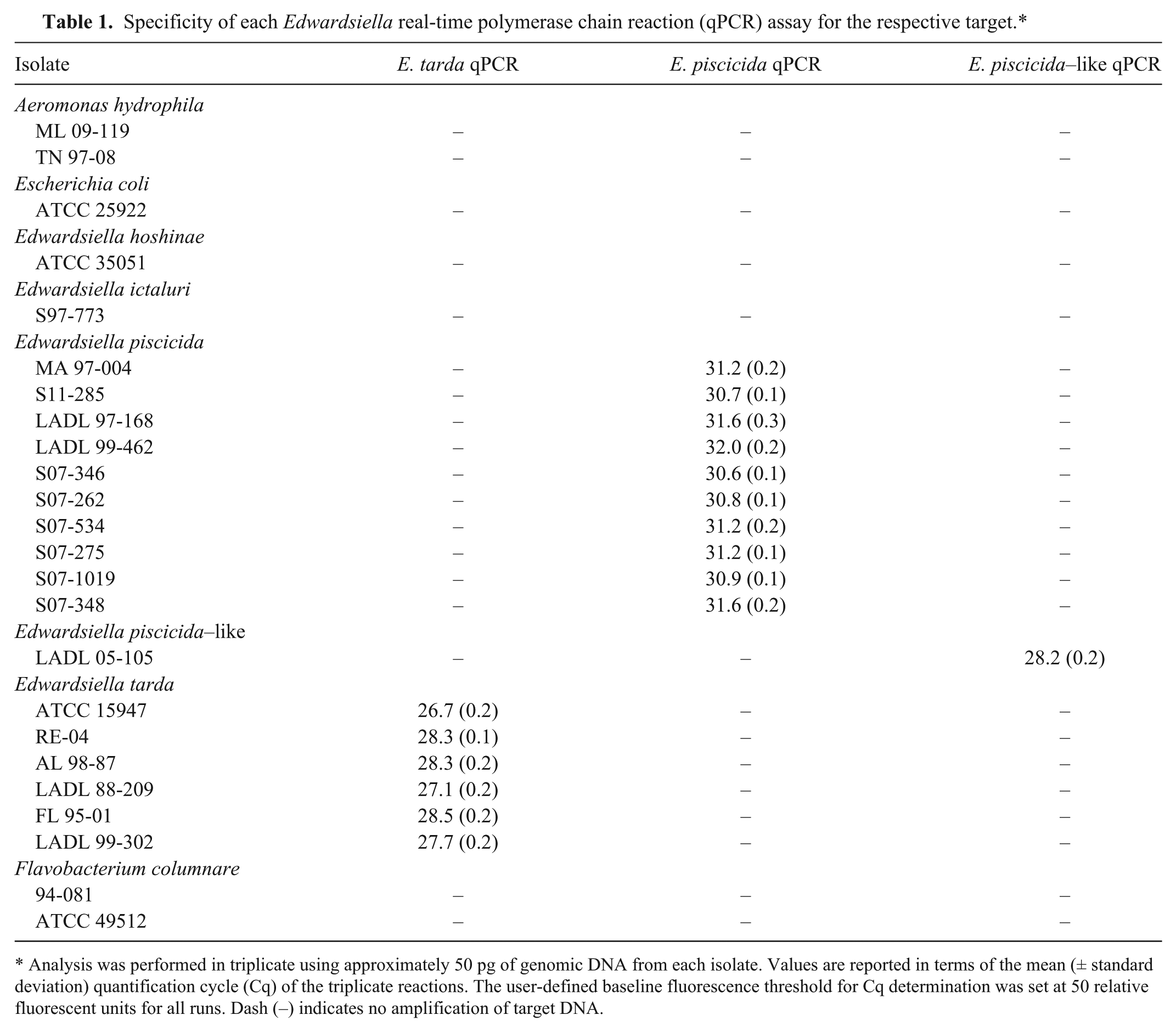
Analysis was performed in triplicate using approximately 50 pg of genomic DNA from each isolate. Values are reported in terms of the mean (± standard deviation) quantification cycle (Cq) of the triplicate reactions. The user-defined baseline fluorescence threshold for Cq determination was set at 50 relative fluorescent units for all runs. Dash (–) indicates no amplification of target DNA.
Design of primer and probe sets
The development of the qPCR assays specific to E. tarda, E. piscicida, and E. piscicida–like sp. was based on previously published PCR primers.13,26 Oligonucleotide probes corresponding to each primer set were designed using primer design software 25 and synthesized commercially. e Each probe was labeled with the fluorescent reporter dye, 6-carboxyfluorescein, on the 5′-end, and the quencher dye, black hole quencher-1, on the 3′-end. Sequences and other relevant information for each primer and probe set can be found in Table 2.
Real-time polymerase chain reaction primers and probes used in the current study.*

Each oligonucleotide probe was labeled with the fluorescent reporter dye, 6-carboxyfluorescein, on the 5′-end, and the quencher dye, black hole quencher-1, on the 3′-end. Melting temperatures (Tm) for oligonucleotide primers and probes were calculated using the default parameters of Primer3. 25 GC = guanine and cytosine content; F = forward; R = reverse; P = probe.
Generation of PCR standards
Standards for relative quantification of target DNA were generated from purified PCR products. Briefly, for each assay, target PCR amplicons were produced from gDNA isolated from E. tarda (ATCC 15947), E. piscicida (S11-285), and E. piscicida–like (LADL 05-105) isolates following published protocols. 13 To confirm the presence of a single, appropriately sized band, amplicons were visualized under ultraviolet light after electrophoretic passage through agarose in the presence of ethidium bromide (1 µg/mL). Band sizes were estimated by concurrent passage of a molecular weight marker. f Last, amplicons were purified using a commercial PCR purification kit, g resuspended in DHS, and quantified spectrophotometrically. d
Quantitative PCR
The 15-μL PCR reactions contained 8 μL of PCR master mix, h 10 pM of each primer, 1 pM of probe, DNA template, and nuclease-free water to volume. Amplifications were performed on a qPCR system i programmed for 1 cycle of 95°C for 15 min followed by 40 cycles of 95°C for 15 sec and 60°C for 1 min. Data collection was carried out following the 60°C annealing/extension step at the end of each cycle. For each plate, samples, as well as no-template negative controls, were run in triplicate.
PCR specificity, sensitivity, repeatability, and reproducibility
The specificity of each assay was tested against both target and nontarget DNA. Genomic DNA (approximately 50 pg) from E. tarda, E. piscicida, E. piscicida–like, E. ictaluri, E. hoshinae, A. hydrophila, F. columnare, and E. coli were analyzed in triplicate using the reaction conditions and thermal cycling parameters described above. Quantification cycles (Cq) for each reaction were based on a user-defined baseline threshold of 50 relative fluorescent units (RFUs).
The sensitivity and linear dynamic range of each assay was determined using serial dilutions of known quantities of target DNA (purified PCR amplicons), ranging from 1 to 1 × 108 copies of target DNA per 15-μL reaction. Each dilution series was run in triplicate on 3 separate occasions to assess repeatability and reproducibility of the assay. The Cq for each reaction was based on a user-defined baseline threshold of 50 RFUs.
Detection of target DNA from broth culture, fish tissue, and pond water
The ability of each assay to detect target DNA from different substrates was evaluated. Known quantities of E. tarda, E. piscicida, and E. piscicida–like sp. were added to catfish kidney biopsies, pond water, or processed directly in broth culture. Initially, cryostocks of E. tarda (ATCC 15947), E. piscicida (S11-285), and E. piscicida–like sp. (LADL 05-105) were streaked for isolation, and individual colonies were grown in 9 mL of BHI broth for 5 hr at 37°C without agitation. After a 10-fold serial dilution, plate counts were performed in triplicate (drop-plate method) on blood agar plates using 20-µL aliquots from each dilution. Additional 20-µL aliquots from each dilution were transferred to 1.5-mL microcentrifuge tubes and stored at −80°C until processing. These cryogenically stored aliquots corresponded to aliquots used for plate counts and represented known quantities of bacteria that could be added to water samples and catfish kidney biopsies. Three aliquots from each dilution were analyzed for each substrate (pond water, catfish kidney tissue, and BHI broth) representing colony forming unit (CFU) equivalents ranging from 1 to 1 × 106 CFUs per 20-μL aliquot. Isolation of gDNA from bacteria in broth culture was carried out using a commercial kit, b following the manufacturer’s suggested protocol for Gram-negative bacteria. Isolated gDNA was resuspended in 100 µL of DHS, and 5 µL of gDNA suspension from each aliquot was used in qPCR analysis. Each aliquot was run in triplicate on a plate containing no-template controls (run in triplicate) and standard positive controls (run in duplicate). Positive controls consisted of purified and quantified PCR amplicons, ranging from 5 to 5 × 105 copies of target DNA.
To evaluate the ability of the assays to detect target organisms in fish tissues, cryogenically stored aliquots of known quantities of bacteria were added to posterior kidney biopsies (approximately 25 mg) collected aseptically from channel catfish reared for disease research at the holding facility of the Thad Cochran National Warmwater Aquaculture Center (TCNWAC; Stoneville, Mississippi). Initially, catfish kidney tissues were confirmed negative for Edwardsiella spp. by culture and by qPCR using the assays described herein. Three aliquots from each dilution were added directly to individual kidney tissue samples prior to homogenization. Genomic DNA from spiked kidney tissues was isolated using a commercial kit, b following the manufacturer’s suggested protocol for animal tissues. The isolated gDNA was resuspended in 200 µL of DHS, and 5 µL of gDNA from each aliquot was used as template in qPCR analysis. As above, each aliquot was run in triplicate on a plate containing no-template controls (run in triplicate) and concurrently run standard positive controls (run in duplicate).
Similarly, to determine the ability of the assays to detect and quantify target organisms in catfish pond water, aliquots of known quantities of bacteria were added to pond water samples. Prior to the addition of bacteria, pond water used for this analysis was confirmed negative for Edwardsiella spp. by qPCR. Based on previously established protocols for the detection of bacteria in catfish pond water,9,10 a water sample (20 L) was collected from a commercial catfish pond and processed within 24 hr of collection. A subsample (35 mL) of the pond water was added to a 40-mL round-bottom centrifuge tube and centrifuged at 20,000 × g for 10 min. The supernatant was removed, and the pellet was resuspended in 1.5 mL of nuclease-free water and transferred to a 1.8-mL microcentrifuge tube. A 20-µL aliquot from each broth culture dilution was added directly to each pellet, and DNA isolation was carried out using a commercial kit, j following the manufacturer’s suggested protocol for wet samples. Isolated DNA was resuspended in 100 µL of elution buffer, k and 5 µL was used in each individual qPCR, carried out as above. For the purposes of calculating averages throughout the study, negative reactions were assigned an RFU of 0, Log10 starting quantities of 0.0, and Cq values of 40.0.
Detection in experimentally infected fish
The ability of the assay to detect target DNA in clinical and subclinical, experimentally infected fish was evaluated. Channel catfish fingerlings (mean weight: 21.9 g; range: 12.8–30.2 g) were obtained from the TCNWAC fish-rearing facility. For the challenge, 30 fish were placed in twenty 80-L aquaria containing 20 L of well water and held under flow-through conditions (1 L/min) with constant aeration. Bacterial cultures of E. piscicida, E. piscicida–like, and E. tarda were grown as described above. Two tanks of channel catfish fingerlings were anesthetized with tricaine methanesulfonate (MS-222) m and injected intraperitoneally with one of the treatments (i.e., 1 dilution of 1 of the 3 bacteria): E. piscicida (1.83 × 105, 1.83 × 106, and 1.83 × 107), E. piscicida–like (1.33 × 105, 1.33 × 106, and 1.33 × 107), and E. tarda (2.92 × 105, 2.92 × 106, and 2.92 × 107). The remaining 2 tanks were negative controls that were handled similarly but injected intraperitoneally with sterile BHI broth. For each treatment, 1 tank was designated for sampling (sampling tank) and 1 tank (mortality tank) was used to estimate the median lethal dose (LD50) of each bacterial strain based on the number of dead fish observed for each dose. 24 The mortality tank was checked twice daily, and the number of dead fish was recorded. Apparently healthy fish (n = 3), with no outward signs of disease, were collected from the sampling tank at 1, 2, 5, and 7 days postinjection. In addition, dead fish observed in the sampling tanks were also collected and processed for qPCR. All sampled fish were euthanized using MS-222, and posterior kidney biopsies (approximately 25 mg) were obtained aseptically, streaked on blood agar plates, and incubated for 24 hr at 37°C to determine the presence of viable bacteria. Genomic DNA was isolated from the kidney biopsies using a commercial kit, b as above. The isolated gDNA was resuspended in 200 µL of DHS, and 5 µL of gDNA was used in each individual qPCR, carried out as above.
Results
PCR specificity, sensitivity, repeatability, and reproducibility
Each qPCR assay demonstrated robust amplification from gDNA isolated from their respective targets, with no amplification from gDNA isolated from nontarget organisms (Table 1). Using 10-fold serial dilutions of PCR amplicons, each assay was linear over 8 orders of magnitude and sensitive to an estimated 5 copies of target DNA (Fig. 1). Reactions with <5 copies of target DNA resulted in inconsistent amplification, often with no observed signal. Throughout the study, reaction efficiencies were calculated from the slope of the log-linear portion of concurrently run standards (PCR efficiency = 10−1/slope − 1) 5 and were within the generally accepted range of 90–110% (E. piscicida, range: 91.3–98.0%, mean: 94.8%; E. piscicida–like sp., range: 94.7–105.3%, mean: 100.6%; E. tarda, range: 101.8–107.9%, mean: 104.4%).
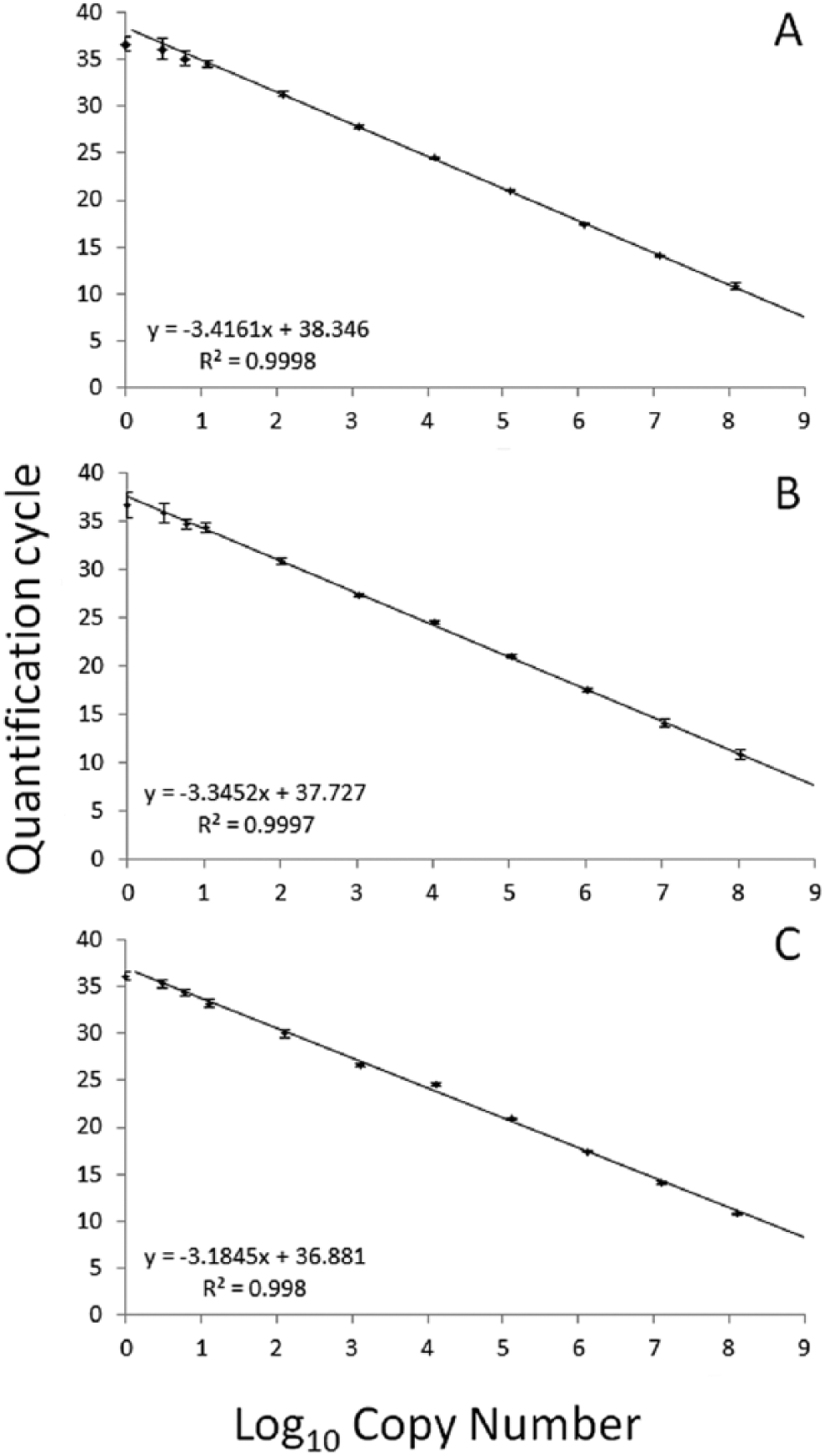
Mean quantification cycles (Cq) for known quantities of Edwardsiella piscicida (
Clinical sensitivity and assay variability
Each assay detected target DNA in gDNA preparations from approximately 100 CFU per sample from broth culture. For samples with <100 CFU, amplification was inconsistent between replicates and occasionally absent, with a proportion (44.4% for E. tarda; 77.7% for E. piscicida; 44.4% for E. piscicida–like sp.) of reactions from aliquots of <100 CFU per sample giving negative results. When present, amplification resulted in Cq values similar to those observed for approximately 100 CFU, demonstrating a plateau effect common near the quantifiable limits of qPCR assays. In addition, several (55.5%) reactions corresponding to aliquots of 136 CFU for E. piscicida were negative, suggesting this quantity was at or below the limits of the quantifiable or detectable range of the assay. Results from pond water and catfish kidney samples spiked with known quantities of target bacteria were similar to results obtained from broth culture, although kidney samples containing <100 CFU resulted in inconsistent amplification between replicates, with a proportion (44.4% for E. tarda; 33.3% for E. piscicida; 44.4% for E. piscicida–like sp.) of reactions giving negative results. Again, several (33.3%) reactions corresponding to aliquots of 136 CFU for E. piscicida were also negative. Each assay was linear over at least 5 orders of magnitude in these experiments and, under the conditions used in this study, the 3 assays had a quantifiable limit ranging from 103 (E. piscicida) to 102 (E. piscicida–like and E. tarda) CFU in kidney tissue biopsies (approximately 25 mg), pond water samples (35 mL), and broth culture (20 μL; Fig. 2; Table 3).
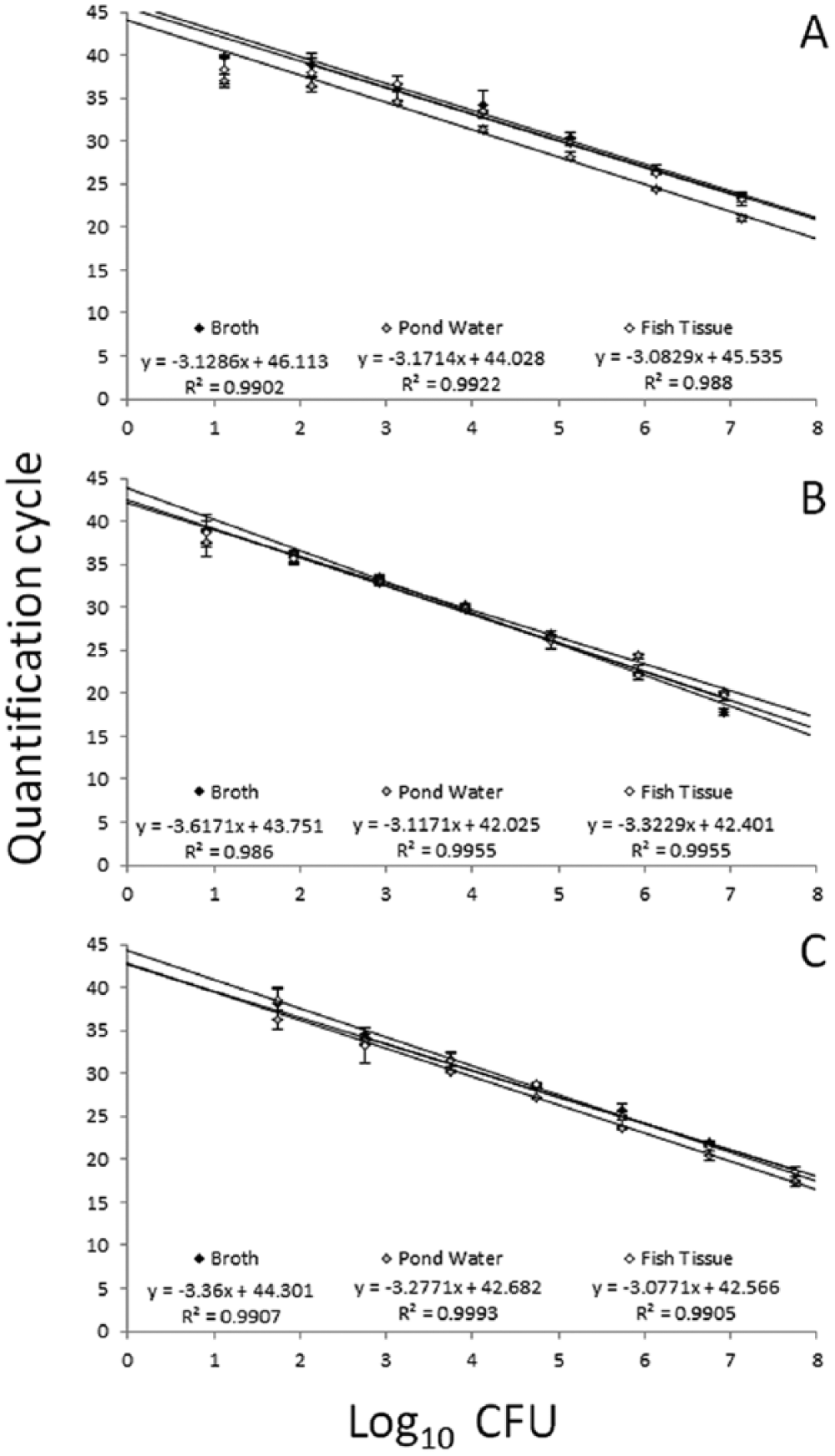
Mean quantification cycles (Cq) for known quantities of Edwardsiella piscicida (
Mean (± standard deviation) quantification cycles (Cq), Log10 copy number, and relative fluorescent units (RFU) for known quantities of Edwardsiella piscicida, Edwardsiella piscicida–like sp., and Edwardsiella tarda cells in brain–heart infusion (BHI) broth, pond water, or channel catfish (Ictalurus punctatus) posterior kidney tissue.*
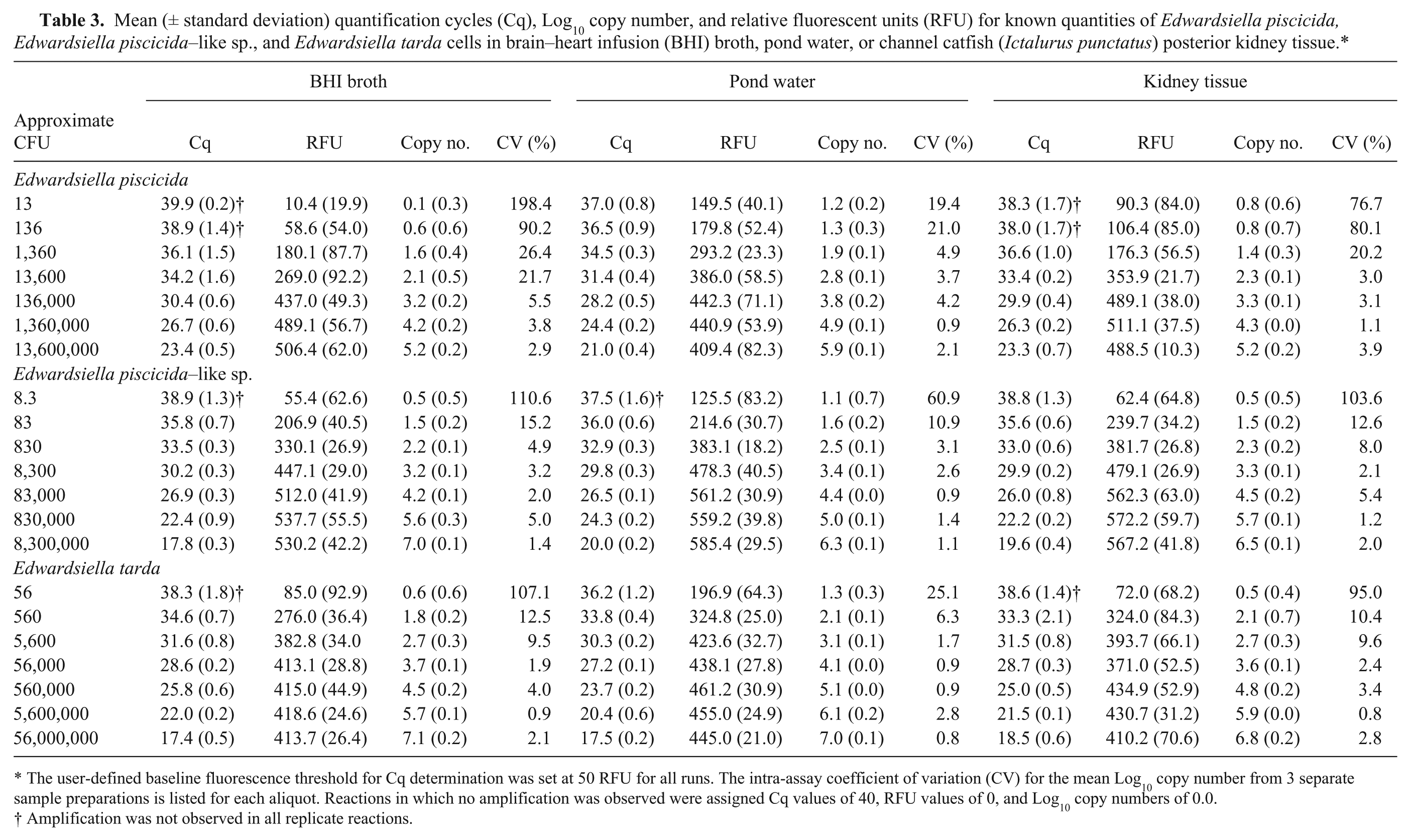
The user-defined baseline fluorescence threshold for Cq determination was set at 50 RFU for all runs. The intra-assay coefficient of variation (CV) for the mean Log10 copy number from 3 separate sample preparations is listed for each aliquot. Reactions in which no amplification was observed were assigned Cq values of 40, RFU values of 0, and Log10 copy numbers of 0.0.
Amplification was not observed in all replicate reactions.
Detection in experimentally infected fish
The qPCR assays consistently amplified target DNA from apparently healthy, subclinically infected fingerlings in all experimental treatment groups up to 5 days postinjection. Large quantities of target DNA were detected from dead or moribund fish clinically infected with E. piscicida and E. tarda (Table 4), often equating to 4 or more orders of magnitude above the clinical sensitivity of the assay. In addition, target bacteria were confirmed by qPCR from culture in 97% (31/32 fish) of E. piscicida mortalities and 100% (7/7 fish) of E. tarda mortalities.
Mean (± standard deviation) quantification cycle (Cq) and Log10 copy number for channel catfish (Ictalurus punctatus) fingerlings experimentally challenged with Edwardsiella piscicida, Edwardsiella piscicida–like sp., and Edwardsiella tarda.*
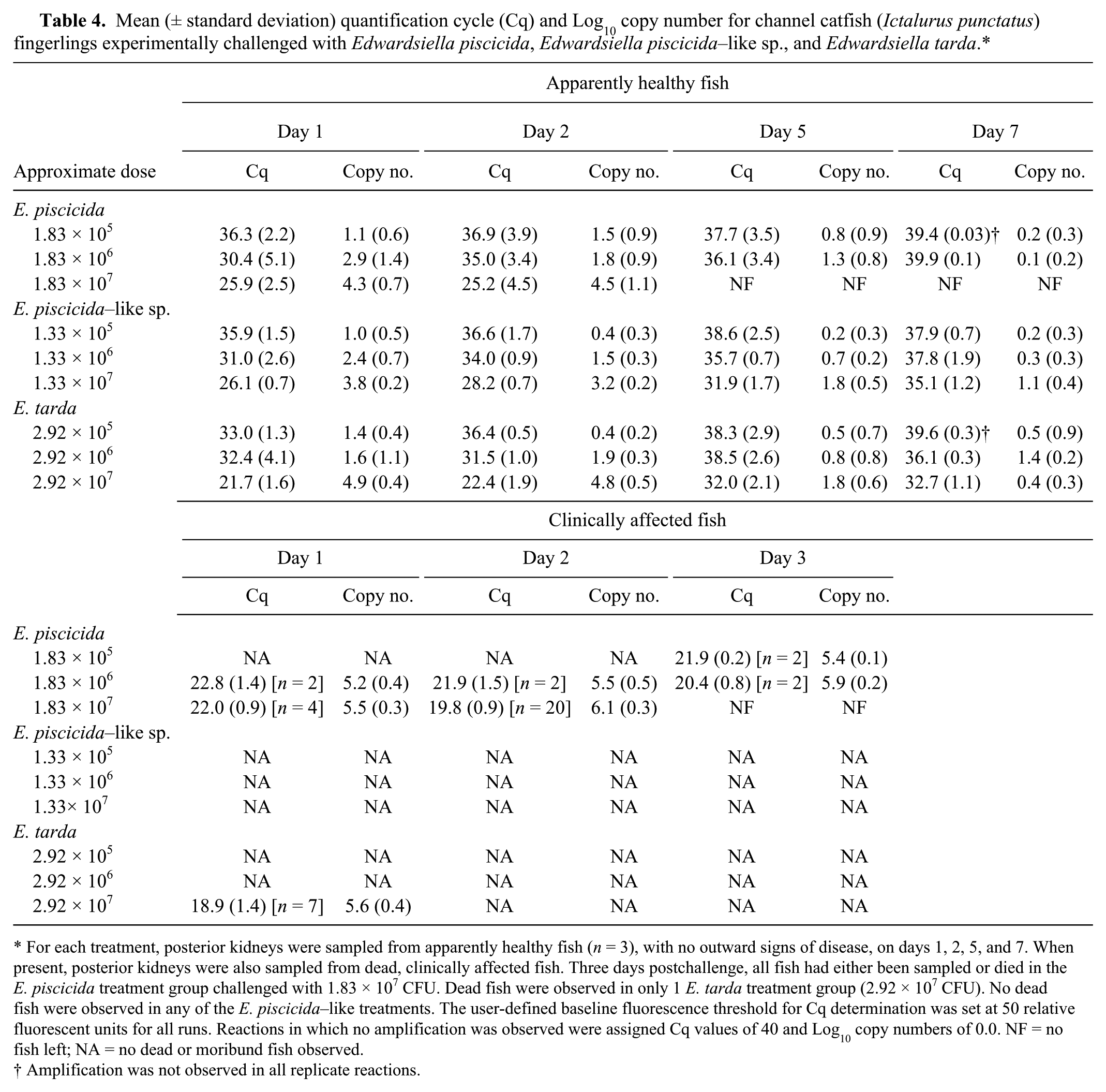
For each treatment, posterior kidneys were sampled from apparently healthy fish (n = 3), with no outward signs of disease, on days 1, 2, 5, and 7. When present, posterior kidneys were also sampled from dead, clinically affected fish. Three days postchallenge, all fish had either been sampled or died in the E. piscicida treatment group challenged with 1.83 × 107 CFU. Dead fish were observed in only 1 E. tarda treatment group (2.92 × 107 CFU). No dead fish were observed in any of the E. piscicida–like treatments. The user-defined baseline fluorescence threshold for Cq determination was set at 50 relative fluorescent units for all runs. Reactions in which no amplification was observed were assigned Cq values of 40 and Log10 copy numbers of 0.0. NF = no fish left; NA = no dead or moribund fish observed.
Amplification was not observed in all replicate reactions.
Regardless of challenge dose or isolate, viable bacteria were recovered from subclinically infected fish up to 5 days postchallenge. Of the apparently healthy, subclinically infected fish, 47% (48/102) did not exhibit bacterial growth on culture, and no viable bacteria were recovered from any fish sampled 7 days postchallenge. For each of the 3 target bacteria, negligible target DNA amplification was observed from several culture-negative fish (mean Cq = 37.2), equating to <3 CFU equivalents (Fig. 2), below the reliable, clinical sensitivity of the assay. Similarly, amplification of target DNA from BHI-injected fish was also negligible, corresponding to quantities below the clinical and analytical sensitivity of the assay (mean Cq: E. tarda, 38.5; E. piscicida, 39.2; E. piscicida–like sp., 39.1).
Based on the cumulative mortalities found 7 days postinjection, the observed LD50 for E. piscicida was 5.77 × 105 CFU. The LD50 for E. tarda and E. piscicida–like sp. could not be determined. Only 17% mortality was observed in fish injected with 2.92 × 107 CFU of E. tarda, with no mortality seen in fish injected with 2.92 × 105 CFU or 2.92 × 106 CFU. Similarly, no mortality was observed in fish injected with E. piscicida–like sp. (Fig. 3), even at doses as high as 1.33 × 107.
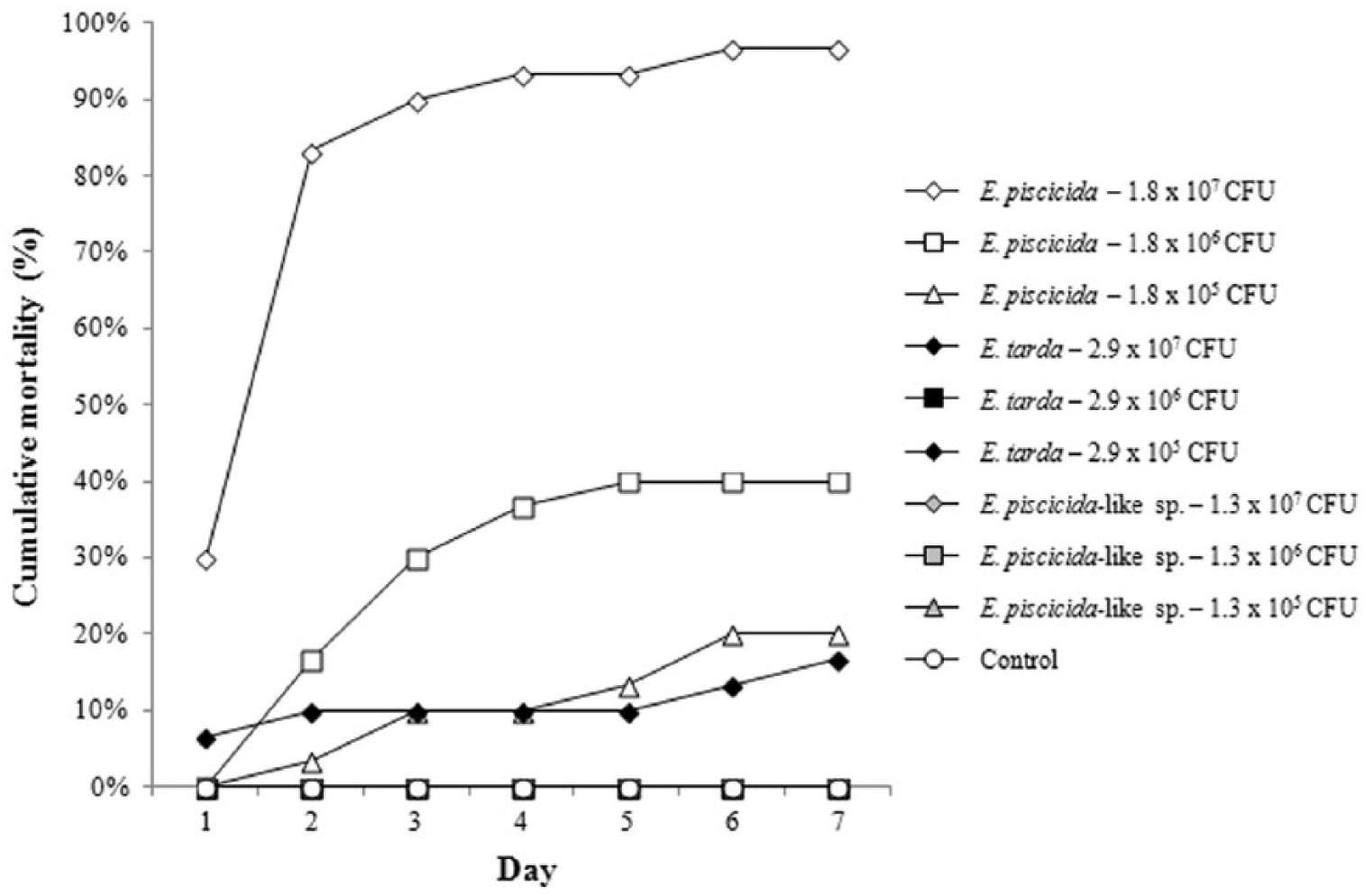
Nonreplicated cumulative mortality for channel catfish (Ictalurus punctatus) challenged with 3 different doses of Edwardsiella piscicida, Edwardsiella piscicida–like sp., and Edwardsiella tarda. Sixty fish were challenged to each dose and distributed into 2 separate aquaria (n = 30 fish/tank). Samples were collected for real-time polymerase chain reaction (qPCR) analysis from one tank (sampling tank) on days 1, 2, 5, and 7 (data presented in Table 4). No samples were collected from the second tank (mortality tank). Rather, the mortality tank was checked twice daily over the course of 7 days and dead fish recorded. The cumulative mortality observed in the mortality tank is reported.
Discussion
Real-time PCR assays are rapidly becoming more commonplace in fish disease work, for both research and diagnostic applications. 23 Assays have been developed for the detection and quantification of a wide array of viral, parasitic, and bacterial fish pathogens in both fish tissues and the environment.3,11,19,27 In catfish aquaculture, several assays have been developed for the detection and quantification of pathogens in the pond environment.9 –11
All 3 assays used in the current study were able to detect target DNA in both clinically and subclinically infected fish. While negligible amplification (Cq values ≥37.0) was observed from some culture-negative fish, it is assumed these high Cq values are artifacts of carryover contamination, amplification of nonspecific or background DNA, or degradation of the fluorescent probe 4 rather than detection of nonviable organisms. While arbitrary cutoffs are not ideal, 5 this inconsistent late amplification represents target copy numbers below the clinical sensitivity of the assay. As such, it likely represents false-positives and, under the conditions described herein, Cq values of ≥37.0 in the absence of viable, cultured organisms should be considered suspect. In addition, due to differences in environmental conditions, dose effect, and other factors not accounted for during laboratory challenges, bacterial loads and qPCR results from naturally infected fish may differ from what is reported here. Future research will focus on establishing the clinical relevance of the values observed during experimental infections and how they relate to natural infections. Lastly, the data further demonstrates the limitations of using qPCR as a stand-alone diagnostic tool. Instead, qPCR is better employed as a research tool or a confirmatory test used in conjunction with identification of pathogens in histological sections or the culture and recovery of viable organisms.
At present, no distinguishing phenotypic trait has been identified to discriminate between E. tarda, E. piscicida, and E. piscicida–like sp.12,13 However, the qPCR assays described herein were demonstrated to be highly reproducible and repeatable, with limited variability between runs. These assays provide rapid identification and differentiation of these phenotypically comparable organisms in both apparently healthy and clinically affected fish, a valuable aide for diagnostic assessments. In addition, the assays provide a means to detect and quantify these Gram-negative pathogens in catfish pond water, providing an invaluable tool for researchers and diagnosticians to evaluate the epidemiology of these organisms in cultured fish.
All surviving experimentally infected fish were culture negative by day 7, regardless of challenge dose or isolate. This is consistent with previous work investigating E. tarda pathogenesis in channel catfish. 6 However, the current study demonstrated an apparent difference in virulence between E. piscicida, E. piscicida–like sp., and E. tarda. This variability warrants further investigation to better understand the pathogenicity of these Edwardsiella spp. in channel catfish.
Historically, E. tarda has been perceived as a pathogen of low occurrence in catfish aquaculture. However, according to summaries of diagnostic case submissions to the Aquatic Research and Diagnostic Laboratory in Stoneville, Mississippi (http://tcnwac.msstate.edu/publications.htm), there has been a relative increased incidence of E. tarda infections in farm-raised catfish in Mississippi over the past decade. Until 2013, all genetic variants of E. tarda were considered to be members of a genotypically diverse, yet phenotypically homogenous species. Research has demonstrated the E. tarda taxa is comprised of at least 2, possibly 3 phenotypically ambiguous, yet genetically distinct taxa, namely E. tarda, E. piscicida, and E. piscicida–like sp.1,2,12,30 A survey of E. tarda isolates collected from diseased fish submissions to the Aquatic Research and Diagnostic Laboratory in Stoneville, MS, from 2007–2012 suggests that E. piscicida is more commonly associated with disease outbreaks in farm-raised catfish than E. tarda. A total of 44 archived isolates were examined, all of which were identified phenotypically as E. tarda on initial isolation. All 44 were identified as E. piscicida by PCR. 13 The factors, both biotic and abiotic, that have contributed to the putative trend of increased incidence of E. piscicida in Mississippi aquaculture are currently unknown. However, these assays used in the current study will provide a valuable tool in identifying the aforementioned factors that may be attributing to this escalation.
Footnotes
Acknowledgements
This study is Mississippi Agricultural and Forestry Experiment Station (MAFES) publication no. J-12543.
a.
BD Diagnostic Systems, Sparks, MD.
b.
DNeasy blood and tissue kit, Qiagen Inc., Valencia, CA.
c.
PureGene DNA hydration solution, Qiagen Inc., Valencia, CA.
d.
NanoDrop ND-2000, Thermo Fisher Scientific, Waltham, MA.
e.
Sigma-Aldrich, St. Louis, MO.
f.
Hyperladder II, Bioline USA Inc., Taunton, MA.
g.
QIAquick PCR purification kit, Qiagen Inc., Valencia, CA.
h.
TaqMan Environmental Mastermix 2.0, Applied Biosystems, Foster City, CA.
i.
CFX96, Bio-Rad Laboratories, Hercules, CA.
j.
Powersoil DNA isolation kit, Mo Bio Laboratories Inc., Carlsbad, CA.
k.
Powersoil solution C6, Mo Bio Laboratories Inc., Carlsbad, CA.
l.
Gentra Puregene DNA purification kit, Qiagen Inc., Valencia, CA.
m.
Western Chemical Inc., Ferndale, WA.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This research was supported by the United States Department of Agriculture Agricultural Research Service (project no. 58-6402-2729), the United States Department of Agriculture Catfish Health Research Initiative (CRIS 6402-31320-002-02), the Mississippi State University College of Veterinary Medicine, and the Mississippi Agricultural and Forestry Experiment Station (MAFES).