
Abstract
Serological testing for toxoplasmosis diagnosis remains the method of choice in human medicine due to the accessibility of the requisite sample, the difficulty in predicting the parasite’s location in the host for direct detection, and the availability of established commercial methods. In veterinary medicine, although the first 2 conditions are unchanged, there is a need for commercially produced test methods that are validated for Toxoplasma gondii detection across the range of animal species that can serve as intermediate hosts. The development of such a serological method for animals would allow the diagnosis of toxoplasmosis in individual animals and a higher throughput method for population-level toxoplasmosis surveys. The incorporation of a non–species-specific chimeric protein A/G conjugate into an anti–Toxoplasma immunoglobulin G enzyme-linked immunosorbent assay is described. Serum from potential intermediate hosts was reevaluated using this method and compared with earlier testing using an established agglutination procedure. Very good agreement between the 2 tests was noted (κ = 0.81), establishing the method as a useful option for veterinary diagnostic testing.
Toxoplasmosis, caused by Toxoplasma gondii, is a disease that has health and economic impacts across a broad spectrum of animal species.8,14,15 Diagnosis of toxoplasmosis is complicated both by the nonspecific nature of clinical signs and the complexity of the pathogen’s life cycle, making direct detection unfeasible in most situations.5-7,9,11,20 A range of serological options have been developed to detect exposure to Toxoplasma in human beings, some of which have utility in veterinary diagnostics.17,18 Various permutations of agglutination testing can be applied successfully to testing a wide range of animal species, as they have no species-specific conjugate requirement.3,4,13 However, there are no commercially produced agglutination tests currently available in the United States, whereas many other serological tests are either available or being developed for human testing. 18 Unfortunately, many of these tests require protein conjugate for the detection of anti-Toxoplasma antibody in the serum of interest. Most of the protein conjugates available are only effective in binding to antibody from specific animal species. The description of the use of protein conjugates that bind to the immunoglobulin (Ig)G of many different animal species is a promising new platform for serological test development.19,22,25
The use of non–species-specific protein conjugates A and G allows the testing of sera from most of the veterinary species of interest on an enzyme-linked immunosorbent assay (ELISA) platform. However, this requires 2 parallel tests to be run with 2 sets of control, one for samples to be tested with protein A conjugate and one for those to be tested with protein G. The development of chimeric protein A/G conjugate from staphylococcal protein A and streptococcal protein G has been described. 10 This chimeric protein A/G conjugate allows the testing of all species formerly tested by the 2 proteins individually with a single conjugate, thereby simplifying the procedure and making it more attractive as a potential substitute for currently used agglutination techniques.1,25 A single test allows higher throughput and is better suited to commercial development and automation. In the current study, the results of a commercially produced indirect hemagglutination assay were compared with those of a commercially available IgG ELISA, modified to detect IgG antibody in multiple animal species by the substitution of protein A/G horseradish peroxidase conjugate for the kit-provided antihuman conjugate.
Blood samples were collected in clot tubes from animals by referring veterinarians. The extracted serum samples were submitted to the Animal Health Diagnostic Center at Cornell University (Ithaca, New York) from 2002 to 2008 for detection of Toxoplasma exposure by indirect hemagglutination. After storage at 5°C, positive samples with a sufficient amount of serum remaining and a sampling of negative samples were reassessed using the modified IgG ELISA technique in 2011. The sera tested included samples from 14 alpacas, 17 sheep, 17 goats, 15 horses, and 56 dogs.
Seroreactivity to Toxoplasma antigens was initially evaluated using an indirect hemagglutination assay (IHA). a The commercial preparation of the IHA had perfect qualitative agreement with the Centers for Disease Control and Prevention Toxoplasma indirect fluorescent antibody test in their evaluation of human serum samples. 24 Serum containing anti-Toxoplasma antibody causes agglutination of the sheep red blood cells coated with antigen as part of the test procedure.3,13 Nonspecific agglutination is detected by assaying coated and noncoated red blood cells in separate wells. Interpretation is based on observation of presence or absence of a red blood cell mat covering the bottom of the test well on completion of incubation. The test is equally effective for human and animal diagnostic work because no species-specific conjugate is required for antibody detection. 23 The procedure was used as a reference test to evaluate the performance of the modified IgG ELISA, with details described in an earlier study. 19
A commercial IgG ELISA b was modified by use of protein A/G horseradish peroxidase conjugate c to replace the conjugated goat antihuman IgG included in the kit. Samples, controls, and a calibrator were evaluated after dilution of 1:21 with kit-supplied diluent (containing Tween-20, bovine serum albumin, and phosphate buffered saline). These components were added to kit wells that had been sensitized by passive absorption with Toxoplasma antigen. Any antigen-specific antibody contained in the sample binds to the immobilized antigen. Protein A/G conjugate was added to each well to attach to any immobilized IgG antibody. Following washing, microwells were incubated with tetramethylbenzidine, a peroxidase substrate solution that changes color when hydrolyzed by the presence of peroxidase. Color intensity of the sample indicated antibody concentration in the microwell. This was evaluated using a microplate reader with light at a wavelength of 450 nm. Comparison to kit-provided calibrators guided the identification of positive versus negative serum, with a ratio of sample color intensity to kit calibrator color intensity of greater than or equal to 1.1 indicating a positive reaction for anti–Toxoplasma IgG antibody as described by the kit insert. A ratio between 0.9 and 1.1 (noninclusive) was considered an indeterminate result, and less than or equal to 0.9 was considered a negative result.
Protein A/G chimeric horseradish peroxidase conjugate was used in the IgG ELISA assays instead of the kit conjugate to allow testing of multiple animal species. This protein is produced by gene fusion of the Fc-binding domains of protein A and protein G, bacterial cell wall proteins with affinity for IgG in a range of species. It contains 4 Fc-binding domains from protein A and 2 from protein G, providing binding for all antibody species and subclasses recognized by either protein A or protein G. 10 Protein A conjugate is reported to provide strong binding to IgG antibody of mice, rabbits, cats, dogs, and pigs, whereas protein G conjugate is reported to provide strong binding to IgG antibody of mice, guinea pigs, rabbits, cows, sheep, goats, alpacas, horses, and pigs.2,19,21 Protein was received as lyophilized powder and was diluted 1:40,000 with phosphate buffered saline (pH 7.2) prior to use. The dilution was determined by testing against kit controls to satisfy quality control parameters.
The significance of association between the results of the IHA and the ELISA was evaluated using the McNemar chi-square test. 16 The degree of agreement between the results from the 2 tests was quantified using kappa statistics. The accuracy of the ELISA test in detecting exposure to Toxoplasma was evaluated in comparison to the IHA and measured using the relative sensitivity and specificity. The expected performance of the ELISA at different cutoff points was examined using the receiver operating characteristic (ROC) curve. 12
Serum samples were evaluated from multiple animal species using both the IHA and the IgG ELISA modified as described. The sera tested included samples from some of the more commonly tested intermediate hosts for the parasite. There was no significant difference detected between positive versus negative results when comparing IHA and ELISA results with McNemar chi-square (P = 1.0), with a kappa value of agreement of 0.81 (95% confidence interval [CI]: 0.71–0.92 calculated using GraphPad d ), as noted in Table 1. This reaffirms the finding of an earlier study, using protein A and G conjugates individually, of very good agreement between the agglutination and ELISA testing. The breakdown of these results by animal species is provided in Table 2, with all species, except horses, demonstrating very good to perfect agreement between the 2 test methods. The relative sensitivity of this ELISA using chimeric protein A/G conjugate was found to be 92% (95% CI: 82–97%) and relative specificity 89% (95% CI: 77–96% calculated using VassarStats e ) when the IHA results were used as a gold standard and when positive versus negative ELISA results were determined using the manufacturer’s guidelines, b including 0.9–1.1 as the equivocal range for optical density ratios. The variation of relative sensitivity and specificity at different optical density ratios is shown in Figure 1. The associated ROC curve is depicted in Figure 2.
Comparison of indirect hemagglutination assay (IHA) to the modified immunoglobulin G enzyme-linked immunosorbent assay (ELISA).

Results of modified immunoglobulin G enzyme-linked immunosorbent assay testing by species.
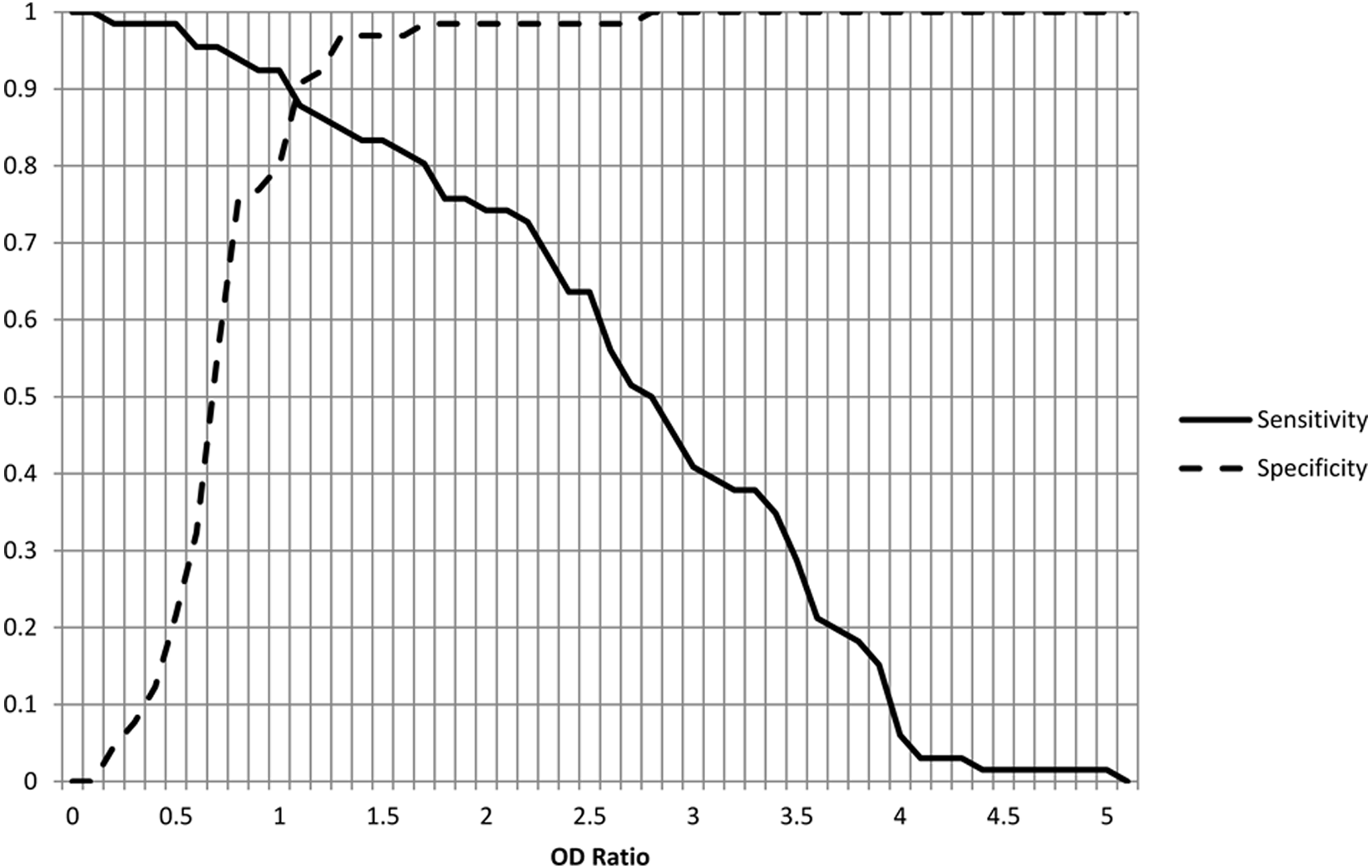
Variation in sensitivity and specificity relative to enzyme-linked immunosorbent assay optical density (OD) ratios.
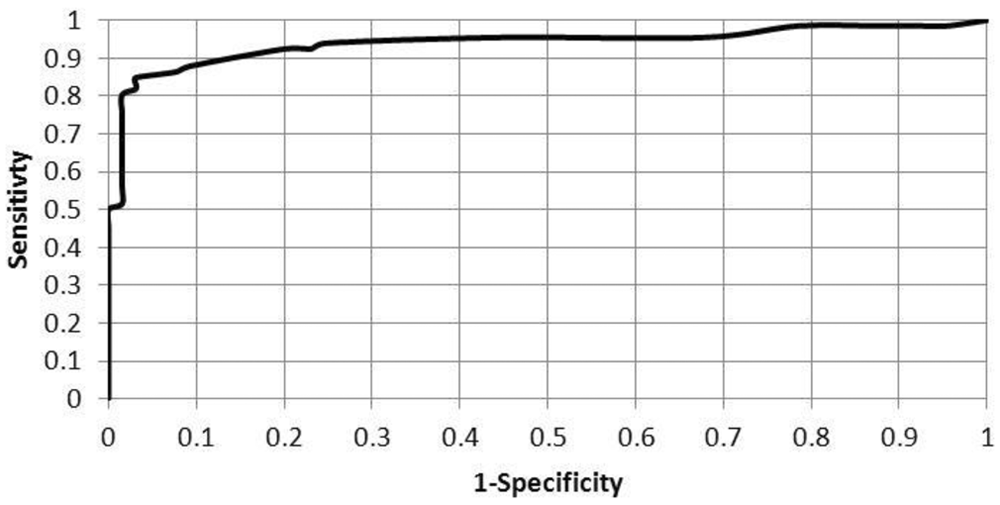
Receiver operating characteristic calculation for modified immunoglobulin G enzyme-linked immunosorbent assay.
The use of chimeric protein A/G conjugate has previously been demonstrated to be an effective means of serological testing in domestic animals. 1 An earlier study explored its use for diagnostic testing in goats, dogs, cats, and pigs using an ELISA method developed in the researchers’ laboratory. 25 In the current study, a commercially produced IgG ELISA kit for the diagnosis of Toxoplasma in human beings was modified to broaden the application of the test to domestic animals that can act as intermediate hosts of the parasite, having the benefit of being easily available to veterinary diagnosticians. Across the species tested, the modified kit produced a relative sensitivity of 92% and specificity of 89% using kit standards for determination of positive, equivocal and negative tests when the IHA was used as a gold standard. At the current time, no commercially available tests in the United States for the diagnosis of toxoplasmosis can be used across the broad spectrum of affected animal species. The current project demonstrates that a human ELISA kit can be modified with the replacement of a non–species-specific protein conjugate for the supplied antihuman IgG protein conjugate and produce very good results for some of the more commonly tested animal species. Non–species-specific protein conjugates, such as protein A/G, have great utility in developing new veterinary diagnostic testing methods for zoonotic diseases in which there are only medical diagnostic methods available.
Footnotes
a.
Toxoplasmosis TPM-Test indirect hemagglutination kit, Wampole Laboratories, Princeton, NJ.
b.
Toxo IgG II ELISA kit, Wampole Laboratories, Princeton, NJ.
c.
Recombinant protein A/G horseradish peroxidase conjugate, Thermo Fisher Scientific, Waltham, MA.
d.
GraphPad Software Inc., La Jolla, CA.
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
This research was made possible by the U.S. Department of Agriculture federal formula fund grant (NYC-478430).