
Abstract
A new highly sensitive and specific hemi-nested reverse transcription polymerase chain reaction (RT-PCR) assay was applied to detect nucleoprotein (NP) gene of Canine distemper virus (CDV) in samples collected from dogs showing respiratory, gastrointestinal, and neurological signs. Thirty-eight out of 86 samples were positive suggesting that despite the vaccination, canine distemper may still represent a high risk to the canine population. The 968 base pair (bp) fragments from the hemagglutinin (H) gene of 10 viral strains detected in positive samples were amplified and analyzed by restriction fragment length polymorphism (RFLP) using AluI and PsiI enzymes in order to differentiate among vaccine and wild-type CDV strains and to characterize the field viral strains. The products of the both enzymatic digestions allowed identification all viruses as wild strains of CDV. In addition, the RFLP analysis with AluI provided additional information about the identity level among the strains analyzed on the basis of the positions of the cleavage site in the nucleotide sequences of the H gene. The method could be a more useful and simpler method for molecular studies of CDV strains.
Keywords
Introduction
Canine distemper is a highly contagious viral infection caused by Canine distemper virus (CDV; order Mononegavirales, family Paramyxoviridae, subfamily Paramyxovirinae, genus Morbillivirus). Gastrointestinal, respiratory, and neurological signs characterize the disease, and it has long been considered one of the leading causes of death for dogs worldwide. In the 1950s, the introduction of live vaccines against CDV has allowed for the control of infection, 8 though despite vaccinations, outbreaks of canine distemper seem to be increasing throughout the world.2,3,5,7,15,23,24 The development of more sensitive and specific diagnostic techniques based on reverse transcription polymerase chain reaction (RT-PCR) amplification, compared to traditional assays such as indirect fluorescent antibody test and enzyme-linked immunosorbent assay, has helped to identify a higher number of infected animals than ever before.2,4,10,12,20,21
Although different viral gene sequences have been identified for detection of CDV in biological specimens and tissues, the highly conserved nucleoprotein (NP) gene appears a better target for amplification of specific fragments from all CDV strains.2-4,10,11,19,20,27 However, the sensitivity of molecular test varies depending on samples collected, RNA extraction protocols, or primers used. In particular, RT-PCR combined with a nested or hemi-nested PCR seems a more sensitive method for the diagnosis of canine distemper.10,12,19,22,25 Furthermore, molecular biology allows the comparison of genetic sequences of CDV strains identified in the field, highlighting that the hemagglutinin (H) genes undergo higher genetic/antigenic variations. 17
Considering the variability in the H gene, the field strains of CDV cluster into 6 major genetic lineages designated as America-1 (including several vaccine strains), America-2, Asia-1, Asia-2, European, and Arctic, according to their geographic spread.9,15,17 However, some viral strains used as vaccines appear distinct from the America-1 lineage, 14 and additional geographic patterns have been proposed based on the H gene analysis of wild-type CDV strains circulating in Asia, Africa, Argentina, and Mexico.2,6,28,29
The impact of genetic heterogeneity remains unclear, even if the sequence variations in viral genome could modify virulence, tropism, or antigenicity features of wild CDV strains with respect to that present in the vaccine, increasing the risk of infection in dog populations. Therefore, the H gene can be considered the most appropriate molecular target to identify genetic/antigenic changes of the virus and the relationships among different strains of CDV as previously reported.1-3,9,15,23,26-29 In addition, H gene analysis appears to be a fast and suitable method to differentiate wild-type viruses from the vaccine strains of CDV when the clinical manifestations of the infection occur in recently vaccinated animals. For this purpose, several protocols of RT-PCR and restriction fragment length polymorphism (RFLP) analysis targeting the H gene sequence have been developed.3,16,23,26,27 The aim of the current study was to apply a new hemi-nested RT-PCR for the laboratory diagnosis of distemper in dogs suspected of CDV infection and to analyze the H gene of the viral strains by means of RT-PCR RFLP assay.
Materials and methods
Animals and clinical samples
Eighty-six samples were collected from 53 dogs with history of gastrointestinal, respiratory, and neurological signs. The samples were taken over the years 2005–2008 at various small animal practices, veterinary hospitals, and dog kennels in central Italy. In particular, 24 dogs (45.3%) belonged to private owners, while 29 animals (54.7%) were stray dogs housed in 6 different kennels. Table 1 details the age and vaccination of dogs. Samples consisted of conjunctival, nasal, genital, and rectal swabs (n = 41), buffy coats (n = 24), blood sera (n = 8), cerebrospinal fluid (CSF; n = 5), urine (n = 1), and various organs (n = 7; cerebellum, intestine, spleen, and lung), with the latter collected from animals that succumbed following suspected CDV infection.
Detection by hemi-nested reverse transcription polymerase chain reaction (RT-PCR) and genetic characterization of wild-type strains of Canine distemper virus (CDV) in suspected infected dogs: vaccination status and age group of the dogs with gastrointestinal, respiratory, and neurological signs examined by hemi-nested RT-PCR to detect the nucleoprotein gene of CDV.*
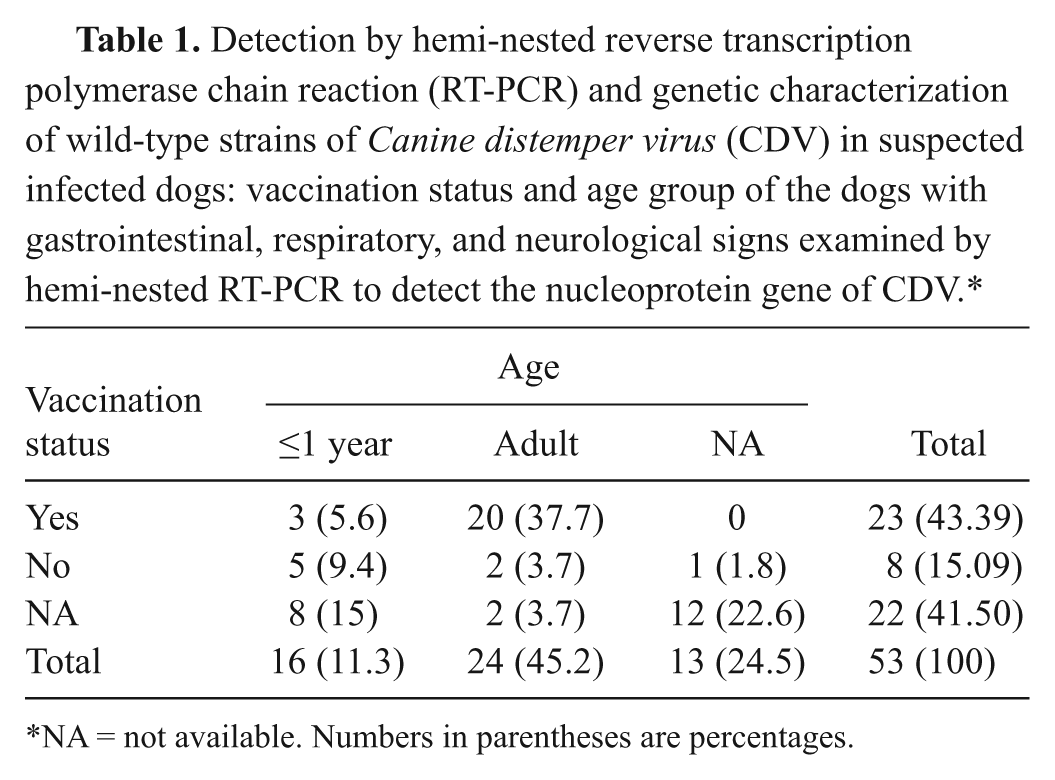
NA = not available. Numbers in parentheses are percentages.
Hemi-nested reverse transcription polymerase chain reaction
Tissue samples were suspended (1 g) in 10 ml of ice-cold phosphate buffered saline (pH 7.2), homogenized using a rotor-stator homogenizer, and centrifuged at 328 × g for 10 min at 4°C to collect the supernatant. The swabs were immersed in ice-cold phosphate buffered saline (pH 7.2) while the urine was centrifuged at 328 × g for 10 min at 4°C to collect the epithelial cells. The viral RNA was isolated from all specimens using a commercial kit, a according to the manufacturer’s instructions.
A primer pair, C2 and REV1441, specific to a 350–base pair (bp) segment within the conservative part of the NP gene of the CDV genome was designed for the first RT-PCR amplification. The reaction was performed in a total mixture of 25 μl containing 12.5 μl of 2× master mix, b 0.5 µl of reverse transcriptase, c 0.5 μl of each primer (100 pmol/μl), and 2 µl of RNA. Reverse transcription was carried out at 45°C for 60 min followed by an amplification protocol consisting of 35 cycles of denaturation at 94°C for 30 sec, primer annealing at 48°C for 45 sec, extension at 72°C for 45 sec, and 5 min of final extension at 72°C. The reaction mixture without the matrix was used as negative control whereas the RNA from the CDV vaccine strain Onderstepoort propagated in Vero (African green monkey kidney epithelial) cells as positive control. Following the RT-PCR, an agarose gel electrophoresis and ultraviolet transilluminator visualized the amplicons.
The second amplification was carried out by the primers FW1261 and REV1441, specific to the internal fragment of 180 bp of the first amplicon, and 1 μl of negative RT-PCR templates diluted 1:10. The reaction mixture was incubated at 94°C for 2 min, and amplification was obtained by 25 cycles of denaturation at 94°C for 30 sec, primer annealing at 48°C for 45 sec, extension at 72°C for 45 sec, and 5 min of final extension at 72°C. The reaction products were analyzed by agarose gel electrophoresis and ultraviolet light visualization. The nucleotide positions and sequences of the primers used in hemi-nested RT-PCR protocols are shown in Table 2.
Detection by hemi-nested reverse transcription polymerase chain reaction (RT-PCR) and genetic characterization of wild-type strains of Canine distemper virus (CDV) in suspected infected dogs: sequences and nucleotide position of the primers utilized in hemi-nested RT-PCR to detect the nucleoprotein gene of CDV in suspected infected dogs and in restriction fragment length polymorphism (RFLP) and sequence analysis of the hemagglutinin gene for genetic characterization of wild-type CDV strains.

See reference 16.
Specificity and sensitivity of hemi-nested reverse transcription polymerase chain reaction
The specificity of the hemi-nested RT-PCR was evaluated by using the CDV strain Onderstepoort, Canine parvovirus type 2 (CPV-2), Canine adenovirus (CAV), Alphacoronavirus 1 (ACoV-1), and mock-infected cells. In order to evaluate the sensitivity of hemi-nested RT-PCR, the 350-bp RT-PCR amplicon was purified from agarose gel by using a commercial kit d and cloned into pCR2.1 vector. e Plasmid DNA was purified using a commercial kit f and quantified by measuring the optical density at 260 nm. Serial 10-fold dilutions of plasmid DNA were amplified in hemi-nested PCR using FW1261 and REV1441 primers.
Restriction fragment length polymorphism analysis of H gene
The RFLP analysis was performed on the 968 DNA fragment from the 10 CDV strains H gene detected by hemi-nested RT-PCR and the Onderstepoort strain the latter used as positive control. The CDV strains selected were collected from animals with different epidemiological features (origin, age, vaccination, and clinical signs; Table 3). The amplicons were produced in a total reaction mixture of 50 µl containing 0.5 μl of the primers 204 16 and CDVHint (Table 2), 25 µl of reverse transcriptase and Taq polymerase reaction mix, g and 3 µl of each RNA. The amplification was performed by 60 min of reverse transcription at 48°C, 40 cycles of denaturation at 94°C for 30 sec, primers annealing at 50°C for 45 sec, extension at 68°C for 1.5 min, and 10 min of final extension at 68°C. Aliquots (2 μl) of the RT-PCR products were digested with PsiI and AluI enzymes h overnight at 37°C. The resulting fragments were detected by electrophoresis in 2% agarose gels and ethidium bromide staining.
Detection by hemi-nested reverse transcription polymerase chain reaction (RT-PCR) and genetic characterization of wild-type strains of Canine distemper virus (CDV) in suspected infected dogs: epidemiological data collected from CDV-positive dogs selected for restriction fragment length polymorphism analysis of the hemagglutinin gene of CDV strains detected by diagnostic hemi-nested RT-PCR.*
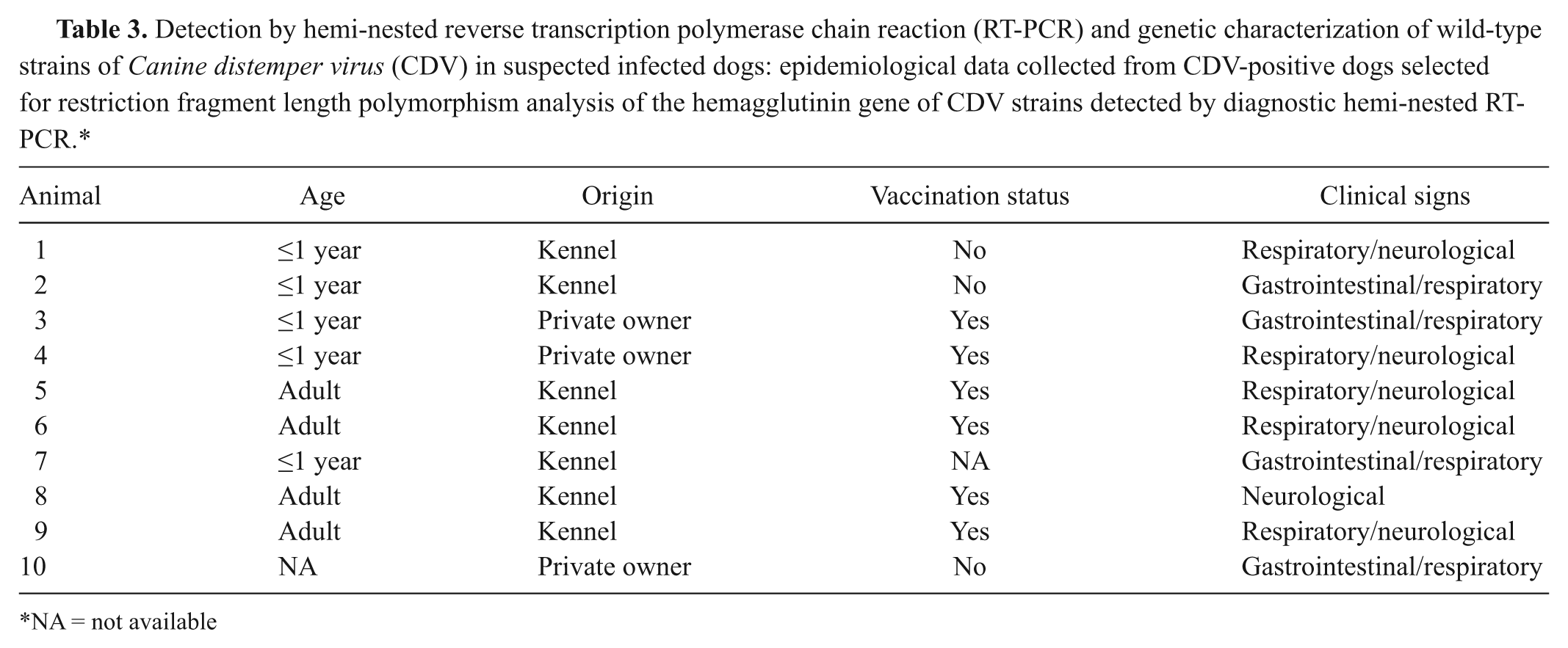
NA = not available
The PsiI enzyme is able to cut the vaccine strain amplicons but not the wild-type strains. 3 The AluI enzyme was used in order to verify its ability to distinguish between vaccine and wild-type CDV strains and to identify a genetic heterogeneity of the CDV strains on the basis of the different positions of the cleavage site in the nucleotide sequences of the H gene.
Sequence and phylogenetic analysis
The amplicons were purified with a commercial gel extraction kit, d sequenced using the primers 204 and CDVHint, and nucleic acid sequences were identified using the Basic Local Alignment Search Tool (BLAST, http://blast.ncbi.nlm.nih.gov/Blast.cgi) and aligned with CDV H-gene sequences by using ClustalW2 (http://www.ebi.ac.uk/Tools/msa/clustalw2/) and FASTA (http://www.ebi.ac.uk/Tools/sss/fasta/) programs. Phylogenetic analysis (neighbor-joining) with bootstrap statistical support (1,000 replicates) was conducted using the MEGA software package, version 3.1. 13 The sequences of the H gene fragments of 3 CDV strains detected in the current study are available in GenBank database under accession numbers GQ328902.1, GQ328903.1, and GQ328904.1.
Results
Hemi-nested reverse transcription polymerase chain reaction
The hemi-nested RT-PCR revealed CDV sequences in 38 samples (Fig. 1). Of these, 19 specimens were positive by RT-PCR (12 mucosal swabs, 5 buffy coats, and 2 blood sera) while the second amplification by hemi-nested PCR was obtained in an additional 19 samples (50%; 13 mucosal swabs, 3 samples of spleen, 1 buffy coat, 1 serum, and 1 urinary sediment; Fig. 2). The positive samples were collected from 30 out of 53 dogs (56.6%) that were periodically vaccinated (60%), >1 year old (63.3%), and came from dog kennels (63.3%).
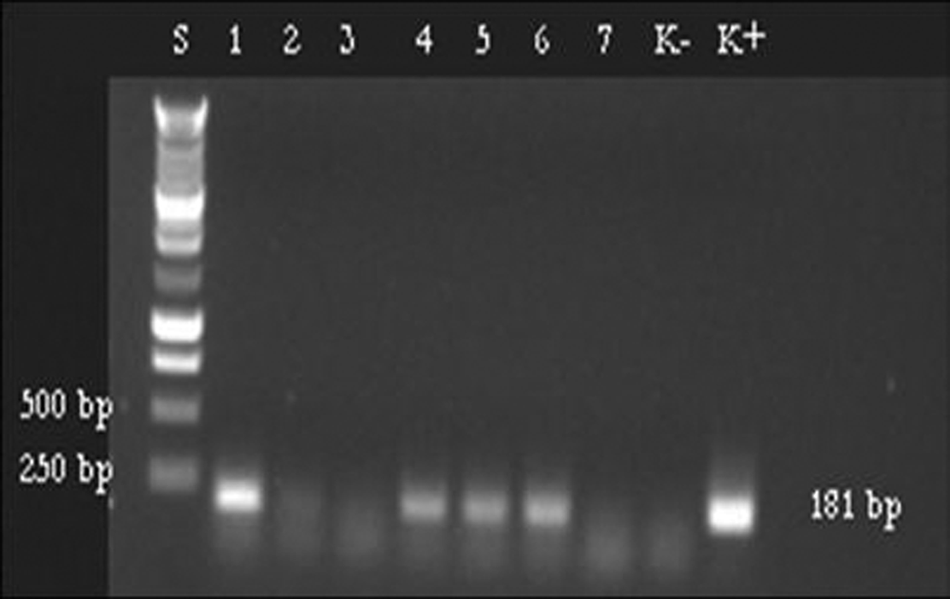
Hemi-nested reverse transcription polymerase chain reaction of 8 samples collected in the study: a 181-bp fragment of the nucleoprotein gene of Canine distemper virus (CDV) was amplified by using C2, Fw1261, and REV1441 primers from 4 samples (lines 1, 4–6). Lane S: 1-kb DNA ladder; lanes 1, 4–6: positive samples; lanes 2, 3, 7: negative samples; lane K–: negative control; lane K+: CDV strain Onderstepoort.
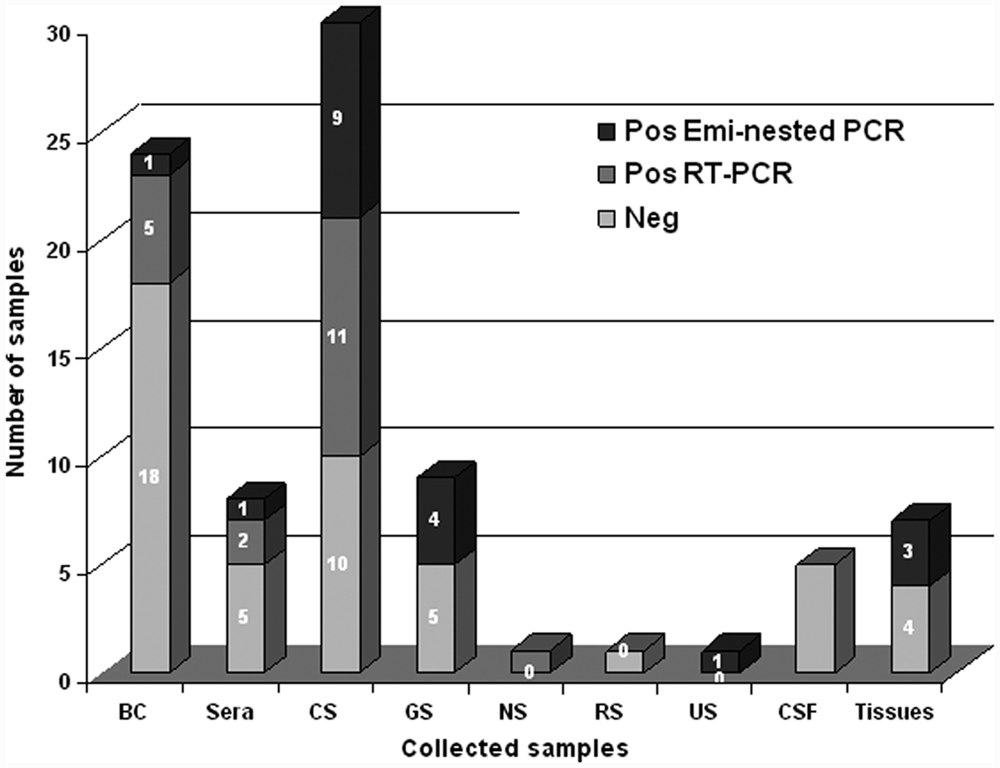
Hemi-nested reverse transcription polymerase chain reaction (RT-PCR): results of the 86 samples collected from dogs with suspected infection of Canine distemper virus. BC = buffy coat; CS = conjunctival swabs; GS = genital swabs; NS = nasal swabs; RS = rectal swabs; US = urinary sediment; CSF = cerebrospinal fluid.
Specificity and sensitivity of hemi-nested reverse transcription polymerase chain reaction
The expected 350- and 180-bp fragments of NP gene were obtained in CDV strain Onderstepoort–infected cells but not in CPV-2–, CAV–, and ACoV-1–infected cells. Similarly, no detectable bands were produced from mock-infected cells (data not shown), while the lower detection limit of the 180-bp fragment was 0.11 pg/ml corresponding to 23.2 copies of cloned DNA (data not shown).
Restriction fragment length polymorphism analysis of H gene
The PCR products for RFLP analysis were obtained from all selected CDV strains, as well as from the Onderstepoort strain used as the positive control (Fig. 3). Following PsiI digestion, 2 bands of 295 bp and 673 bp were obtained from a CDV vaccine strain only. The PsiI enzyme did not cut the PCR products obtained from field strains (Fig. 4). The RFLP using the AluI enzyme produced 4 fragments (570 bp, 214 bp, 97 bp, and 87 bp) from the vaccine strain; while 3 bands (2 fragments included between 500 bp and 250 bp, and 1 less than 250 bp molecular weights) were visualized from all field strains of CDV (Fig. 5).
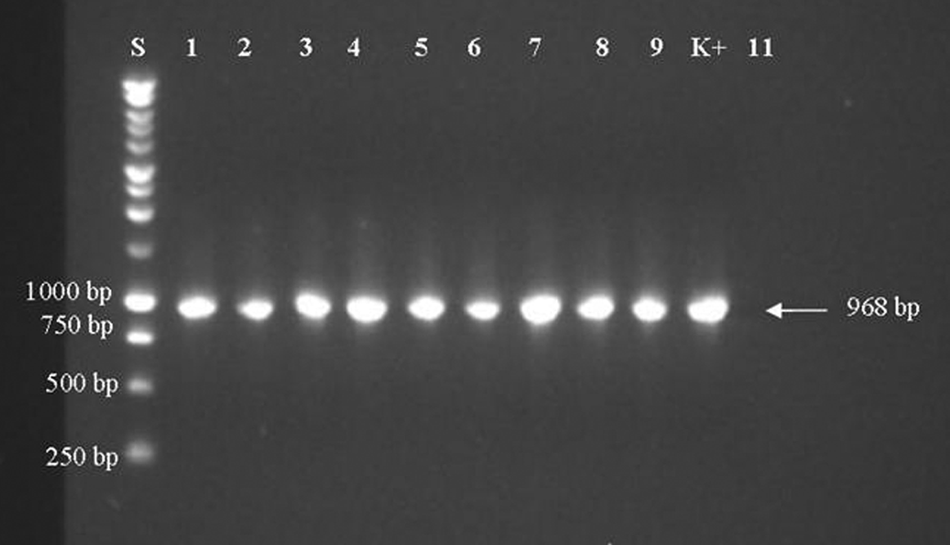
Reverse transcription polymerase chain reaction amplicons of the hemagglutinin gene: a 968-bp fragment was amplified by using 204 and CDVHint primers. Lane S: 1-kb DNA ladder; lanes 1–9: positive samples; lane K+: Canine distemper virus strain Onderstepoort; lane 11: negative control.
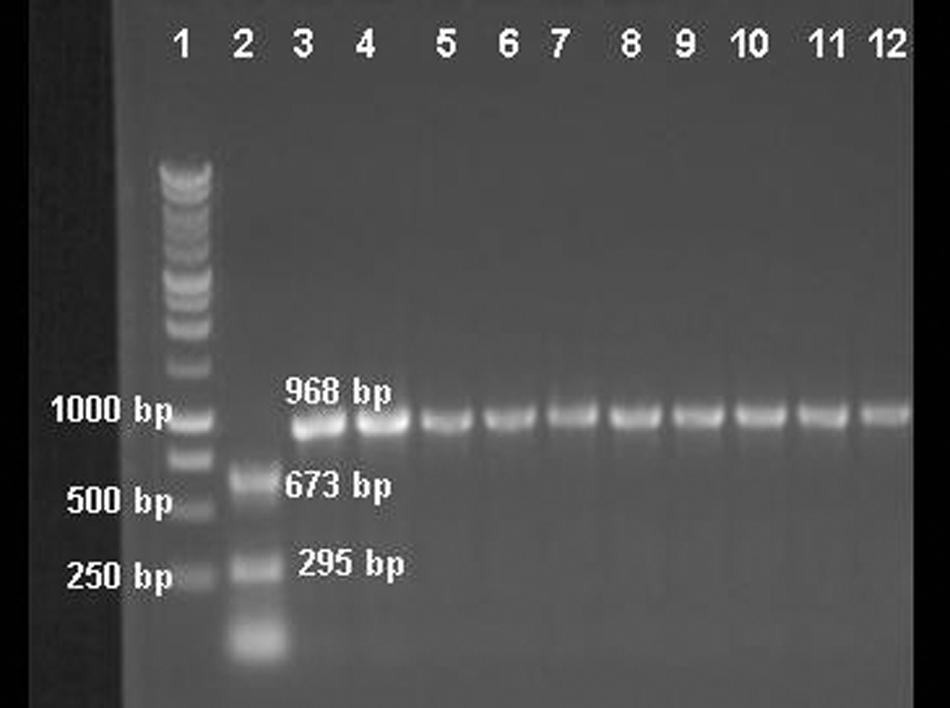
Restriction fragment length polymorphism analysis: the 968-bp fragment of the hemagglutinin gene of Canine distemper virus (CDV) strain Onderstepoort was digested by PsiI enzyme, while the field strains were not cut by the enzyme. Lane 1: 1-kb DNA ladder; lane 2: CDV strain Onderstepoort digested by PsiI enzyme; lanes 3–12: field strains of CDV that were not cut by the enzyme.
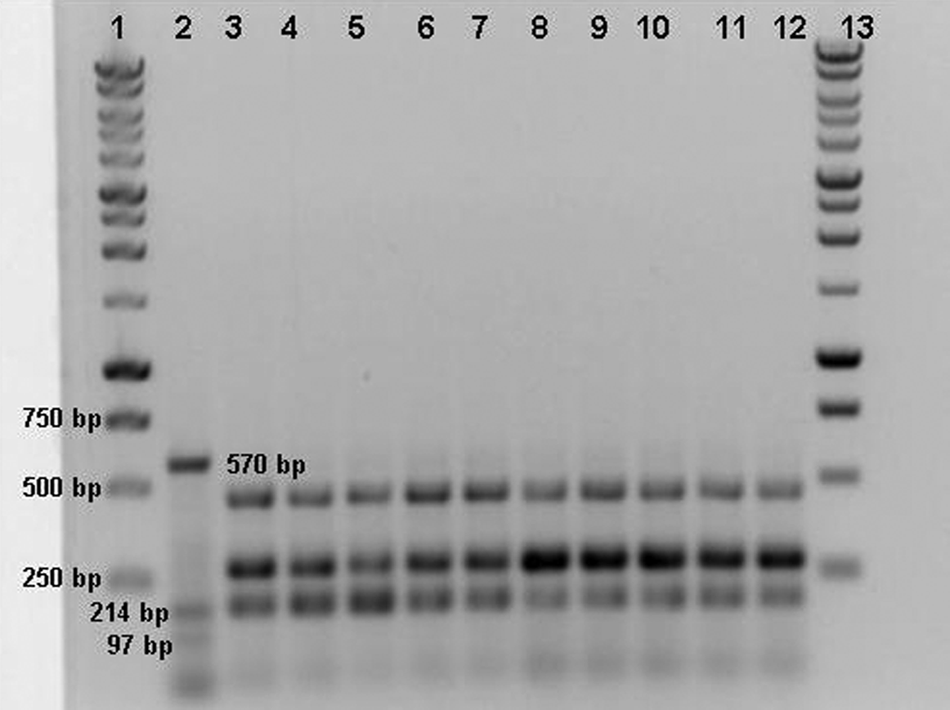
Restriction fragment length polymorphism analysis: the 968-bp fragment of the hemagglutinin gene of Canine distemper virus (CDV) strain Onderstepoort was cut by AluI enzyme in 4 fragments (570 bp, 214 bp, 97 bp, and 87 bp); all field strains were digested producing 3 bands with a molecular weight that ranged from 150 bp to 500 bp. Lanes 1, 13: 1-kb DNA ladder; lane 2: CDV strain Onderstepoort; lanes 3–12: field strains of CDV.
Sequence and phylogenetic analysis
All of the 968-bp fragments were specific for the H gene of CDV. The sequence alignments demonstrated that the nucleotide identity level among the strains varied from 97% to 99%. In comparison with other sequences available in GenBank, the highest identity (98.7%) was found to 111/03B (GenBank accession no. DQ494318) and 265/02 (GenBank DQ494319) CDV strains, both detected in southern Italy and included in the European lineage. 15 The nucleotide identity decreased to 89% compared to the Onderstepoort strain. The phylogenetic tree based on the nucleotide sequences of the H-gene fragment (Fig. 6) confirmed that the CDV strains detected in the current study segregated with the DQ494318 and DQ494319 Italian strains along with a CDV strain recently detected in Hungary. 3 The sequence analysis showed that all the amplicons contained the cleavage site of AluI at the nucleotide positions 154 and 461 in accordance with the RFLP profile obtained after enzymatic digestion. Therefore, the fragments visualized after the AluI digestion of all field strains of CDV resulted in 184 bp, 306 bp, and 448 bp long, and the same RFLP pattern was observed comparing the sequences of related 111/03B and 265/02 strains (Fig. 7).
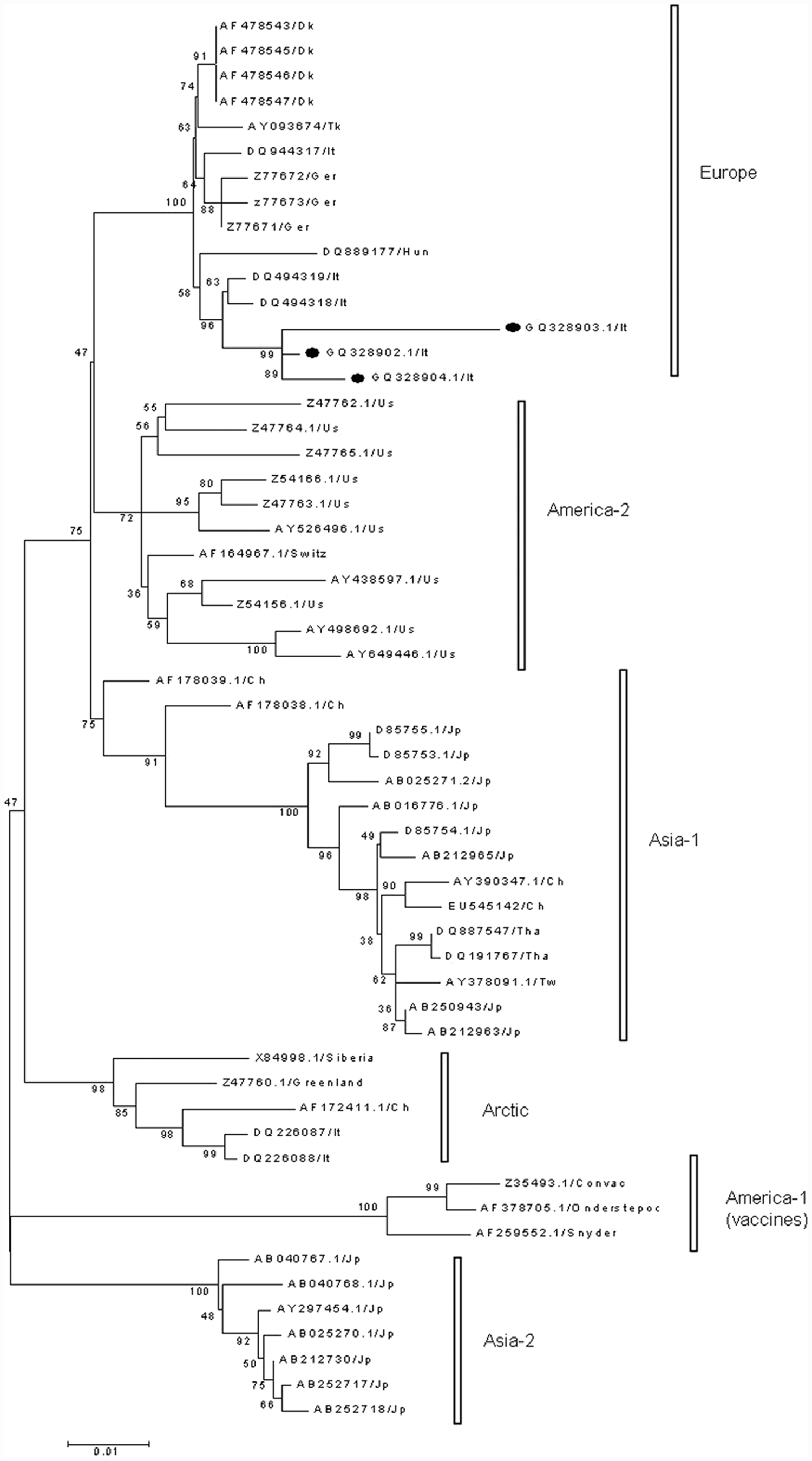
Phylogenetic tree constructed on the 968 base pair (bp) nucleotide sequence of the H protein of Canine distemper virus strains: the tree was generated by the neighbor-joining method with bootstrap analysis (1,000 replicates) and p-distance model. The published sequences GQ328902.1, GQ328903.1, and GQ328904 are highlighted with black circles. Ch = China; Dk = Denmark; Ger = Germany; Hun = Hungary; It = Italy; Jp = Japan; Switz = Switzerland; Tw = Taiwan; Tha = Thailand; Tk = Turkey; US = United States.

Partial alignment by using ClustalW2 of the hemagglutinin gene fragment of field strains of Canine distemper virus analyzed by restriction fragment length polymorphism in comparison with related 111/03B and 265/02 viruses. The positions of the cleavage site of AluI enzyme is marked by bold font.
Discussion
The results obtained herein show that the hemi-nested RT-PCR carried out on clinical specimens can be considered a very sensitive test to confirm CDV infection in dogs with suspected symptoms. Indeed, the second step of PCR allowed for the doubling of the number of positive samples as compared with RT-PCR, detecting the virus even in specimens containing low or partially degraded RNA. In biological samples, endogenous and exogenous RNase enzymes can degrade RNA due to incorrect handling or storing of samples at elevated temperature, potentially compromising the efficiency of the molecular test. Furthermore, the amplification of small size fragments of the viral genome could be useful to improve the laboratory diagnosis of canine distemper from clinical specimens with a low viral load. 18
Several samples, including mucosal swabs, buffy coat, blood serum, urine, CSF, and tissues (including lung, brain, and others) have been used for the diagnosis of canine distemper. Conjunctival swabs have provided a higher detection rate (20/30; 66%) than the other specimens in accordance with a recent study 11 in which CDV was detected in conjunctival epithelium before viremia and for longer periods. In addition, conjunctival swabs are the most suitable specimens for the diagnosis of distemper because they are noninvasive and easy to collect along with genital swabs.
More than 50% of dogs with suspected infection resulted positive, and the epidemiological data collected showed that the majority of infected dogs were >1 year old and underwent regular vaccination against CDV suggesting that CDV continues to be a serious threat to the general dog population despite vaccination. Improper storage or exposure to extreme temperatures may have compromised effective immunization of dogs. In addition, vaccine failure could have occurred by applying incorrect schedules, which did not provide protective immunization. 6 The origin of the dogs might have been an additional risk factor capable of increasing the diffusion of CDV infection in already immunocompromised animals. No biosecurity measures, such as quarantine of newly acquired dogs, isolation of ill dogs, and strong cleaning and disinfecting protocols, had been applied in kennels from which 50% of infected dogs originated.
The RFLP analysis performed on the 10 CDV field strains showed that these viruses are genetically different from the Onderstepoort strain. Therefore, it can be excluded that the hemi-nested RT-PCR had given positive results due to recent vaccination. The results obtained suggest that both enzymes, PsiI and AluI, may be used to confirm the etiological diagnosis of distemper especially when the symptoms related to the CDV infection occur after vaccination. Moreover, RFLP using AluI provided additional information about the identity level among the strains analyzed in accordance with the sequencing analysis that identified all field strains belonging to European lineage with the nucleotide identity level of 97–99%. Considering the small number of viruses tested, further analyses are needed to establish if RFLP analysis by AluI enzyme is a valid method to characterize CDV field strains as well as any new circulating viruses belonging to other genetic lineages. In conclusion, canine distemper is a disease of dog populations that should be included in the list of differentials for clinical diagnosis of any multisystemic condition characterized by fever, respiratory, enteric, and neurological signs. Laboratory tests are essential to confirm the infection, and molecular assays are useful for monitoring and differentiating the CDV field strains.
Footnotes
Notes
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) received no financial support for the research, authorship, and/or publication of this article.