
Abstract
Although neurological signs have been reported sporadically in dogs with systemic Lyme disease, it is unknown if neuroborreliosis occurs in dogs. The current study systematically evaluates canine brains for evidence of Borrelia burgdorferi infection. Twelve Beagles were experimentally challenged with B. burgdorferi–infected ticks at 18 weeks of age, and 2 unexposed dogs served as controls. One of the uninfected dogs and 6 infected dogs were each given 5 daily immunosuppressive doses of dexamethasone starting at 153 days post-infection. Eleven dogs were confirmed as infected by skin punch biopsy polymerase chain reaction (PCR) and serology. Neurological signs were not seen in any dogs through the end of the 190-day study. Whole blood, serum, cerebrospinal fluid (CSF), and brains from all dogs were collected. DNA was extracted from blood, CSF, and brain and evaluated by PCR for B. burgdorferi. Formalin-fixed brain tissue was examined by histopathology, immunohistochemistry, and PCR. Immunohistochemical staining for B. burgdorferi antigen was negative in all cases. The CSF analysis was normal, and PCR was uniformly negative for B. burgdorferi in all dogs. Six of the 11 (45%) infected dogs had mild to moderate lymphoplasmacytic choroid plexitis, which was more pronounced in the immunosuppressed dogs. The lack of B. burgdorferi DNA and immunohistochemical evidence of organisms, including within the choroid plexus lesions, suggests that B. burgdorferi does not have a direct role in the etiopathogenesis of canine central nervous system pathology.
Lyme disease is a zoonotic multisystem disease caused by infection with the Gram-negative spirochete Borrelia burgdorferi. The organism is carried by Ixodes scapularis and Ixodes pacificus ticks affecting many animal species including dogs. 11 Neuroborreliosis is a distinct but uncommon clinical neurological manifestation of Lyme disease in human beings and occurs in up to approximately 10% of chronic, untreated cases in North America. 11 A diagnosis of neuroborreliosis is made when there are signs and symptoms of neurological disease, supporting laboratory findings, and evidence of possible tick exposure. 11,19 Unless clinical signs are manifested, human beings with other symptoms of Lyme disease are not routinely screened for subclinical neurological pathology. 1
Despite sporadic reports of neurological signs occurring in dogs with Lyme disease, whether or not B. burgdorferi infection causes the neurological signs remains controversial. 3,15,17 Variable lesions in skin, lymph nodes, aorta, kidney, and meninges are found in experimentally infected dogs, but the dogs do not manifest consistent cardiac, neurologic, or renal signs. 6,15,21–24 In particular, subclinical central nervous system (CNS) lesions have not been described consistently in naturally or experimentally infected dogs. Experimentally, a mild peripheral perineuritis has been described, 6,24 and a single report of mild focal infiltrations of lymphocytes and plasma cells in the dura mater of 3 dogs and a single small focus of encephalitis with lymphocytic infiltrates and gliosis was reported within the caudal brain stem of 1 dog. 6
Approximately 95% of exposed dogs are asymptomatic even in endemic areas in which 90% of dogs have seroconverted. 14,16 Because of persistent seropositivity following exposure to the organism, in these endemic areas, seropositive dogs with CNS signs cannot be assumed to have neuroborreliosis, and other causes of neurological disease should be investigated. 14–16 The authors of the current study as well as other researchers recently evaluated large groups of dogs with neurological disorders of unknown etiology and failed to detect the presence of Borrelia antibodies and/or DNA in cerebrospinal fluid (CSF). 4,12 Natural Borrelia infections in dogs therefore do not seem to be as commonly associated with meningoencephalitis as it occurs in human beings.
A systematic evaluation of canine brains for evidence of B. burgdorferi infection combining histological evaluation, immunohistochemistry (IHC), CSF analysis, and CSF polymerase chain reaction (PCR) is critically lacking. The goals of the present study were: 1) to determine whether or not clinical signs consistent with neuroborreliosis occur in experimentally infected immunocompetent and immunosuppressed dogs, and 2) to characterize any subclinical pathological features of CNS involvement in dogs infected with B. burgdorferi.
Fourteen male and female purpose-bred Beagles were included in the study. Twelve dogs were challenged at 18 weeks of age with B. burgdorferi–infected ticks using the method described below. Two dogs were not exposed to ticks, and served as negative controls. Infection with B. burgdorferi was confirmed by PCR of skin punch biopsies and serum enzyme-linked immunosorbent assay (ELISA) at 83 days postinfection (DPI) as per the methodology described below. 7,13,22 An additional serum sample was obtained at humane euthanasia for confirmatory C6 peptide antigen evaluation of Lyme status and for co-infection with Anaplasma phagocytophilum.
Dogs were purchased through a commercial supplier a at 8 weeks of age. Dogs received routine core vaccination (Canine distemper virus, Canine adenovirus, Canine parainfluenza virus, Canine parvovirus) and immunization for Bordetella bronchiseptica and were acclimated to the housing facility and handling procedures prior to experimental infection at 18 weeks of age. Dogs were confirmed to be seronegative for Lyme antigen by ELISA at a commercial diagnostic laboratory b prior to tick exposure. Dogs were fed a well-balanced, commercially available adult dog food and water was provided ad libitum. After tick challenge, the dogs were comingled in divided runs in conventional housing fitted with fine-mesh screens with sticky tape at all entrance points to control entry and exit of crawling arthropods. The floors and walls of the facility were treated with insecticide after challenge, and all dogs were treated with a topical insecticide before they were returned to conventional housing for the duration of the study.
After tick exposure, dogs were observed daily for clinical manifestations of Lyme disease. Special attention was given to signs of lameness, fatigue, or malaise. Early humane euthanasia criteria included: non–weight-bearing lameness, lameness persisting greater than 3 days, severe depression, inability to stand, dehydration >10%, body temperature <36.7°C or >40.5°C for more than 2 consecutive days, weight loss >20%, or profoundly diminished sensorium (e.g., moribund dogs). Dogs were humanely euthanized with 65 mg/kg of pentobarbital intravenously using a 21-guage, 0.75-inch (19.1-mm) butterfly catheter placed in the cephalic vein. Animals were handled in compliance with Merial Institutional Animal Care and Use Committee (IACUC) approvals and applicable local regulations.
Wild-caught I, scapularis ticks captured from a B. burgdorferi endemic area, having a reported 66% infection rate with B. burgdorferi, were purchased from a commercial supplier. c Tick exposure procedure is as previously described. 2,13,24 Briefly, 13 adult female and 12 male I. scapularis ticks were placed on each dog near the clipped skin of the left shoulder, confined in a plastic capsule secured to the dogs’ sides with adhesive tape, and allowed to feed to repletion and detachment, which was approximately 4–7 days. Infection with B. burgdorferi was confirmed if the dog was positive by PCR on skin biopsies and serology test 83 DPI as described previously. 2,21–23 Two 5.0-mm skin punch biopsies d were obtained aseptically from all dogs within the area of tick attachment under anesthesia (medetomidine [0.02 mg/kg intramuscularly], butorphanol [0.2 mm/kg intramuscularly], and carprofen [2.2 mg/kg orally]). Biopsy sites were closed with skin staples. Samples were chilled on ice and delivered the same day to a commercial laboratory e for PCR. Blood was collected aseptically from the jugular vein for serologic tests, and serum was shipped on ice overnight for whole antigen Lyme ELISA to a commercial laboratory. b
Starting on day 153 postinfection, 7 dogs (1 uninfected control and 6 infected dogs) were selected at random to receive daily subcutaneous injections of dexamethasone at an immunosuppressive dose of 0.88 mg/kg for 5 consecutive days in order to promote manifestation of clinical signs, and in coordination with the needs for another study, as described previously. 6,13 The remaining 7 dogs did not receive any injections.
At 191 and 192 DPI and immediately prior to humane euthanasia, serum was collected and transported directly to a commercial laboratory e for serological testing f for Lyme disease (B. burgdorferi) and anaplasmosis (A. phagocytophilum), as well as canine heartworm (Dirofilaria immitis) and ehrlichiosis (Ehrlichia canis). At the same time, 1.0 ml of ethylenediamine tetra-acetic acid–anticoagulated whole blood was collected and frozen at −80°C for PCR analysis as detailed below.
Cerebrospinal fluid was collected immediately following humane euthanasia from the cerebellomedullary cistern. The dorsal cervical area was shaved and aseptically prepared. Cerebrospinal fluid was collected routinely using a 22-gauge, 2.5-inch (63.5-mm) spinal needle g and stored in a 1.0-ml sterile cryogenic vial. h Within 4 hr of collection, a 0.5-ml aliquot of CSF fluid was frozen at −80°C for subsequent PCR analysis detailed below. If there was sufficient CSF, an additional 0.5-ml aliquot was used for a routine CSF analysis performed within 3 hr. The CSF analysis included a red blood cell count (RBC), white blood cell count (WBC), total protein, and cytospin evaluation. The RBC and WBC counts were performed manually on a hemocytometer. The CSF protein was measured using Coomassie blue on a commercial analyzer. i The cytospins were evaluated by the same clinical pathologist (Krimer).
Necropsies were performed 1–4 hr after humane euthanasia, and the brain was removed. A 1.0-cm3 section of fresh brain taken from a consistent area of the dorsal cranial left frontal cortex was collected and frozen at −80°C until PCR analysis as detailed below. The remainder of each brain was placed in 10% neutral buffered formalin and fixed for a minimum of 7 days. Gross examination of each brain involved sectioning every 5.0 mm using a brain-trimming knife by the same anatomic pathologist (Miller). From each animal, identical sections of frontal cortex, basal ganglia, thalamus, hippocampus, medulla oblongata, and cerebellum were embedded in paraffin, sectioned at 5.0 µm, and stained with hematoxylin and eosin. Histological lesions were graded on a scale of 0–4 by a single pathologist (Miller). A score of 0 represented no inflammatory cells present, a score of 1 indicated rare lymphocytes and plasma cells present (<10 per 40× field), a score of 2 indicated small numbers of lymphocytes and plasma cells present (10–20 per 40× field), a score of 3 indicated moderate numbers of lymphocytes and plasma cells present (20–30 per 40× field), and a score of 4 indicated large numbers of lymphocytes and plasma cells present (>30 per 40× field).
For any areas with histological lesions, additional sections were evaluated by IHC for the presence of B. burgdorferi antigen, and two 5.0-µm shavings (5.0–10.0 mg) were cut and placed into a sterile 1.0-ml vial, h using a new scalpel blade for each block, for subsequent PCR analysis as detailed below. For IHC, 5 µm-thick sections were cut and placed on positively charged slides. j Briefly, after deparaffinization and dehydration, tissue sections were subjected to heat-mediated antigen retrieval with citrate buffer k for 10 min at 120°C followed by application of a universal blocking reagent l for 5 min at room temperature. A primary mouse monoclonal anti–B. burgdorferi antibody against the flagellar antigen m was used at a 1:1,000 dilution for 60 min. An alkaline phosphatase detection kit n was applied for 30 min, followed by a naphthol-based red dye o as a chromogen. Sections were then counterstained lightly with Gill hematoxylin and bluing before a cover slip was applied. Cultured whole B. burgdorferi spirochetes were used as a positive control.
For PCR, total nucleic acids were extracted from all samples (whole blood, CSF, fresh frozen brain, and shavings of paraffin-embedded, formalin-fixed tissues with histological lesions) with commercially available kits. p Polymerase chain reaction for the canine housekeeping genes histone 3.3 or glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was performed on all samples. 10 Negative controls with no DNA template were run in parallel with all PCR reactions. Additionally, mock nucleic acid extractions of sterile water were performed in parallel with all clinical cases and utilized as negative controls in all PCR reactions. Genomic DNA from a healthy dog was used as a PCR-negative control for all PCR reactions. DNA was amplified by real-time PCR by primers FLALS and FLARS previously designed to target a fragment of the flagellin gene (flaB). 5 Amplification was performed in a 20.0-µl final volume reaction containing 2× PCR mix, q 2.0 U of uracil–DNA glycosylase, r 0.2 µM of each primer, s and 5.0 µl or 2.0 µl of template DNA from CSF or brain tissue, respectively. After a single hot-start cycle at 95°C for 5 min, PCR cycled 70 times with the following parameters: 15 sec at 95°C, 30 sec at 55°C, and 30 sec at 72°C. t Borrelia burgdorferi strain B31 was used as a positive control for flaB PCR. Melting curve analysis was used to evaluate amplification specificity.
The results of the analyses of the individual dogs are summarized in Table 1. Exposure of dogs to ticks and tick attachment was successful in all 12 cases. By day 90, 11 of 12 exposed dogs were PCR positive for B. burgdorferi based on skin punch biopsy and were positive on ELISA. The second ELISA and serological test indicated that one of these dogs (case 14) did not have an immunoglobulin G response; however, this dog became clinically lame and anorexic requiring early termination, had severe histological joint lesions (data not shown), and was considered infected. Of the Lyme-infected dogs, 6 were co-infected with A. phagocytophilum. One tick-challenged dog (case 3) was negative on all Lyme tests and was considered not infected, though this dog did become infected with A. phagocytophilum. The unexposed, control dogs were confirmed negative by serological tests. All 14 dogs were also negative for D. immitis and E. canis infection on the serological test, though this is not indicated in Table 1.
Summary of infection status and neuropathological results in immunocompetent and immunosuppressed juvenile Beagles exposed to Borrelia burgdorferi–infected ticks.*
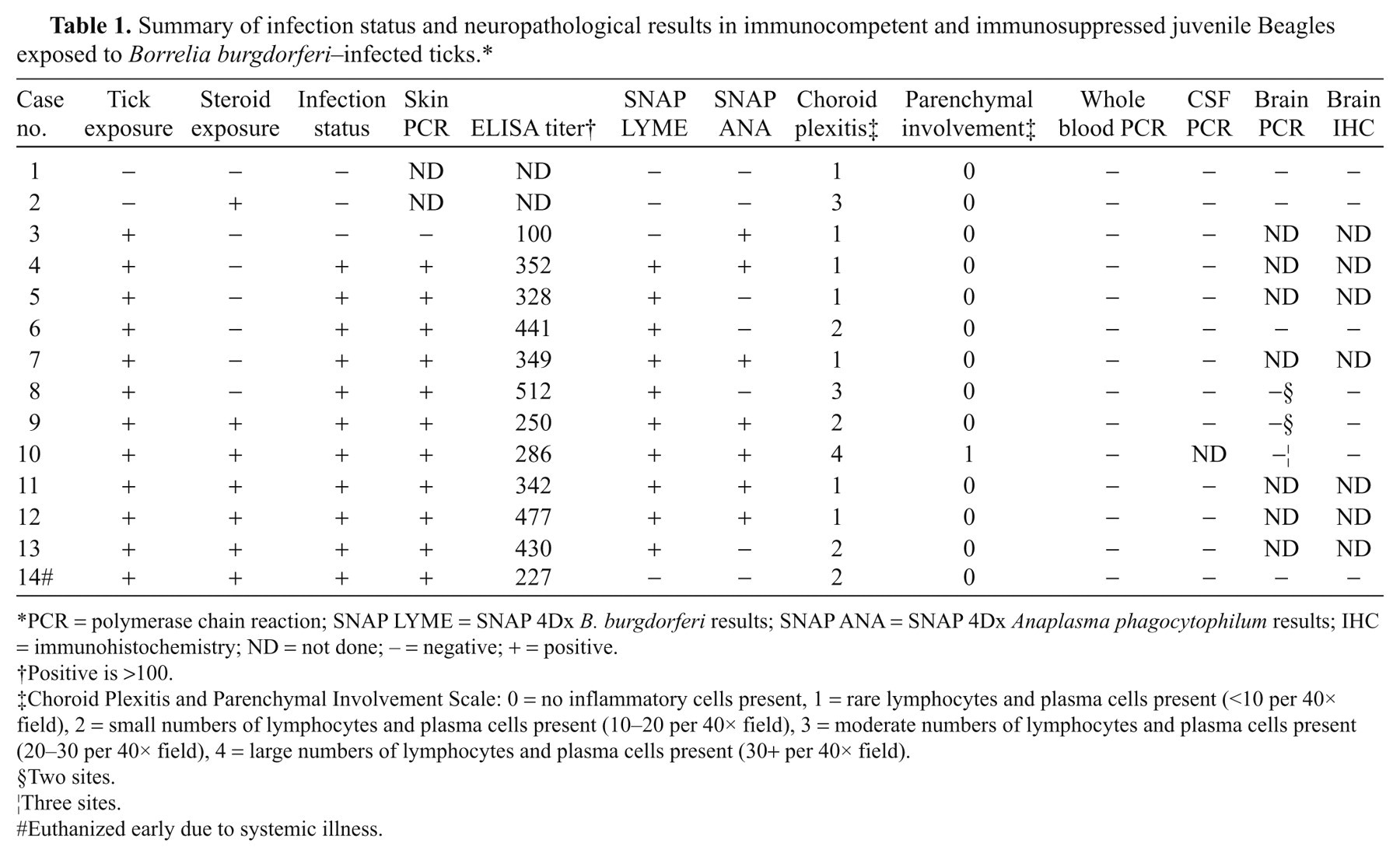
PCR = polymerase chain reaction; SNAP LYME = SNAP 4Dx B. burgdorferi results; SNAP ANA = SNAP 4Dx Anaplasma phagocytophilum results; IHC = immunohistochemistry; ND = not done; − = negative; + = positive.
Positive is >100.
Choroid Plexitis and Parenchymal Involvement Scale: 0 = no inflammatory cells present, 1 = rare lymphocytes and plasma cells present (<10 per 40× field), 2 = small numbers of lymphocytes and plasma cells present (10–20 per 40× field), 3 = moderate numbers of lymphocytes and plasma cells present (20–30 per 40× field), 4 = large numbers of lymphocytes and plasma cells present (30+ per 40× field).
Two sites.
Three sites.
Euthanized early due to systemic illness.
None of the infected or control dogs manifested neurological signs of disease. Four infected dogs showed lameness intermittently between day 5 and day 72 of infection and 2 dogs developed anorexia for 1 day or less in association with lameness and articular swelling. None of the dogs developed fever or weight loss, except for 1 dog euthanized early. This single infected dog and 1 immunosuppressed dog were euthanized at 164 DPI due to severe lameness, fever, and prolonged anorexia 11 days after initiation of corticosteroid exposure. All other dogs were euthanized as scheduled and did not exhibit clinical signs of infection.
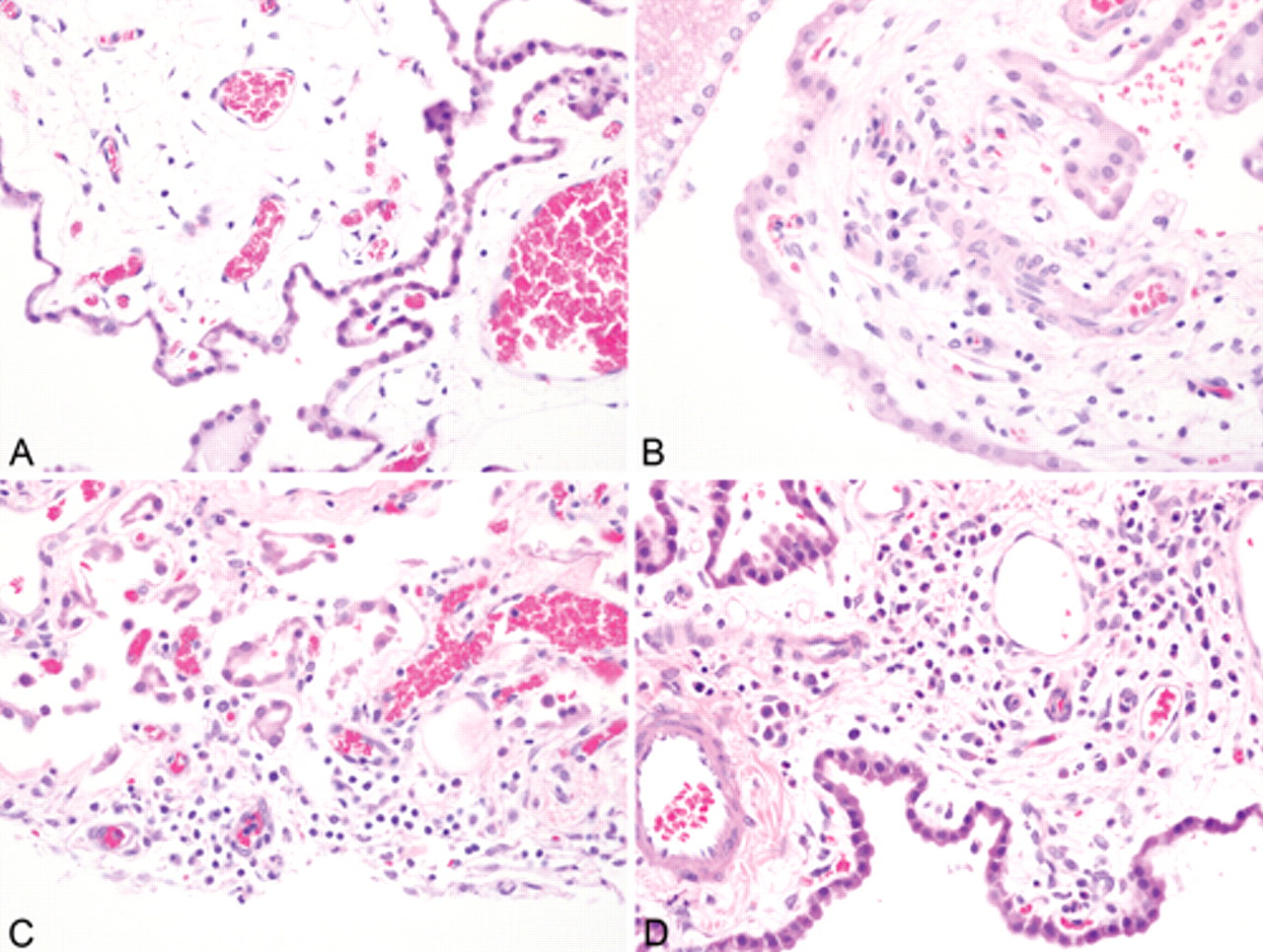
Six infected dogs (4 immunosuppressed, 2 immunocompetent) had evidence of inflammation (histological score >1) in the choroid plexus that ranged from scattered plasma cells to moderate numbers of lymphocytes, plasma cells, neutrophils, and macrophages in roughly equal proportions in the choroid plexus of both the lateral ventricle and the lateral aperture (Fig. 1). In 1 of the 4 immunosuppressed dogs (case 10), there was a single perivascular cuff of lymphocytes in the gray matter of the frontal cortex. Significant histological lesions in the brain were not identified in the 5 other infected dogs (2 immunocompromised, 3 immunocompetent), and the choroid plexus in these animals contained either no or extremely rare plasma cells. Lesions were not found on any other sections of brain, including the meninges. Not all immunosuppressed animals had histological lesions in the choroid plexus; however, those with choroid plexitis had an increased number of inflammatory cells compared to the animals that were not immunosuppressed. Of the 3 uninfected dogs, 1 control dog had a single follicular-like aggregation of lymphocytes bordered by rare plasma cells in the choroid plexus of the lateral ventricle. The dog that was exposed to ticks but did not become infected had minimal lesions graded at a histological score of 1, as did the other uninfected control dog. There were 11 brain sections with histological lesions from a total of 7 dogs (2 control dogs, 5 infected dogs) to which IHC was applied. All were negative for B. burgdorferi.
Nine dogs had sufficient CSF fluid for a complete CSF analysis and cytospin evaluation. One additional dog had sufficient CSF fluid for routine analysis but not cytospin evaluation. Two cases for which cytospins were performed had too much blood contamination for accurate cell counts. The other samples disclosed no abnormalities, and all had <5 cells/µl and total protein levels <16 µg/dl. The single sample for which a cytospin was not performed also had normal counts. Unfortunately, the only dog (case 10) in which CSF collection was unsuccessful was the case with the most severe histological lesions.
Polymerase chain reaction for B. burgdorferi in all dogs was negative on whole blood and fresh frozen brain, and was negative in the 13 CSF samples collected. Polymerase chain reaction on shavings from formalin-fixed, paraffin-embedded sections from areas of the choroid plexus with histological lesions, 11 sections from 7 dogs, also was negative in both control dogs and 5 infected dogs. Positive and negative controls produced expected results.
Clinical neuroborreliosis was not identified in any of the animals in the current experiment. Similar to a previous report, 23 CSF from experimentally infected dogs was uniformly normal, and Borrelia PCR testing on CSF was uniformly negative. Evaluation of CSF for anti-Borrelia antibodies would have been useful to evaluate for subclinical neuroborreliosis, but there was insufficient sample volume, and PCR was selected as it is considered the gold standard in human beings due to higher sensitivity. 1,19 Choroid plexitis (histological score >1) was observed in 7 dogs, which has not been reported previously in association with natural or experimental Borrelia infections; however, the significance of this finding is unclear, as one of the dogs with notable choroid plexitis was a negative control dog. In other species, small numbers of lymphocytes have been described in the choroid plexus. 9 Though a similar finding has not been reported in dogs, it is logical to assume that a small number of inflammatory cells would be expected within the choroid plexus of young dogs, and this concurs with the authors’ personal observations.
Choroid plexitis is a potential lesion that could be associated with the neurological signs previously reported in dogs with Lyme disease. 4,6,7 A similar pathogenesis has been proposed recently for leishmaniasis in dogs. 18 The function of the vascular choroid plexus is to produce CSF, and because of this, it is exposed to a relatively high burden of circulating organisms and antibodies. It is possible that for neurological disease to manifest, pre-existing damage to the choroid plexus and the blood–brain barrier must already exist to permit a breach in CNS defenses to organisms, immune complexes, and/or autoimmune antibodies. Biological variability in both the infective strain of B. burgdorferi and animal susceptibility will play a role in the manifestation of clinical disease and subclinical pathology. A pattern or relationship between co-infection status with A. phagocytophilum and choroid plexitis or other lesions was not found in the current study, and the single dog solely infected with A. phagocytophilum had minimal histological lesions.
In both dogs and human beings, the most severe systemic lesions described with experimental and natural infection include lymphoid hyperplasia, lameness, and peripheral neuritis, which often manifests more severely in close proximity to the tick attachment site. 6,20–22,24 As has been described by others, 6,24 lymphoid perineuritis in skin and around joints of the infected dogs was also identified in the current study (data not shown), which suggests the possibility of a retrograde spread of organisms to the CNS.
Synovial lesions in Lyme disease are presumed to occur because of a cyclical inflammatory response related to increasing numbers of spirochetes, which are destroyed, and then multiply repeatedly. 6 In these instances, the lymphoplasmacytic phase is presumed to be subclinical. Given the negative PCR results, a similar cyclical inflammatory response relating to waxing and waning organism burden in the CNS seems unlikely. Lyme nephritis studies in dogs have led investigators to conclude that renal involvement is likely attributable to immune complexes rather than direct infection, 7,8 and a similar immune-mediated pathogenesis is possible for the choroid plexitis identified in the dogs in the current study.
The complete etiopathogenesis of Lyme neuroborreliosis in human beings has yet to be elucidated. It is hypothesized that following tick attachment, spirochetes spread to the CNS either hematogenously or along peripheral nerves. 20 The mechanism by which Borrelia gain entrance to the CNS is unknown; but organisms have been detected in CSF in human beings by both PCR and culture. 20 In human beings, the most common manifestations of neuroborreliosis are T-cell lymphocytic and plasmacytic aseptic meningitis and a painful radiculitis. Rarely, parenchymal brain and spinal cord inflammation occur, and cranial nerve inflammation also has been reported. 19,20 Consistent with previous studies, meningoencephalitis was not identified on postmortem evaluation of the experimentally infected dogs reported herein. 6,7,24 It is also noteworthy that brainstem or leptomeningeal involvement was not detected as has been reported previously in other experimental studies. 6,23
The negative PCR findings on brain sections associated with choroid plexitis suggest that colonization by B. burgdorferi and persistent infection (i.e., direct cytotoxicity) is an unlikely mechanism of canine neuroborreliosis. Borrelia DNA was not detected in blood or CSF of infected dogs in the present study. This is in contrast to one other investigation, in which Borrelia DNA was identified by PCR in the meninges of 7 experimentally infected and immunosuppressed dogs. 6 The authors of the previous study reported lymphocytes and plasma cells in the dura mater of 3 dogs and a single small focus of encephalitis with lymphocytic infiltrates and gliosis within the caudal brain stem of 1 dog. However, the authors did not specify if PCR was performed directly on the brain lesions in affected dogs. Interestingly, the dogs from the previous report were younger than the dogs in the current study, and were also immunosuppressed during the active phase of infection, suggesting that immunocompetence may play a role in CNS involvement in dogs. However, based on the paucity of neuropathological lesions in the dogs of the current study, dogs seem less susceptible to the neurological manifestations of borreliosis that occur in human beings.
In conclusion, the lymphoplasmacytic choroid plexitis identified in dogs experimentally infected with B. burgdorferi warrants further investigation. Infection status with A. phagocytophilum did not appear to have any relationship with histological lesions in the brain. Any breach of the blood–brain barrier, such as exacerbation of preexisting choroid plexitis, could allow entry of immune complexes or autoimmune antibodies into the choroid plexus, and create more significant lesions that produce clinical signs. The lack of nucleic acids demonstrated on PCR, including lesions in the choroid plexus, provides evidence that B. burgdorferi does not have a direct role in the etiopathogenesis of canine CNS pathology.
Footnotes
Acknowledgements
The authors would like to thank Abbie Butler for help with immunohistochemistry, and Marsha Royston and her team of animal health technicians at Merial for their dedicated care of the research dogs.
a.
Covance Research Products, Cumberland, VA.
b.
Cornell University Animal Health Diagnostic Center, Ithaca, NY.
c.
Tick Encounter Resource Center, Kingston, RI.
d.
5mm skin punch biopsy, Miltex Instrument Co., Lake Success, NY.
e.
University of Georgia, Athens Veterinary Diagnostic Laboratory, Athens, GA.
f.
Canine SNAP 4Dx, IDEXX Laboratories, Westbrook, ME.
g.
BD, Franklin Lakes, NJ.
h.
Nalge Nunc International, Rochester, NY.
i.
Citra Buffer, BioGenex Laboratories Inc., San Ramon, CA.
j.
Hitachi 912, Roche Diagnostics Corp., Indianapolis, IN.
k.
Superfrost Plus Microscope Slides, VWR Scientific, Bridgeport, NJ.
l.
PowerBlock, BioGenex Laboratories Inc., San Ramon, CA.
m.
Clone B615M, Meridian Life Sciences, Saco, ME.
n.
BioGenex Laboratories Inc., San Ramon, CA.
o.
Vulcan Fast Red, Biocare Medical, Concord, CA.
p.
Qiagen MinElute Virus Spin Kit, Qiagen Inc., Valencia, CA.
q.
LightCycler 480 SYBR Green I Master, Roche Diagnostics Corp., Indianapolis, IN.
r.
LightCycler Uracil-DNA Glycosylase, Roche Diagnostics Corp., Indianapolis, IN.
s.
Integrated DNA Technologies Inc., Coralville, IA.
t.
Roche LightCycler, Roche Diagnostics Corp., Indianapolis, IN.
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Partial funding for this project was provided by Merial Limited, Athens, GA.