
Abstract
A rapid, easy method involving a polymerase chain reaction (PCR) assay followed by restriction fragment length polymorphism (RFLP) analysis was developed to differentiate the 2 phylogenetic lineages of Pasteurella multocida. The PCR targeted the 16S ribosomal RNA gene, and the RFLP involved separate digestions with HindIII, EarI, and MlsI. The method was applied to 16 isolates of P. multocida from different hosts and the isolates were clearly assigned to 1 of the 2 lineages. The phylogenetic lineages did not match with the phenotypic-based identification at the subspecies level.
Pasteurella multocida represents a diverse group of bacteria causing disease in a wide range of hosts, including fowl cholera in birds, hemorrhagic septicemia in ruminants, atrophic rhinitis and bronchopneumonia in swine, snuffle in rabbits, and sporadic wound infections in human beings. 2 Pasteurella multocida shows considerable variety in phenotypic properties5,9,14 causing frequent confusion in the exact identification and definition of this species. DNA-DNA hybridization–based reclassification of the genus Pasteurella Trevisan 1887 led to the recognition of 3 P. multocida subspecies: subsp. multocida, subsp. septica, and subsp. gallicida. 10 On the other hand, multilocus enzyme electrophoresis and sequence analysis of the 16S ribosomal RNA (rRNA) coding gene and certain housekeeping genes (atpD, rpoB, sodA, recN, thdF, and rpoA) suggest the existence of 2 main distinct phylogenetic lineages favoring the combination of the 2 phenotypically and genetically close subspecies (subsp. multocida and gallicida) into 1 new subspecies and leaving P. multocida subsp. septica as a separate subspecies.1,3,6
In routine clinical practice, identification of P. multocida at the species level is usually sufficient. However, the exact definition of subgroups (subspecies and/or phylogenetic lineages) within the species may have taxonomical relevance and may provide useful information for epidemiological investigations. The discrimination of subspecies is rather difficult for practical applications. The range of adequate phenotypic characters (fermentation of dulcitol, D(-)sorbitol, trehalose, and α-glucosidase activity) is still a subject of debate (Bisgaard M, Kuhnert P, Olsen JE, Christensen H: 2005, Investigations on the existence of phenotypic criteria for separation of 16S rRNA clusters A and B of Pasteurella multocida. Pasteurellaceae 2005. In: Proceedings of the ASM conference, p. 45. October 23–26, 2005, Kohala Coast, Hawaii)7,10 because the results of biochemical fermentation tests align poorly if at all to taxonomic or phylogenetic status of the individual strains.4,9,14 The most widely accepted method for differentiation of phylogenetic lineages is DNA sequencing of 16S rRNA gene (Bisgaard et al., 2005), 8 a technique that cannot be performed in every routine laboratory. Therefore, the purpose of the present work was to establish a sequencing-independent molecular differentiation of P. multocida subgroups to replace the expensive and time-consuming sequence analysis approach.
Sixteen strains isolated from various hosts and identified as P. multocida by a species-specific polymerase chain reaction (PCR) 15 were characterized in the current study (Table 1). The strains were stored at −70°C in 20% skimmed milk a from which they were inoculated on Columbia agar b supplemented with 5% sheep blood and incubated under aerobic conditions at 37°C for 24 hr. Genomic DNA was extracted using a commercial kit c as described previously. 12 The 16S rDNA (1235 nucleotide in length) was amplified by PCR carried out in a 50-μl volume that was comprised of 5 μl of DNA template, 12.5 pmol of each modified universal primer d (forward: 5′-AGAGTTTGATCATGGCTCAG-3′, position 9-29; reverse: 5′-CCATTGTAGGACGTGTGAAGCCC-3′, position 1222-1244) in 1× PCR buffer, e 200 nM deoxyribonucleotide triphosphate mix, e and 1.25 U commercial DNA polymerase. e The reactions were run in a thermal cycler f with the following parameters: 4 min at 94°C, followed by 35 cycles of 45 sec at 94°C, 1 min at 55°C, and 45 sec at 72°C, followed finally by extension for 7 min at 72°C. The PCR products were visualized by electrophoresis on a 1.5% agarose gel stained with ethidium bromide.
Identity of the Pasteurella multocida strains used in the current study.
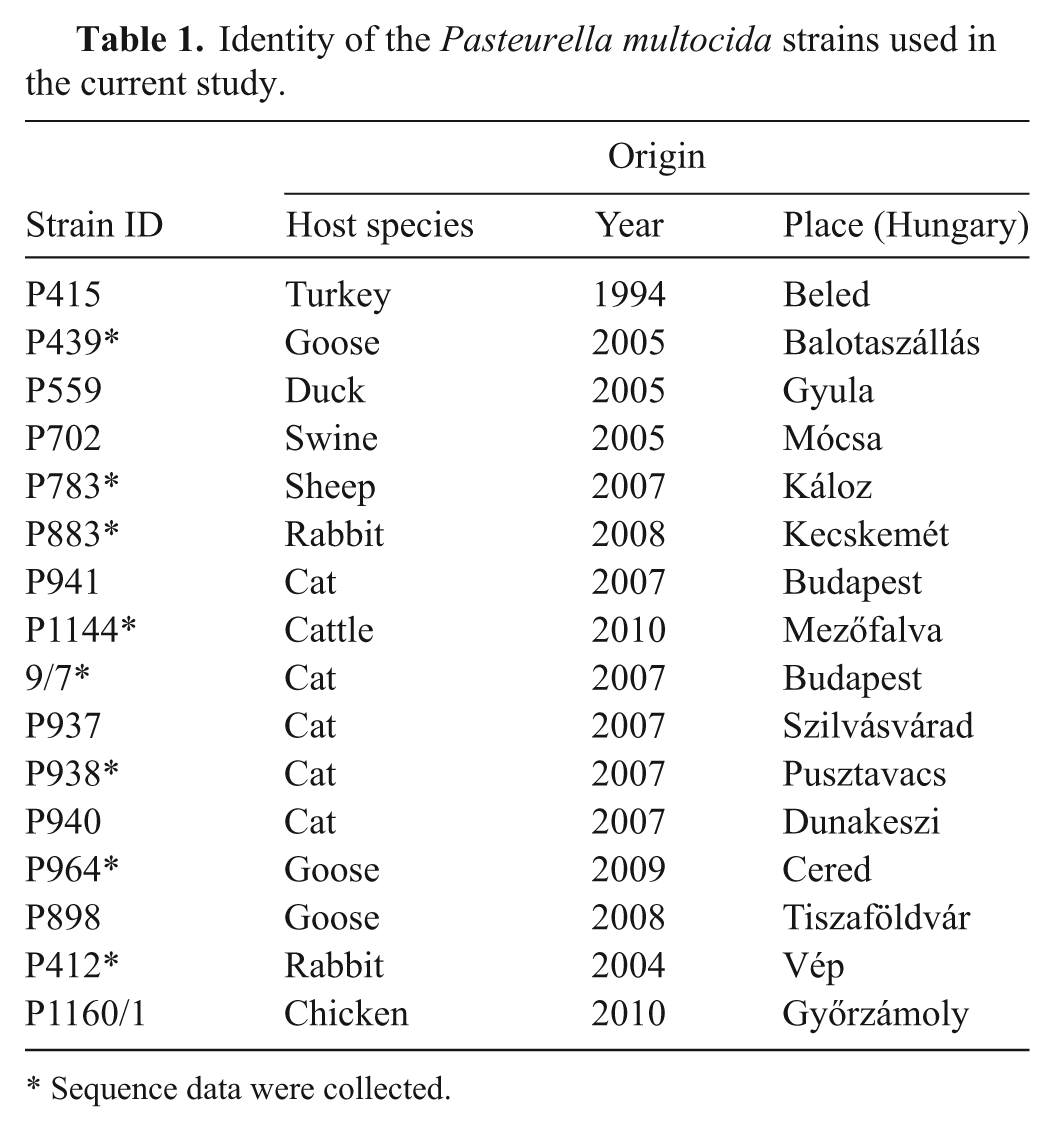
Sequence data were collected.
The comparative analysis of available partial or full-length P. multocida 16S rRNA in GenBank (http://www.ncbi.nlm.nih.gov/genbank) revealed abundant (19) phylogenetically informative nucleotide substitutions corresponding with the results of previous studies4,8 and being suitable to identify phylogenetic lineages (Table 2). These characteristic nucleotide substitutions allowed discrimination between the 2 main lineages by restriction fragment length polymorphism analysis (RFLP). The RFLP procedure was carried out by digesting the amplified PCR products with HindIII, EarI, and MlsI endonucleases e in the supplied buffer according to the manufacturer’s protocols. The RFLP products were run on 2.0% agarose gels at 5 V/cm for 2 hr and 30 min, stained with ethidium bromide, and then analyzed under ultraviolet illumination. DNA fingerprints of the strains were compared by visual inspection of the band patterns.
Conservative nucleotide changes between 16S ribosomal RNA lineages A and B of Pasteurella multocida.*

Sequence alignment of the 16S ribosomal DNA fragments from 4 representative strains within each polymerase chain reaction–restriction fragment length polymorphism group with reference sequence confirms their separation. The numbers above the sequence identify the nucleotide positions (NC_002663 was used from GenBank as reference). The dots represent similar nucleotides. Shaded boxes indicate emerged substitutions resulted restriction enzyme recognition sites.
Pasteurella multocida subspecies type strain.
The amplified 16S rDNA fragments represented approximately 80% of the gene. HindIII cleaved the amplicons into 2 (74 bp and 1,160 bp) or 3 fragments (74 bp, 238 bp, and 922 bp; Fig. 1A), dividing the strains into 2 separate groups (I and II). The use of EarI and MlsI restriction endonucleases provided a different way of distinction of the same 2 lineages: EarI cut fragments (243 bp and 991 bp) only from strains belonging to group I of HindIII, while MlsI cleaved fragments (207 bp and 1,027 bp) only from strains in group II (Figs. 1B, 1C).
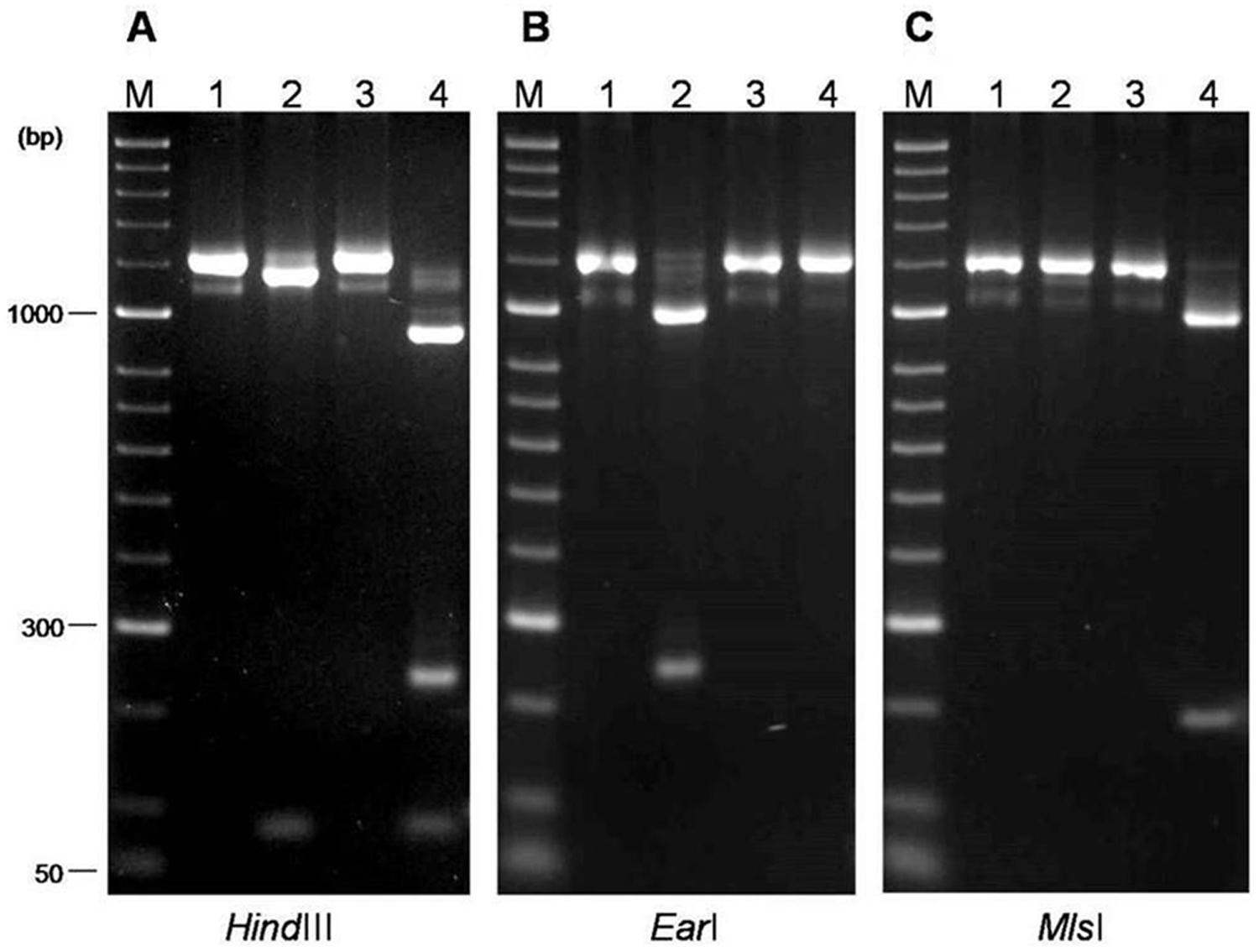
Polymerase chain reaction–restriction fragment length polymorphism patterns of 16S ribosomal DNA fragment (1,235 bp) using different restriction endonucleases (
Amplicons of 8 of the 16 P. multocida strains (marked with asterisk in Table 1) representing the 2 RFLP groups (4 from each one) were sequenced. The PCR fragments were purified using a commercial kit h for sequencing. Dye-labeled products were run on a sequencer i at the DNA laboratory of the Agricultural Biotechnology Center. j The obtained 16S rRNA partial gene sequences were aligned with the GenBank database using the MegAlign program in a commercial software package, k with the ClustalW multiple alignment algorithm. The lineage-specific nucleotide substitutions could be identified in the strains examined. Existence of unique recognition sites were found on G – T substitution at 215 position (MlsI TGG↓CCA) and AG(C) – CT(T) substitutions at 1009-1011 positions (EarI ↓(N)4GAAGAG and HindIII A↓AGCTT, respectively; Table 2). The strains from the 2 different RFLP groups (I and II) corresponded with phylogenetic lineage A and B of 16S rDNA, respectively.
For evaluating the relationship between lineages and traditional subspecies, the phenotypic characteristics of the strains were examined in classical biochemical tests (fermentation of dulcitol, D(-)sorbitol, trehalose, and detection of α-glucosidase activity; Table 3).11,13 The presumption is that P. multocida subsp. gallicida strains are dulcitol and sorbitol positive but trehalose negative; subsp. multocida strains are dulcitol negative but sorbitol and trehalose positive; and subsp. septica strains are dulcitol and sorbitol negative but trehalose positive. The α-glucosidase activity characterizes dominantly strains of subsp. septica. 7 However, the results were confused and confirmed the disagreement between phenotypic subspecies identification and molecular genetic analysis, as described earlier by other authors.4,9,14
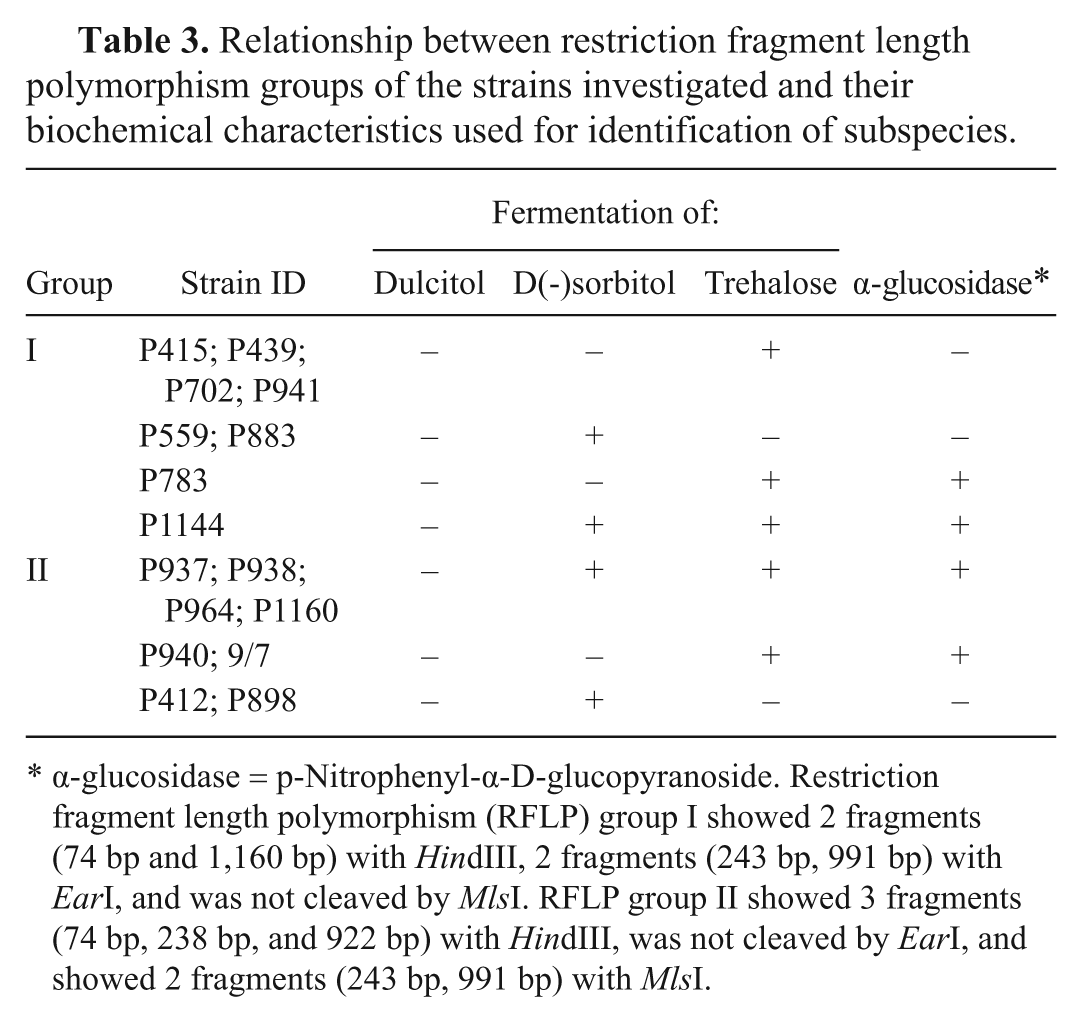
α-glucosidase = p-Nitrophenyl-α-D-glucopyranoside. Restriction fragment length polymorphism (RFLP) group I showed 2 fragments (74 bp and 1,160 bp) with HindIII, 2 fragments (243 bp, 991 bp) with EarI, and was not cleaved by MlsI. RFLP group II showed 3 fragments (74 bp, 238 bp, and 922 bp) with HindIII, was not cleaved by EarI, and showed 2 fragments (243 bp, 991 bp) with MlsI.
The remarkable phenotypic diversity makes recognition of subgroups formed by genetically related P. multocida strains difficult although their identification might be useful in epidemiological studies. 8 Sequence analysis of housekeeping genes and the 16S rRNA gene can distinguish 2 main lineages (A and B) within the species providing a powerful but relatively expensive method for the differentiation of strains. Based on nucleotide substitutions in the 16S rRNA gene recognized by digestion with HindIII, EarI, and MlsI restriction endonucleases, a practical and easily performable PCR-RFLP method was established that facilitates the simple tracking of the distribution of P. multocida phylogenetic lineages in different hosts that may help in better understanding their role in various diseases.
Footnotes
a.
Difco, BD, Sparks, MD.
b.
Nebotrade, Biatorbágy, Hungary.
c.
Bio-Rad Laboratories Inc., Hercules, CA.
d.
Sigma-GenoSys, Steinheim, Germany.
e.
DreamTaq, Fermentas Inc., Burlington, Ontario, Canada.
f.
Biometra T1, Biometra GmbH, Gottingen, Germany.
g.
HyperLadder II, Bioline Reagents Ltd, London, UK.
h.
PureLink Quick Gel Extraction Kit, Invitrogen Corp., Carlsbad, CA.
i.
ABI Prism 3100 sequencer, Applied Biosystems, Foster City, CA.
j.
Biomi Ltd, Gödöllő, Hungary.
k.
Lasergene, DNASTAR Inc., Madison, WI.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the Hungarian Scientific Research Fund (OTKA, project no. PD101091).