
Abstract
Cutaneous mast cell tumors (MCTs) are among the most common neoplasms in dogs and show a highly variable biologic behavior. Histological grading, cell proliferation markers, and KIT immunohistochemistry are typically used to predict post-surgical prognosis. In the present study, c-KIT messenger RNA (mRNA) expression was measured in canine MCTs and its relationship with tumor grade, immunohistochemical staining pattern, post-surgical prognosis, and mutations was investigated. A significant increase of c-KIT mRNA was observed in MCTs versus healthy skin and surgical margins. Mutations were observed in 8.3% of cases. The KIT staining pattern was investigated for both grading systems. In particular, staining pattern III was associated with grade II (G2) and G3 MCTs, while staining patterns I and II were associated with G1 and G2 MCTs. Considering the 2-tier histological grading, the high grade was mainly associated with pattern III (71%) while the low grade was associated with patterns II (70%) and I (28%). A weak association between the KIT staining pattern and outcome was also observed. The results obtained suggest that c-KIT mRNA is overexpressed in canine MCT, although the fold variations were not associated with the protein localization or complementary DNA mutations. These observations suggested that the 3 events were independent. The histological grading and the KIT staining pattern have prognostic value as previously published. Staining pattern I could be especially helpful in predicting a good prognosis of G2 MCTs. Sequence mutations were not necessarily suggestive of a worse prognosis, but might be useful in choosing a chemotherapy protocol.
Keywords
Introduction
Dogs frequently develop solitary and multiple mast cell tumors (MCTs) that account for approximately 6% of all canine neoplasms and 20% of skin tumors.5,21 In human beings, cutaneous mastocytosis is rare and benign, 7 while the biological behavior of canine MCTs is often difficult to predict.5,34 Some MCTs are benign, develop slowly, and persist for years, while other MCTs may have malignant behavior and exhibit rapid progression, metastases, and eventual death. 21 The etiology of MCTs is still unknown but it is presumably multifactorial with a well-documented breed genetic predisposition (e.g., Boxers) that may influence individual susceptibility to the tumor and increased aggressiveness.41,42 Even the anatomic location has an effect on prognosis, as MCTs that originate in the nail bed, oral cavity, muzzle, or inguinal, preputial, perineal, and mucocutaneous areas often have a worse clinical prognosis. 41
The Patnaik histological grading system 26 and clinical staging are the most frequently used and internationally recognized MCT prognostic parameters.6,34 Histologically, MCTs are graded as well (grade I, G1), moderately (grade II, G2), or poorly differentiated (grade III, G3) tumors. Grade III MCTs are usually more aggressive and have a greater possibility of metastasis than G2 MCTs. 12 The clinical staging classifies MCTs by the extent of their growth. Stage 0 is usually assigned to tumors that are incompletely excised from the dermis or lack regional lymph node involvement. Stages I and II encompass single tumors that are confined to the dermis without or with lymph node involvement. Stage III neoplasms include multiple dermal tumors or large, infiltrating tumors without or with lymph node involvement. Stage IV includes neoplasms with distant metastases. 34 Despite increasing use, the prognostic aspects of histological grading are still subject to debate. 10 This is especially the case for G2 MCTs that may have considerable variation in biological behavior. 31 Assignment of MCT histological grade has varied among pathologists, and these dissimilarities mostly depend on the subjective parameters such as invasiveness, cellularity, and cellular morphology. 41 To improve concordance among pathologists and to provide better prognostic significance, a 2-tier histologic grading system has been proposed. 9
Other prognostic markers including microvessel density, 27 mitotic index,27,30 and markers of cellular proliferation and growth rate, such as argyrophilic nucleolar organizer regions, proliferating cell nuclear antigen, and MKi-67 immunoreactivity 31,32,40 have been investigated in canine MCT.
The study of the c-KIT receptor (KIT or cluster of differentiation [CD]117) has been recently done to explain the pathogenesis of canine cutaneous MCTs. 20 This receptor is a surface growth factor that is normally expressed in mast cells (MCs) and is encoded by the proto-oncogene c-KIT. This gene encodes for a transmembrane type III tyrosine kinase which is the receptor for stem cell factor. The protein consists of an extracellular domain with 5 immunoglobulin-like domains (encoded by exons 1–9), a transmembrane domain (exon 10), and an intracellular domain (exons 11–21). The intracellular domain is further divided into a negative regulatory juxtamembrane domain (exons 11 and 12) and a cytoplasmic tyrosine kinase domain that is split by an insert into adenosine 5’-triphosphate (ATP)-binding (exon 13) and phosphotransferase lobes (exon 17). 15 Ligand binding triggers receptor dimerization, activating the tyrosine kinase, and results in autophosphorylation. This cascade of events causes phosphorylation of exogenous substrates leading to downstream signal transduction phenomena with subsequent proliferation, migration, and differentiation of MCs, hematopoietic stem cells, germ cells, and melanocytes. 16
A ligand-independent mechanism has been proposed to explain MC proliferation. 1 Two types of alteration were hypothesized. First, the c-KIT gene amplification and/or overexpression might increase the cell response to normal growth factor levels or the occurrence of receptor dimerization, even in the absence of the activating ligand. 43 Second, c-KIT genetic modifications of the juxtamembrane and tyrosine kinase domains (i.e., mutations, insertions, deletions) might enhance the constitutive phosphorylation of KIT receptor, resulting in ligand-independent KIT activation and ensuing tumor development. 20
At present, KIT immunohistochemistry reactivity and its pattern of distribution have been used as diagnostic criteria for canine MCTs. 10 Normal and abnormal patterns of KIT expression have been described including a “surface-associated” or membranous pattern with an immunopositivity of the cell membrane, a cytoplasmic perinuclear pattern where KIT is detected in the cytoplasm of neoplastic MCs close to the nucleus, and a diffuse pattern where MCs have diffuse KIT expression throughout the cytoplasm.20,22,27 The presence of aberrant cytoplasmic KIT localization has been correlated with a reduced post-surgical survival.10,25
A large number of c-KIT mutations have been identified in canine MCTs, mostly localized in exon 11,8,15,19,29,38,40,42 occasionally in exons 8 and 9, 15 and sometimes in exon 17. 37 Exon 11 mutations usually consist of internal tandem duplications and have been associated with higher grade MCTs and an increasing incidence of recurrent disease and death. 15
With the exception of a kinetic study in which c-KIT messenger RNA (mRNA) levels were measured in blood samples of dogs suffering from MCTs, 36 no reports concerning c-KIT gene expression in canine MCTs of different histological grades have been published according to the authors’ knowledge. In the present study, the potential mechanism of c-KIT gene amplification/overexpression was evaluated using quantitative real-time reverse transcription polymerase chain reaction. Furthermore, the relationship between c-KIT gene amplification/overexpression and KIT staining pattern, tumor grade, post-surgical prognosis, and mutations were investigated.
Materials and methods
Case selection, tissue specimens, and clinical data
Overall, 60 confirmed cases of spontaneous canine cutaneous MCTs were obtained from veterinary clinics of northern Italy throughout the years 2007–2009. Patients were enrolled in the study after the registration of breed, age, sex, number of tumors, tumor size and anatomical location, and treatments. All dogs underwent surgical excision of the entire tumor, and the diagnosis was confirmed by histological examination. In addition, outcome data, including local or distant recurrence, metastasis, and cause of death, were recorded. A distant recurrence was defined as the development of an additional mass placed far from where the original mass was observed. Outcome time was defined as the time from date of diagnosis to date of the last outcome or death.
For total RNA extraction, aliquots from the central part of the tumor mass (up to 100 mg each) and from the surgical margin (SM; 3 cm wide) were aseptically collected during surgical intervention and immediately stored in RNA stabilization solution a at −20°C until use. Skin samples, obtained from adult pathogen-free Beagle dogs provided by GlaxoSmithKline Research Centre (Verona, Italy) and from adult dogs not suffering from neoplasm and euthanized in veterinary clinics of Padua due to poor general conditions or traumas (i.e., car accidents), were used as controls for the relative quantification of gene expression data. Cutaneous biopsies were collected in sterility up to 30 min after euthanasia and stored in RNA stabilization solution until use.
Histology and immunohistochemistry
After surgical excision, all MCTs were fixed in 10% neutral buffered formalin, routinely processed, and paraffin-embedded. Four-micron serial sections were cut for hematoxylin and eosin staining and immunohistochemical detection of KIT receptor. The tumor grade was determined according to Patnaik 26 by consensus of 3 pathologists (Vascellari, Carminato, and Vercelli). Furthermore, the recently proposed 2-tier histologic grading system (low and high grade) for MCTs was applied. 9
Immunohistochemistry was carried out by an automated immunostainer. b Sections were deparaffinized in xylene, rehydrated in graded ethanol, and rinsed in distilled water. The heat-induced antigen retrieval was performed in 10 mM of citrate buffer (pH 6.0) at 98°C. Endogenous peroxidases were neutralized by incubating the sections in 3% H2O2 in phosphate buffered saline (PBS). Sections were incubated with 1:50 diluted polyclonal rabbit anti-human KIT receptor primary antibody c for 15 min at room temperature. A horseradish peroxidase system b and 3,3’-diaminobenzidine tetrahydrochloride were used as detection system and chromogen, respectively. Sections were then counterstained with Mayer hematoxylin. The specificity of the immunostaining was verified by incubating sections with PBS instead of the specific primary antibody. Three KIT receptor patterns of localization were considered: a) perimembrane (pattern I); b) focal to stippled cytoplasmic (pattern II); and c) diffuse cytoplasmic (pattern III). Each MCT was classified based on the prevalent staining pattern of the neoplastic cell population.
Total RNA isolation and reverse transcription
Total RNA was isolated using TRIzol reagent d according to the manufacturer’s instructions. Samples were then purified with a classical phenol-chloroform e extraction step. Total RNA concentration and quality (260/280 and 260/230 nm absorbance ratios) were measured by using a spectrophotometer f and a denaturing gel g electrophoresis. To generate complementary DNA (cDNA), 2 μg of total RNA and a commercial cDNA transcription kit a were used, following manufacturer’s instructions. Complementary DNA was finally stored at −20°C until use.
Quantitative real-time polymerase chain reaction
The cDNA was used as a template for the c-KIT qPCR assay. The c-KIT primer pairs and the appropriate human Universal ProbeLibrary h (UPL) probe were selected from the extracellular domain (exon 6, which does not represent a hot spot for mutations) of the canine c-KIT mRNA sequence, published in the Ensembl Genome Browser database (ENSCAFT0000 0003274), by using UPL Assay Design Centre web service default parameters. The canine c-KIT transcript was amplified with 5’-CCTTGGAAGTAGTAGATAAAGGATTCA-3’ (forward primer i ), 5’-CCCAATTTGCAACCTTGAAC-3’ (reverse primer), and number 60 human UPL. Calibration curves, using a 4-fold serial dilution of a cDNA pool, revealed PCR efficiencies close to 2 and error values < 0.2. The canine CGI-119 and GOLGA1 were chosen as reference genes for the absence of tissue- and pathological-dependent differences in their mRNA expression. 14 The amplification efficiency of target and reference genes was approximately equal. No statistically significant differences in the reference genes expression profile of healthy and pathological samples were observed.
Quantitative PCR was performed using a commercial instrument h using standard PCR conditions. A volume of 2.5 µl of 1:50 diluted cDNA and the 1× hot start reaction mix, h containing 100 nmol/l of the selected human UPL probe, were used. Data were analyzed and quantified with the instrument software, h by using the second derivative maximum method. 28
For the relative quantification of c-KIT mRNA, the ΔΔCt method was used. 17 The relative quantification values of tumors and SMs were expressed as fold changes, which are normalized to the ΔΔCt mean value of control skin samples to whom an arbitrary value of 1 was assigned.
Amplification and sequencing of c-KIT exons 8, 9, and 11
Exons 8, 9, and 11 were chosen to disclose c-KIT mutations, as they represent the most common and frequent sites of c-KIT mutations. 15 Two microliters of 50-fold diluted cDNA were used as a template for the PCR amplification. Reactions were carried out in a thermocycler j by using 16.5 pmol of each primer, i 1.5 U of DNA polymerase, k and 200 μM deoxyribonucleotide triphosphate mix, k 1.5 mM MgCl2, k and 1× reaction buffer k (final concentrations). Primers pairs used are reported in Table 1. Oligonucleotides for exons 8 and 9 were designed ex novo using Primer3 software (http://primer3.sourceforge.net/), while for exon 11, published primers were used. 42 The following PCR conditions were used: an activation step at 95°C for 2 min, 35 cycles of 30 sec at 95°C, 45 sec at 55°C or 65°C (for exons 8, 9, and 11, respectively), 45 sec at 72°C, and a final extension step of 5 min at 72°C. Amplicons were visualized in a 2% agarose g gel electrophoresis. When 2 bands, different in length of approximately 50 bp, were obtained for the presence of internal tandem duplication or deletion, bands were first individually excised and then purified using a commercial kit. h If only 1 band was obtained, the PCR product was purified according to the manufacturer’s protocol. All the purified templates were directly sequenced on an automated sequencer, a by using a commercial cycle sequencing kit a according to the manufacturer’s instructions. Unincorporated terminators were removed by isopropyl alcohol e precipitation. Samples were sequenced starting from both forward and reverse primers used for the PCR amplification. Sequences were finally compared by using the software ChromasPro 1.5. 1
Primers for polymerase chain reaction amplification and sequencing of c-KIT exons 8, 9, and 11.
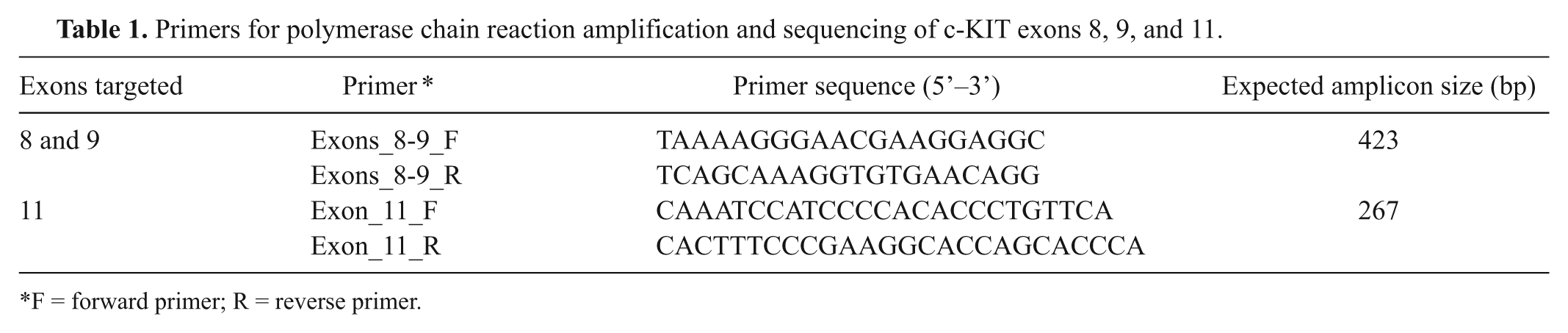
F = forward primer; R = reverse primer.
Statistical analysis
To evaluate the distribution of the KIT staining pattern compared to the histological grading and outcome data, the Fisher exact test was used. The prognostic significance of the KIT staining pattern was quantified by means of negative and positive predictive values, considering the prognosis as the gold standard.
To evaluate the distribution of the c-KIT gene expression based on the 2 histological grading systems and the prognosis, and to compare the mRNA expression in MCTs and control skin samples and SMs, respectively, the Mann–Whitney test was used. Given the low size of the G3 MCTs (n = 3), the analysis was performed considering G1 versus G2, and G1 versus G2 and G3. The comparison of c-KIT mRNA expression in MCTs and SMs was performed using the Wilcoxon signed-ranks test. The survival time was investigated by means of Kaplan–Meier survival analysis, stratified by KIT patterns, and the log rank test was used to compare the survival among groups. For all analyses, the software STATA v. 9.2 m was used, and a value of P < 0.05 was considered as statistically significant.
Results
Clinical and pathological features of tumor patients
Overall 60 MCTs were collected from 59 dogs. Both male (22) and female (34) dogs were included in the study. The sex was not available for 3 cases. The mean age at surgery was 8 years (range: 3–13 years). The following breeds were represented: crossbred (17), Boxer (10), Labrador Retriever (10), English Setter (3), Golden Retriever (3), Dachshund (2), Cocker Spaniel (2), Dogo Argentino (2), American Staffordshire Terrier (1), Boston Terrier (1), Bouvier des Flandres (1), Bulldog (1), Cane Corso (1), Dalmatian (1), pit bull (1), and Shi Tzu (1). For 2 dogs, the breed was unknown.
According to the Patnaik classification, 32 G1, 25 G2, and 3 G3 MCTs were diagnosed. The distribution of the cases according to the 2-tier histologic grading system consisted of 53 low-grade (32 G1, 21 G2) and 7 high-grade (4 G2, 3 G3) MCTs.
Fifty-one out of the 59 dogs that underwent a mean follow-up period of 18 months (range: 3–36 months) were considered for prognostic evaluations. Thirty-eight patients were still alive at the end of the study period (follow-up range: 12–36 months); 3 of the dogs showed development of an additional MCT at a site distant from that in which the original mass was observed, and were still alive at the end of the study. Eight dogs showed clinical signs of MCT recurrence at the initial tumor site and died or were euthanized for progressive MCT disease. The mean survival time was 8.5 months (range: 3–13 months). Five patients died for other diseases, not correlated with MCT.
KIT immunohistochemistry
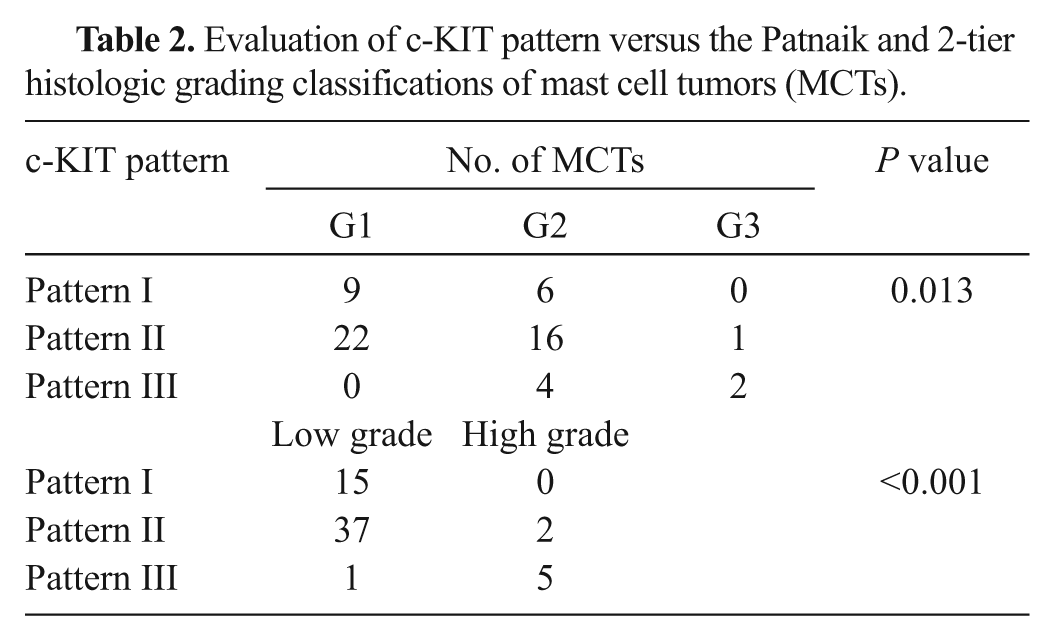
Fifteen MCTs showed a predominant KIT staining pattern I (25%; Fig. 1A), 39 MCTs a staining pattern II (65%; Fig. 1B), and 6 a diffuse cytoplasmic pattern III (10%; Fig. 1C). The distribution of the staining pattern in G1, G2, and G3 or low- and high-grade MCTs is reported in Table 2. A significant association between the KIT staining pattern and the Patnaik grading system was observed (P = 0.013). Particularly, staining pattern III was strongly associated with the G2 and G3 MCTs, while both patterns I and II were mainly associated with G1 and G2 MCTs. Considering the 2-tier histological grading, a significant association was observed between grading and KIT staining pattern (P < 0.001): the high-grade was mainly associated with pattern III (71%) while the low-grade was associated with patterns II (70%) and I (28%).
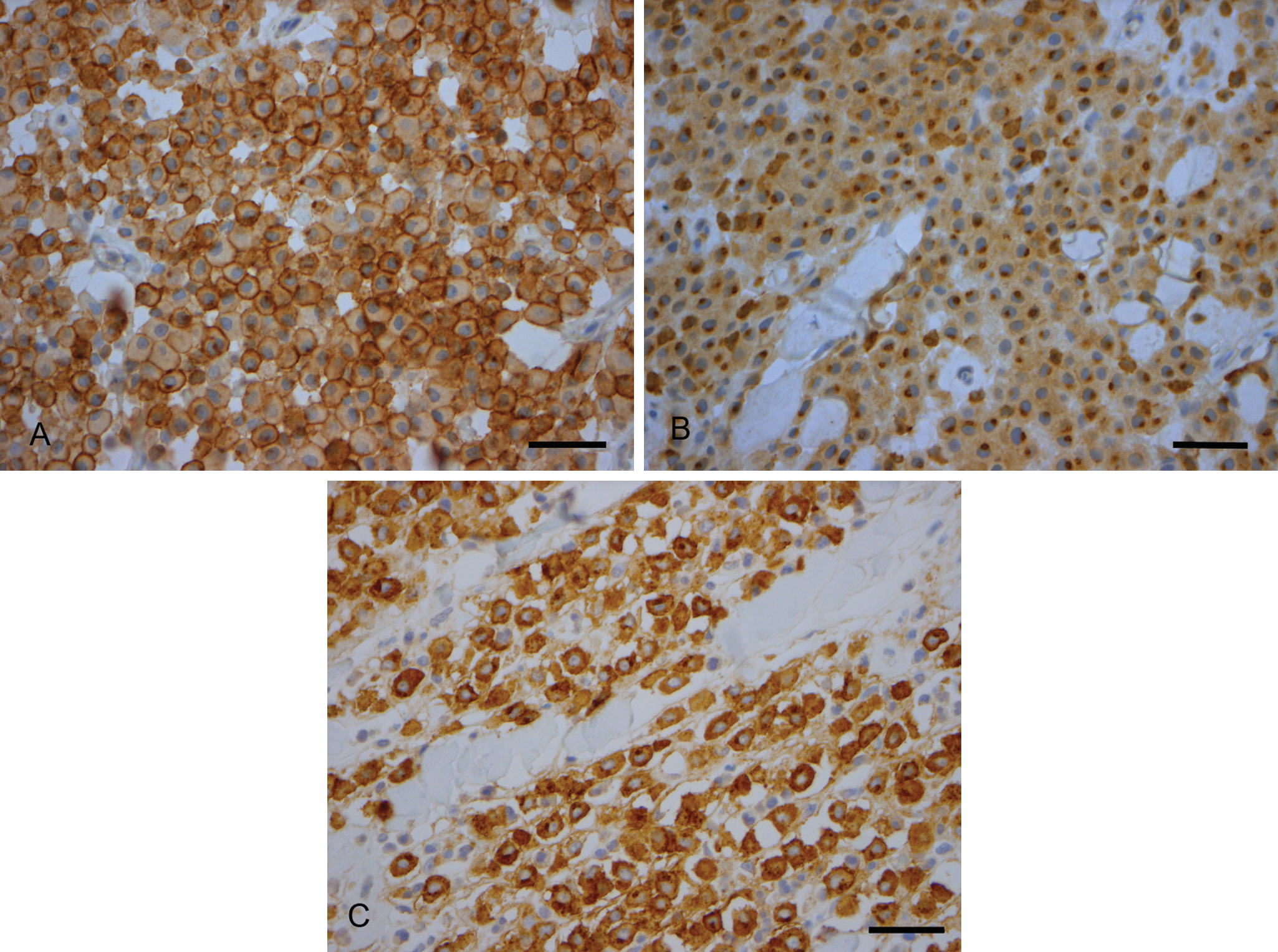
Immunohistochemical staining of c-KIT in mast cell tumors. Three representative images of c-KIT pattern protein localization. KIT receptor polyclonal:
Evaluation of c-KIT pattern versus the Patnaik and 2-tier histologic grading classifications of mast cell tumors (MCTs).
A weak association between the KIT staining pattern and the local recurrence or survival time was also observed (P = 0.06). It is noteworthy that 7 out of 8 dead dogs were affected by MCT with patterns II (n = 4) and III (n = 3), while 82% of dogs with a KIT staining pattern II or III were still alive 12 months after surgery. Thus, in the current survey, the cytoplasmic localization of KIT (patterns II and III) was not a good predictor of a worse prognosis (Positive Predictive Value [PPV] = 17.5%, 95% confidence interval [CI]: 7.34–32.8%), while the perimembranous localization of KIT was indicative of a good prognosis (Negative Predictive Value [NPV] = 92.3%, 95% CI: 64–99.8%).
The survival probabilities at 12 months were 0.92 (n = 13, 95% CI: 0.56–0.98), 0.94 (n = 34, 95% CI: 0.77–0.98), and 0.11 (n = 6, 95% CI: 0.11–0.80) for patterns I, II, and III, respectively. Considering the entire outcome period, the 3 groups showed different survival curves (P = 0.0054; Fig. 2).
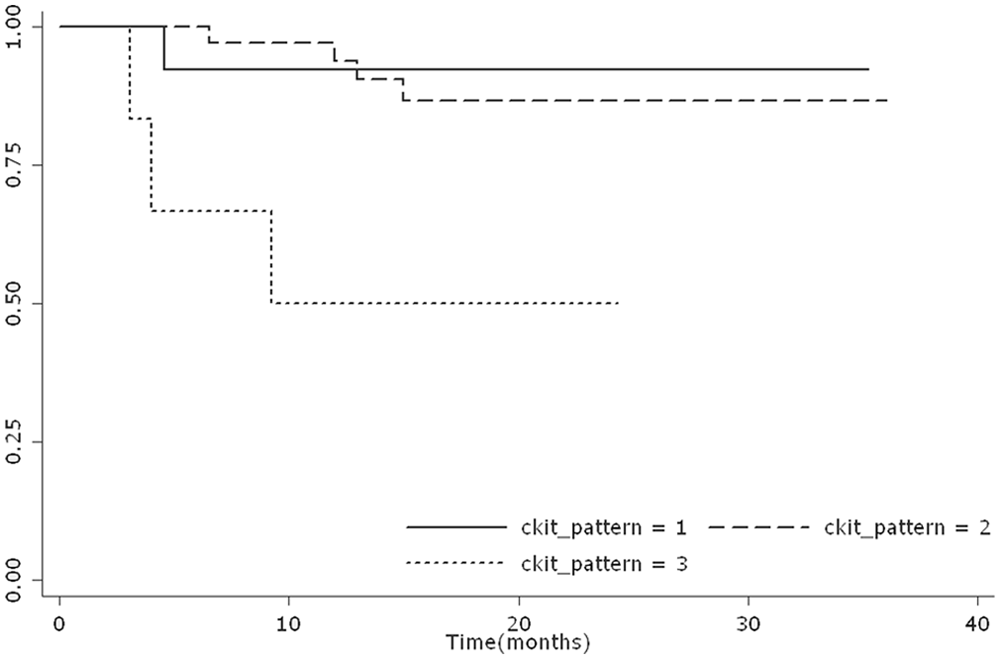
Survival curve of dogs with different c-KIT staining patterns. Kaplan–Meier survival plot stratified by different c-KIT staining patterns.
c-KIT messenger RNA expression
The c-KIT mRNA expression was measured, through a qPCR approach, in control skin tissue, in 30 SM biopsies and 59 MCTs. The transcript was amplifiable and quantifiable in all the samples examined.
A statistically significant increase in the c-KIT gene expression was observed between the MCTs and the control skin samples: the median value was equal to 71.98- and 0.86-fold changes (arbitrary units), respectively (P < 0.001; Fig. 3). Similar results were obtained comparing the MCTs and SM (P < 0.001; Fig. 4A). In contrast, the c-KIT gene expression in SM (median value of 1.23-fold changes) and control skin tissue was almost equal (Fig. 4B).
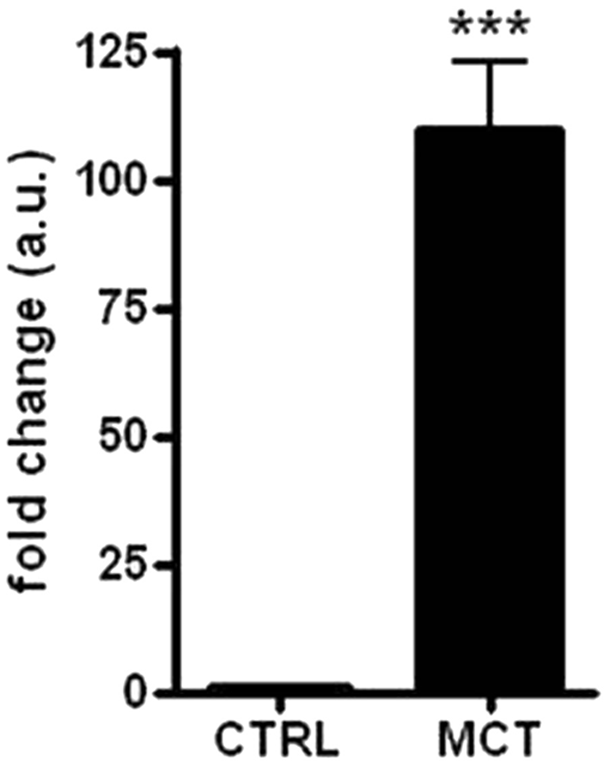
c-KIT messenger RNA (mRNA) expression in skin compared with mast cell tumors. Level of c-KIT mRNA expression in control skin tissue (CTRL) and in tumor samples (MCT). Data of relative quantification (RQ), obtained by using the ΔΔCt method, were transformed in fold changes (means ± standard error of measurement of arbitrary units), which is normalized to the mean RQ of control group (skin of healthy dogs). Statistical analysis: Mann–Whitney test. *** P < 0.001.
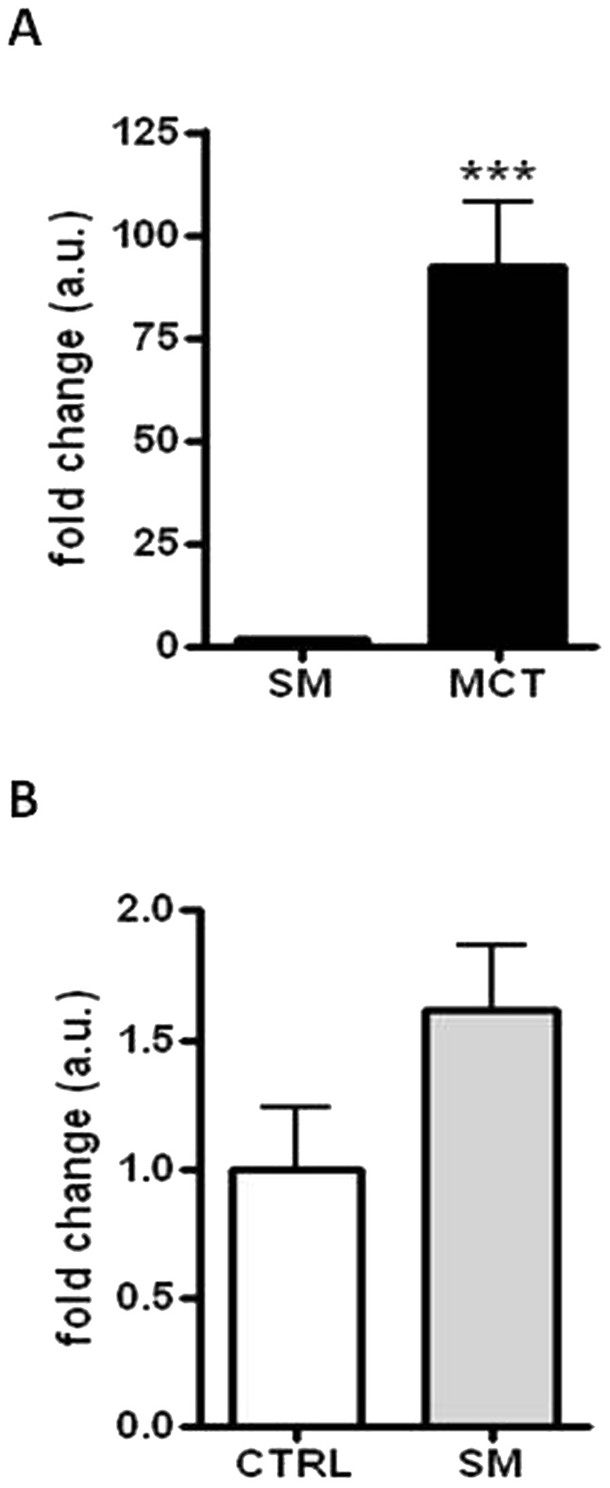
c-KIT messenger RNA (mRNA) expression in surgical margins compared with mast cell tumors (MCTs) and skin. Level of c-KIT mRNA expression in surgical margins (SM) compared to MCTs (A) and control skin tissue (CTRL: B). Data of relative quantification (RQ), obtained by using the ΔΔCt method, were transformed in fold changes (means ± standard error of measurement of arbitrary units), which is normalized to the mean RQ of control group (skin of healthy dogs). Statistical analysis: Mann–Whitney test. *** P < 0.001 (
Taking into account the 2 histological grading systems, median values of c-KIT gene expression were 52.69-, 90.79-, and 124.14-fold higher than control skin tissue in G1, G2, and G3 MCTs, respectively; in contrast, 71.81- and 124.14-fold values were noticed in low- and high-grade MCTs. Despite the increase of mRNA levels among histological subgroups, a significant association between c-KIT gene expression and the MCT histological grade was never observed (see Table 3).
c-KIT messenger RNA (mRNA) expression in mast cell tumors (MCTs) following the 2 criteria of histologic grading classification.*
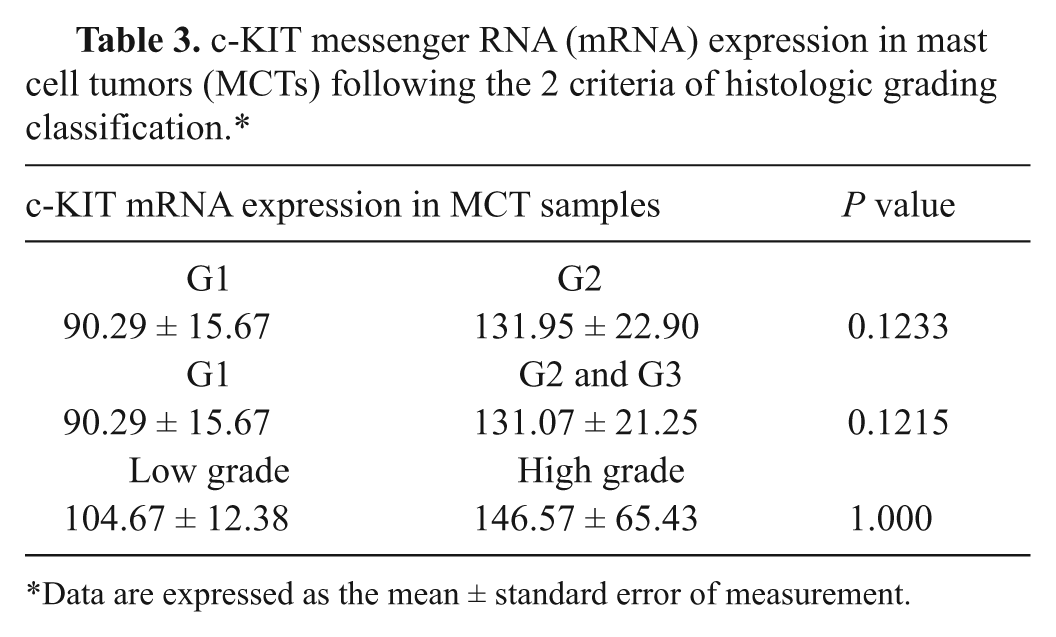
Data are expressed as the mean ± standard error of measurement.
Furthermore, in terms of health status, alive and dead dogs showed a mean value of c-KIT mRNA amount of 71.64- and 122.24-fold versus control tissue, respectively. No significant relationship between c-KIT gene expression and disease recurrence as well as overall survival time was observed.
c-KIT mutations
The presence of alterations in the c-KIT cDNA sequence was investigated in 59 tumor biopsies. A portion of the c-KIT extracellular ligand-binding domain (exons 8 and 9) as well as of the juxtamembrane domain (exon 11) was amplified. The agarose gel electrophoresis separation of PCR products showed that 3 out of 59 tumors contained a larger band in addition to the expected one. An extra band, below the expected one, was also noticed in an additional sample. These findings were respectively suggestive of possible duplications and 1 deletion. In Figure 5, an agarose gel with a duplication in exon 11 is shown. The aforementioned bands were individually excised, purified, and sequenced; this same approach was applied to all of the PCR products of exons 8, 9, and 11, to screen the potential presence of mutations too small to be seen as separate bands. As a result, 1 additional mutation was found.
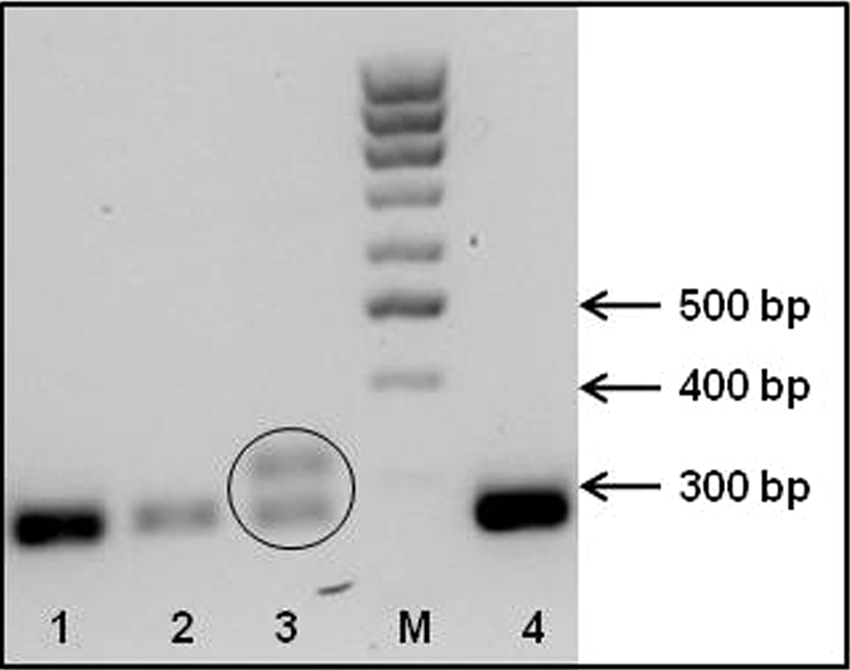
Agarose gel electrophoresis of c-KIT exon 11 polymerase chain reaction (PCR) products. Representative 2% agarose gel showing PCR products obtained by the amplification of c-KIT complementary DNA (exon 11) in different mast cell tumors. Lanes 1, 2, and 4 contain the wild-type sequence. In lane 3, the lower band corresponds to the wild-type sequence, and the upper one to a duplication. Lane M contains a 100-bp DNA ladder.
As a whole, mutations in the c-KIT cDNA were observed in 5 tumors (8.5%); in particular, 1 in exons 8 and 9 and 4 in exon 11 (Figs. 6, 7). Two mutations were identified in patients with G1 MCTs (2/31, 6.5%), 2 in patients suffering from a G2 MCT (2/25, 8.0%), and 1 in a G3 MCT (1/3, 33.3%). Considering the alternative histological classification, 4 and 1 mutations were recorded in low- and high-grade MCTs, respectively. In exons 8 and 9 (see nucleotide and protein sequences in Fig. 6), a 12-bp internal tandem duplication (ITD), resulting in the insertion of the amino acid sequence QILT at the residue 421 (ITD417-421) was observed, as previously described. 15
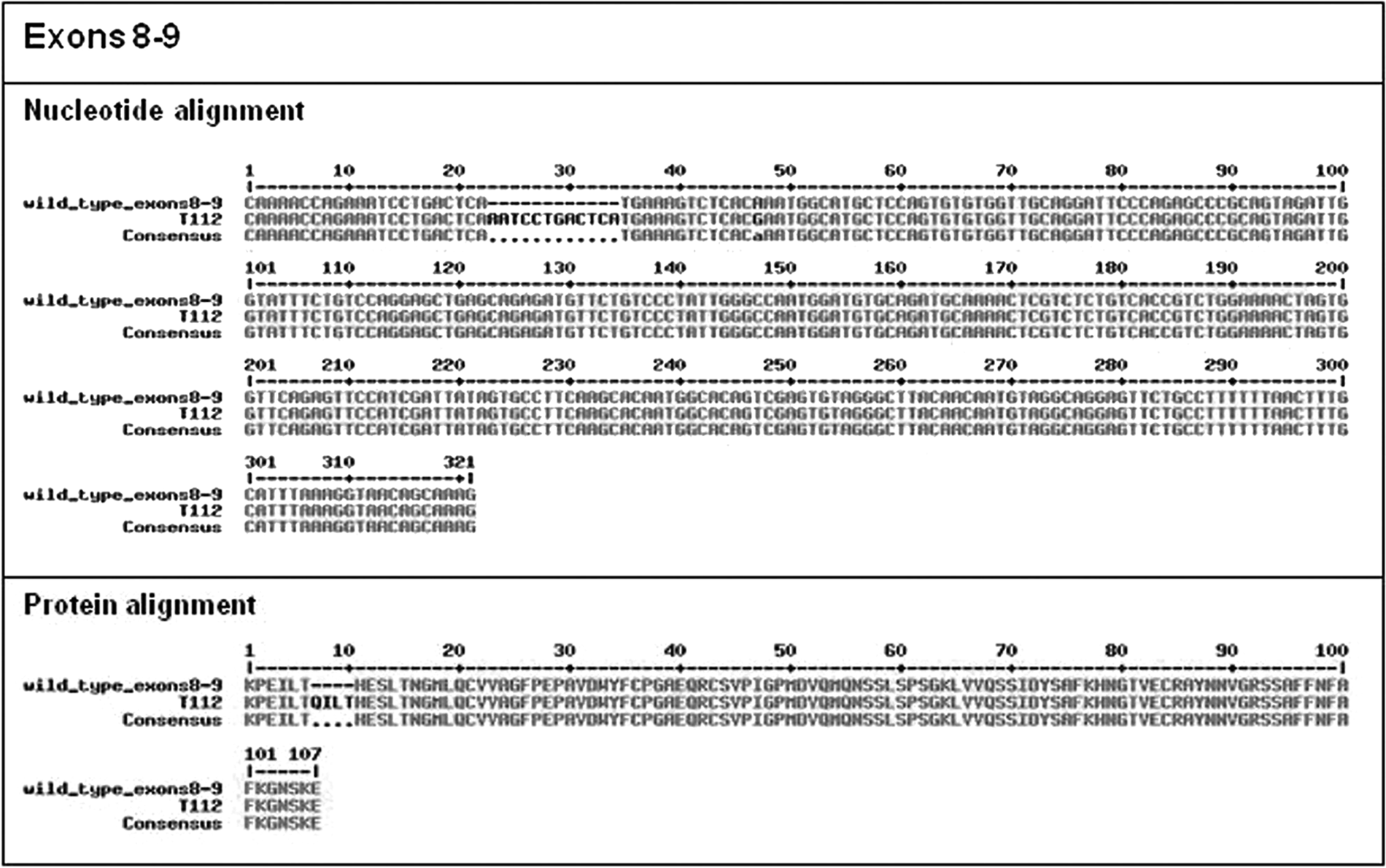
c-KIT mutations of exons 8 and 9 evidenced in the present study: nucleotide and protein alignment of wild-type and mutated sequences. Nucleotide and amino acid alignment of c-KIT (exons 8 and 9) in the wild-type and mutated (T112 mast cell tumor) sequences. In T112, the insertion of the amino acid sequence QILT in exon 8 at residue 421 (ITD417-421) was noted.

c-KIT mutations of exons 11 evidenced in the present study: nucleotide and protein alignment of wild-type and mutated sequences. Nucleotide and amino acid alignment of c-KIT (exon 11) in the wild-type and mutated (T53, T90, T106, and T132 mast cell tumors) sequences. An internal tandem duplication (ITD) of residues 576–594, 576–590 (ITD2), 41 and 577–588 in T53, T106, and T132 were observed, respectively. A further deletion of 9 amino acid residues, comprised between 553 and 561, was identified in T90.
In the c-KIT juxtamembrane domain, 2 duplications in frame of 45 bp (corresponding to 15 amino acids) and 1 of 36 bp (corresponding to 12 amino acids), located near the 3’ end of exon 11, were noticed (see nucleotide and protein sequences in Fig. 7). In detail, an ITD causing a tandem repeat within the protein sequence of residues Pro576-Asn590, corresponding to the Dup2 previously described, 42 was observed; furthermore, an ITD of residues Pro580-Phe594 and an ITD of residues Thr577-Pro588 were herein identified. Finally, a deletion encompassing 27 bp at the 5’ end of exon 11, with a consequent 9 amino acids deletion (Lys553-Lys561), was also detected. This latter mutation confirmed what has been previously hypothesized. 42
Five patients displayed c-KIT mutations, but recurrence and death were observed in only 1 case corresponding to a G3 or a high-grade MCT, localized at the scrotum and characterized by a pattern III of c-KIT protein localization. The other 4 MCTs with c-KIT sequence alterations were characterized by c-KIT staining pattern II.
Discussion
The goal of the current study was to evaluate potential c-KIT gene amplification and/or overexpression in canine MCTs and investigate its relationship with the histological grade, c-KIT protein receptor localization, alterations in the c-KIT cDNA sequence, and clinical outcome data. Several attempts have been made to develop alternative methods to clarify the biological behavior of MCTs. 41 A qPCR approach is considered a reliable, highly sensitive, rapid, and economic tool to obtain quantitative data. Its usefulness has been evaluated in this respect.
To date, few reports have been published that use qPCR to quantitate c-KIT mRNA in canine tumors.13,36 In a previous study, c-KIT mRNA was measured in the blood of MCT-affected dogs. 36 The study 36 detected c-KIT mRNA expression in tumor biopsies as well as in most blood specimens, although at lower levels. The amount of mRNA was highest in blood at the time of surgery, decreased progressively between 1 and 3 months after surgery, and remained persistently low. No correlations were found between c-KIT gene expression in blood and tumor grading, degree of neoplasm differentiation, or clinical prognosis. 36 In the present study, c-KIT gene expression was measured in MCT biopsies of different histological grades and compared with the level of expression detected in SM and control skin biopsies. A significant increase in c-KIT gene expression was present in MCTs if compared with both SM and control skin samples. This result would be consistent with the ligand-independent mechanism of MC proliferation, such as c-KIT gene amplification and/or overexpression. 43 However, evaluation of mRNA data versus health status in the present study lacked statistically significant differences between dead and living dogs. Therefore, c-KIT gene expression did not seem to be predictive of biological behavior and only indicated proliferation of MCs.
At present, histological grading is often used to predict the prognosis of canine MCT, but the ambiguity of intermediate-grade tumors and the diagnostic expertise of pathologists have led to questions concerning the relevance of the current histological grading system.4,11,39 Thus, new proposals for more accurate prediction of the biological behavior of canine cutaneous and subcutaneous MCTs have been formulated.9,35 In the present study, the Patnaik and 2-tier histological systems were used to classify MCT cases, and both systems were compared with post-surgical clinical prognosis. Both histological classifications were associated with clinical prognosis and KIT immunohistochemical staining pattern as previously reported.4,10,27,31,40 In contrast, no statistically significant differences in c-KIT gene expression were observed among G1, G2, and G3 or low- and high-grade MCTs. Thus, it was not possible to establish grade-specific cut-off points or to assign thresholds of minimum and maximum c-KIT mRNA amounts for each tumor grade. This problem might be the result of high variability in c-KIT mRNA expression observed in groups G2, G3, and high-grade, or to the small number of neoplasms analyzed (especially G3 and high-grade MCTs). Therefore, evaluation of more neoplasms might be helpful to determine if c-KIT gene overexpression could be a key factor in neoplastic progression, as previously hypothesized. 25
The immunohistochemical staining of KIT receptor protein is still considered one of the most useful prognostic parameters in canine MCTs. Previous studies4,10,25,38,39,42 often showed a strong correlation between altered (cytoplasmic) KIT expression and increased cellular proliferation, higher histological grade, presence of c-KIT mutations, increased local tumor recurrence, and/or decreased clinical survival. In the current survey, the KIT staining pattern was associated with histological differentiation but was only weakly associated with the post-surgical prognosis. The KIT staining pattern I was strongly associated with a good prognosis. Dogs with MCTs characterized by KIT patterns I and II had similar survival curves, while dogs with pattern III had a lower survival time, even if the CI was wide. Thus, altered cytoplasmic localization of the KIT protein did not seem to be predictive of a worse prognosis; however, this observation might be influenced by the small number of dogs that died in the present survey. These findings are in contrast with previous studies that highlighted the prognostic value of cytoplasmic KIT staining in association with an increased rate of tumor recurrence and decreased survival time. 10 Furthermore, results of the present study indicate that KIT staining pattern is not related to an overexpression of c-KIT mRNA. In a previous study, no correlation between KIT protein expression and KIT protein localization was observed in a tissue microarray using an immunofluorescence approach. 38 Taken together, these data suggest that the level of c-KIT mRNA expression is probably independent of protein localization, tumor differentiation, and tumor progression. Further studies will be required to understand the molecular mechanisms underlying KIT pathways.
Alterations in the extracellular and juxtamembrane domains of c-KIT, which represent well-known hot spots for c-KIT mutations, were also considered in the current study. The analysis was performed on the cDNA sequence to highlight ITD, deletions, or mutations causing functional alterations. Mutations of the c-KIT sequence are usually found in 9–15% of MCTs,38,42 and in the present study, the mutations accounted for 8.3%. One alteration in exons 8 and 9 and 4 alterations in exon 11 were observed. Among them, 2 ITDs and 1 deletion have been previously described,15,42 while the ITD of residues Pro580-Phe594 and Thr577-Pro588 are novel findings. Previously, the presence of mutations were associated with the tumor grade,2,29,38,42 aberrant KIT protein localization, increased incidence of tumor recurrence, and MCT-related death. 38 No clear relationships between the presence of c-KIT mutations and the above-mentioned variables were obtained in the present study. In fact, mutations were found in two G1, two G2, and one G3 MCTs, or in 4 low-grade and 1 high-grade MCT. Only 1 tumor revealed pattern III of KIT protein localization, while all the other tumors showed pattern II localization. In this respect, it has been previously shown that an aberrant pattern of distribution of KIT protein may be present even without concurrent c-KIT mutations 38 ; consequently, the previous study hypothesized that ITD or deletions and changes in KIT localization might represent separate events, occurring independently in the progression of MCTs. In the present work, only 1 patient with c-KIT mutation died because of the tumor. In contrast, 7 patients characterized by a wild-type c-KIT sequence died of MCT or were euthanized due to poor prognosis. Altogether, the results confirm that c-KIT mutation-negative MCTs could proliferate and behave in a malignant manner. 23 Alternatively, the presence of c-KIT mutations could aid the veterinary oncologist to choose an appropriate chemotherapeutic regimen. Furthermore, the use of tyrosine kinase inhibitors may be considered because they apparently are effective against c-KIT mutation-positive MCTs. 18
Several studies have described a close relationship between SM and prognosis or recurrence of G1 and G2 MCTs. 24 In general, it is believed that tumor excision using a 2-cm lateral and a deep margin of one fascial plane are adequate to completely excise G1 and G2 MCTs in dogs.3,33 However, many veterinary oncologists ask for more accurate prognostic factors in cases of incomplete surgical excision. In the present study, the classical histological evaluation of MCT SM and c-KIT gene expression profile were measured concurrently and might be of prognostic usefulness.
In conclusion, the findings of the present study suggest that c-KIT mRNA is overexpressed in canine MCTs although the levels of gene expression, protein localization, and c-KIT mutations are probably 3 independent events. Furthermore, c-KIT mRNA overexpression may to play a role in MC malignant transformation and proliferation, but apparently does not indicate the biological behavior of MCTs. In contrast, histological grading and KIT staining patterns confirm their prognostic value, particularly the association of membrane-associated protein localization and the presence of a good prognosis for intermediate grade MCTs.
Footnotes
Acknowledgements
The authors would like to thank Prof. Ilaria Iacopetti and Drs. Helen Poser, Barbara Carobbi, and Patrizia Cristofori for their contribution in providing part of tumor and control samples included in the present investigation. Part of this work has been presented at the ESVONC Annual Congress 2010 (Turin, Italy, March 19–20).
Notes
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
This work was supported by grants from Istituto Zooprofilattico Sperimentale delle Venezie (RC IZSVE 09/06) to Marta Vascellari, Anna Granato, and Mauro Dacasto.