
Abstract
An indirect enzyme-linked immunosorbent assay (ELISA) was modified and validated to detect antibodies against Salmonella Abortusovis in naturally infected sheep. The ELISA was validated with 44 positive and 45 negative control serum samples. Compared with the immunoblot, the sensitivity and specificity of the assay were 98% and 100%, respectively. To follow antibody levels over time, samples from 12 infected ewes were collected at 1, 3, and 10 months after abortion. All animals showed antibody levels above the cutoff value throughout the observation period. One and 3 months after abortion, high antibody levels could be detected in all but one animal, whereas after 10 months, 9 animals had markedly lower but still positive antibody levels. The test characteristics and evidence for the persistence of detectable antibody levels in all infected animals for up to 10 months indicates that the ELISA can be used for herd surveillance testing.
Introduction
Since the lambing season of 2003–2004, several sheep flocks in the western part of Switzerland have been affected by abortion “storms” causing fetal losses in up to 70% of ewes. Bacterial examination from abortion material revealed an infection with Salmonella enterica subspecies enterica serovar Abortusovis, a disease that had not been reported since 1977 in Switzerland. 1,3
Salmonella Abortusovis is specifically adapted to sheep, and no zoonotic potential is known. After its introduction into naïve flocks, the abortion rate may reach approximately 60%, whereas in an endemic situation, abortions occur sporadically and only in young and recently introduced animals. 9 Transmission takes place through vaginal excretions and uptake of the bacteria from a contaminated environment, in which bacteria can survive for at least 6 months. 15 The agent can be isolated and cultured from fetal tissues, placenta, and vaginal discharges. 8
Serology may be useful for the diagnosis of S. Abortusovis infection. However, the most commonly used test is a serum agglutination assay, which detects immunoglobulin M (IgM) 11 present in the primary immune response of a first infection with S. Abortusovis. 5 Using serum agglutination, it has been shown that 3 weeks after experimental infection, agglutinin titers increase dramatically but revert to “noninfected” levels at 3 months after abortion. 8
Nevertheless, because ewes seem to be protected from further fetal losses for several years after abortion caused by S. Abortusovis, 9 a strong and long-lasting immune response seems to be induced by natural infection. The nature of this immune response is known to be antibody-mediated, and cell-mediated with gamma interferon production. 4 To investigate a long-lasting immune response, a diagnostic test able to detect IgG present in a secondary immune response would be indispensable. At present, no reliable screening assays are available.
The overall objective of the current study was, therefore, to modify and validate an enzyme-linked immunosorbent assay (ELISA) able to detect IgG antibodies directed against S. Abortusovis in ewes. The ELISA was used to detect antibodies during a 10-month follow-up after abortion in 12 naturally infected ewes and thus establish the basis for a seroepidemiological survey in Switzerland.
Materials and methods
Experimental design
Sera from sheep suffering from spontaneous S. Abortusovis abortion were analyzed by immunoblot, and a selection of them were taken as positive controls for ELISA establishment. Selection of sera serving as negative controls was based on the absence of abortion history and negative immunoblot results. To identify a representative S. Abortusovis strain for the ELISA, antigens of different S. Abortusovis strains were compared by immunoblot using 2 positive S. Abortusovis sera as well as 1 negative serum. To rule out nonspecific reactions between antibodies specific for Salmonella enterica subspecies diarizonae serovar 61:k:1,5,(7) (S. diarizonae O61; a Salmonella subspecies common in sheep in Switzerland) and antigens of S. Abortusovis, a serum that was shown to be positive for S. diarizonae O61 antibodies was analyzed with S. Abortusovis antigens by both immunoblot and ELISA. The ELISA developed in the current study was subsequently used for the follow-up study of the humoral immune response to S. Abortusovis, and results were compared with those of the serum agglutination assay commonly used in serologic diagnosis.
Sera
Samples were collected from 1 herd from the canton of the Valais (Switzerland) where during an outbreak “storm,” 56% of pregnant ewes had aborted within a period of 2 weeks. The etiology was confirmed by bacteriologic examination of abortion material. Blood samples were taken from the jugular vein of infected animals and centrifuged, and serum was stored at −20°C until use. Forty-one sera were collected 1 month after abortion, and 26 sera were collected 2 months later. Fourteen sera were collected at the end of the 10-month observation period.
Negative control serum samples were kindly provided by the Institute of Virology, Vetsuisse Faculty University of Zurich (Zurich, Switzerland) and had been collected in the region of Zurich from herds without abortion history in the past few years (based on information provided by sheep owners). Samples had previously been tested for antibodies against Brucella melitensis a and Chlamydophila abortus (formerly Chlamydia psittaci serotype 1) 2 with negative results. Salmonella diarizonae O61 serum was collected 2 weeks after abortion from an animal with a bacteriologically confirmed S. diarizonae O61 infection and stored at 20°C until use.
Bacterial strains
Salmonella Abortusovis strains were isolated from abortion material of several herds in western Switzerland. 1 Field strains from France were kindly provided by Dominique Gauthier, Laboratoire Départemental Vétérinaire et d'Hygiène Alimentaire (Gap, France), and field strains from Germany by Reinhard Sting, Chemisches und Veterinäruntersuchungsamt (Stuttgart, Germany). Salmonella diarizonae O61 was isolated from aborted lambs in western Switzerland. Bacteria for antigen production were grown in a brain–heart broth under constant shaking and aerobic conditions at 37°C to give an optical density (OD) of 0.9 at a wavelength of 600 nm.
Antigen preparation (lipopolysaccharides)
Salmonella Abortusovis and S. diarizonae O61 cells were harvested by centrifugation at 13,000 × g for 2–3 min, washed 3 times with phosphate buffered saline (PBS; 8.0 g/l NaCl, 2.9 g/l Na2HPO4 × 12 H2O, 0.2 g/l KCl, 0.2 g/l KH2PO4 at pH 7.2–7.4), and resuspended in 1 ml of extraction buffer (0.01 M Tris–HCl buffer, pH 6.8 with 0.5% sodium dodecyl sulfate) as previously described. 6 After boiling for 20 min, the samples were centrifuged at 14,000 × g for 20 min. The pellets were removed and the supernatants were treated with 0.5 mg/ml proteinase K b at 50°C for 60 min. After inactivation of the proteinase K at 100°C for 5 min, the extracted antigens (lipopolysaccharides) were dialyzed in coating buffer (0.01 M carbonate/bicarbonate) for 3 days and stored at 4°C.
Immunoblotting
Antigen preparations were separated by polyacrylamide gel electrophoresis in a 14% gel using the Laemmli buffer system. 10 The antigens were blotted on a polyvinylidene fluoride membrane with a pore size of 0.45 μm. c After blotting, the membrane was allowed to dry completely at room temperature (RT), cut into strips, and stored at 4°C. The strips were incubated for 1 hr at RT with the sheep serum samples diluted 1:200 in milk buffer (0.01 M Tris [pH 7.5], 0.1 M NaC1, 0.1% Tween 20, 1% skim dry milk), rinsed twice with PBS, and washed with the same solution for 10 min. Bound antibodies were detected by incubating for 1 hr at RT with an alkaline phosphatase-labeled monoclonal antibody specific for sheep and goat IgG. 4 After rinsing twice with PBS and washing in the same solution for 10 min, 4-nitro blue tetrazolium (NBT) plus 5-bromo-4-chloro-3-indolyl phosphate (BCIP) e was used to visualize bound antibodies according to supplier's recommendations.
Enzyme-linked immunosorbent assay
The indirect ELISA described previously 14 was slightly modified and validated to detect S. Abortusovis-specific antibodies in sheep using 4 types of microtiter plates: PolySorp™, MaxiSorp™, MediSorp™, and MultiSorp™. f To determine ideal antigen and serum dilution, a checker-board titration was performed. First, lipopolysaccharide (LPS) extracts were diluted 1:5 to 1:20,480 in coating buffer (0.01 M carbonate/bicarbonate), and the plates were coated with 100 μl/well. After incubating for 18 hr at 37°C, plates were washed 4 times with distilled water and dried for 20 min at 37°C.
In a second step, a negative serum, as well as a weak and a medium positive serum identified by immunoblot, were diluted 1:100–1:800 in PBST (8.0 g/l NaCl, 2.9 g/l Na2HPO4 × 12 H2O, 0.2 g/l KCl, 0.2 g/l KH2PO4, 0.05% Tween 20, pH 7.2–7.4). The ELISA plates were then incubated with 100 μl/well of the diluted sera for 1 hr at 37°C and washed 4 times with PBST. For detection of bound antibodies, plates were incubated for 1 hr at 37°C with a peroxidase-labeled monoclonal antibody specific for sheep and goat IgG d diluted 1:2,500 in PBST. After washing the plates 4 times with PBST, 100 μl of 3,3′,5,5′-tetramethylbenzidine sub-strate d was added to each well, plates were incubated for 20 min at RT, and reaction stopped with 100 μl H2SO4. Optical densities were measured at 450 nm.
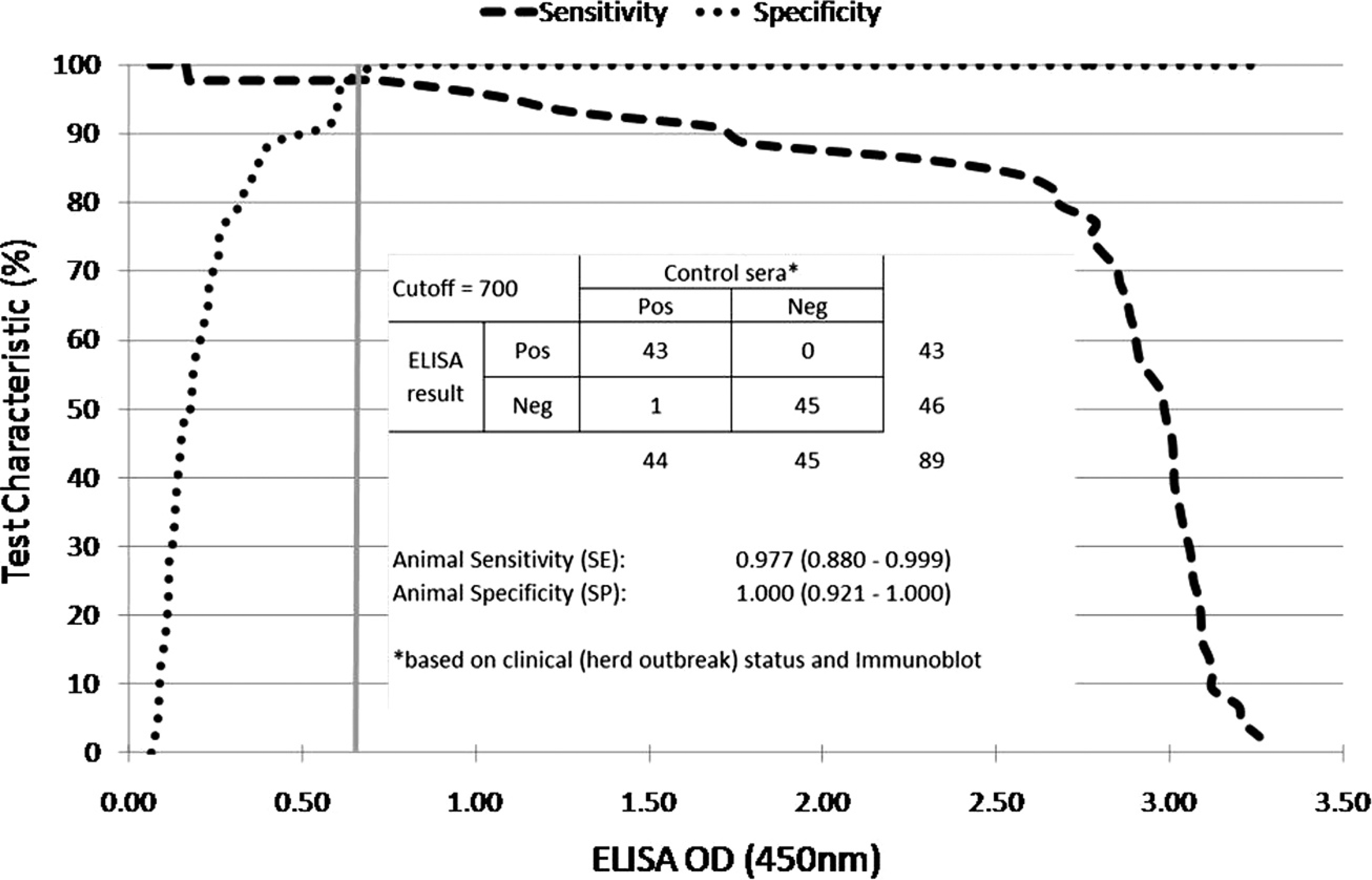
Validation of the enzyme-linked immunosorbent assay (ELISA) for Salmonella Abortusovis using receiver operating characteristic analysis for cutoff determination and cross-tabulation with comparison against positive and negative control sera for sensitivity and specificity determination. The vertical axis represents the respective animal-level assay sensitivity and specificity, and the vertical line indicates the respective cutoff value (0.7 optical density [OD] units) that yielded the frequencies in the 2 × 2 table.
To validate the ELISA, LPS and positive, as well as negative control serum samples (identified by immunoblotting), were diluted 1:600 and 1:100, respectively, according to the results of the checkerboard titration. An optimum cutoff value was determined using receiver operator characteristic (ROC) analysis and sensitivity and specificity determination resulted from cross-tabulation with comparison against positive and negative control sera (Fig. 1). For the longitudinal study, sheep field sera were double-tested, and results were defined as positive when at a positive control of at least 3.0 OD units, the mean OD value of both results was equal or above the test cutoff of 0.7 OD units chosen according to the results of the ROC analysis. To complete cross-reactivity testing with antibodies of S. diarizonae O61, 1 positive serum was analyzed by ELISA.
Agglutination assay
Agglutination assays were done as described previously. 14 Briefly, standard II agar plates were densely inoculated with S. Abortusovis and incubated for 24 hr at 37°C under aerobic conditions. The bacteria were killed by incubating the plates for 6 hr at 37°C in 96% ethanol, removed from the media using a spreading spatula, and centrifuged at 3,000 × g for 30 min. The pellet was resuspended in Ringer's solution and adjusted to a transmission of 5%. Sera were serially diluted from 1:10 to 1:1,280 in a 0.0025% wt/vol solution of safranin g in PBS (pH 7.4). Fifty microliters of bacterial suspension were added to 50 μl of diluted sera in round bottom 96-well plates and incubated for 24 hr at 37°C. Antibody titers of ≤1:320 were considered positive.
Results
Serum samples from ewes with confirmed S. Abortusovis infection were collected 1, 3, and 10 months after abortion. Immunoblotting revealed 68 positive sera of which 44 were used to validate the ELISA. All 45 negative control serum samples were negative for S. Abortusovis–specific antibodies on immunoblot. Using the same technique, the S. diarizonae O61 serum sample was confirmed to be positive for S. diarizonae O61-specific antibodies. To obtain a bacterial growth of S. Abortusovis reaching an OD of 0.9, a period of cultivation of 25 hr was required. In contrast to this, S. diarizonae O61 yielded this density under the same culture conditions already after an overnight cultivation.
Because there were no detectable differences between immunoblot patterns of S. Abortusovis strains from Switzerland, France, and Germany, the Swiss isolate that had caused the last outbreak in Switzerland was used. Testing different microtiter plates revealed the PolySorp plate to be the only one appropriate for S. Abortusovis LPS coating. This plate offered maximum resolution at the lowest background reactions. In contrast, the MaxiSorp and MediSorp plates showed higher background values culminating in equally high OD values for the negative serum and the positive sera using the MultiSorp plate. Titration experiments yielded best signal-to-noise ratios at an antigen dilution of 1:600 and a serum dilution of 1:100.
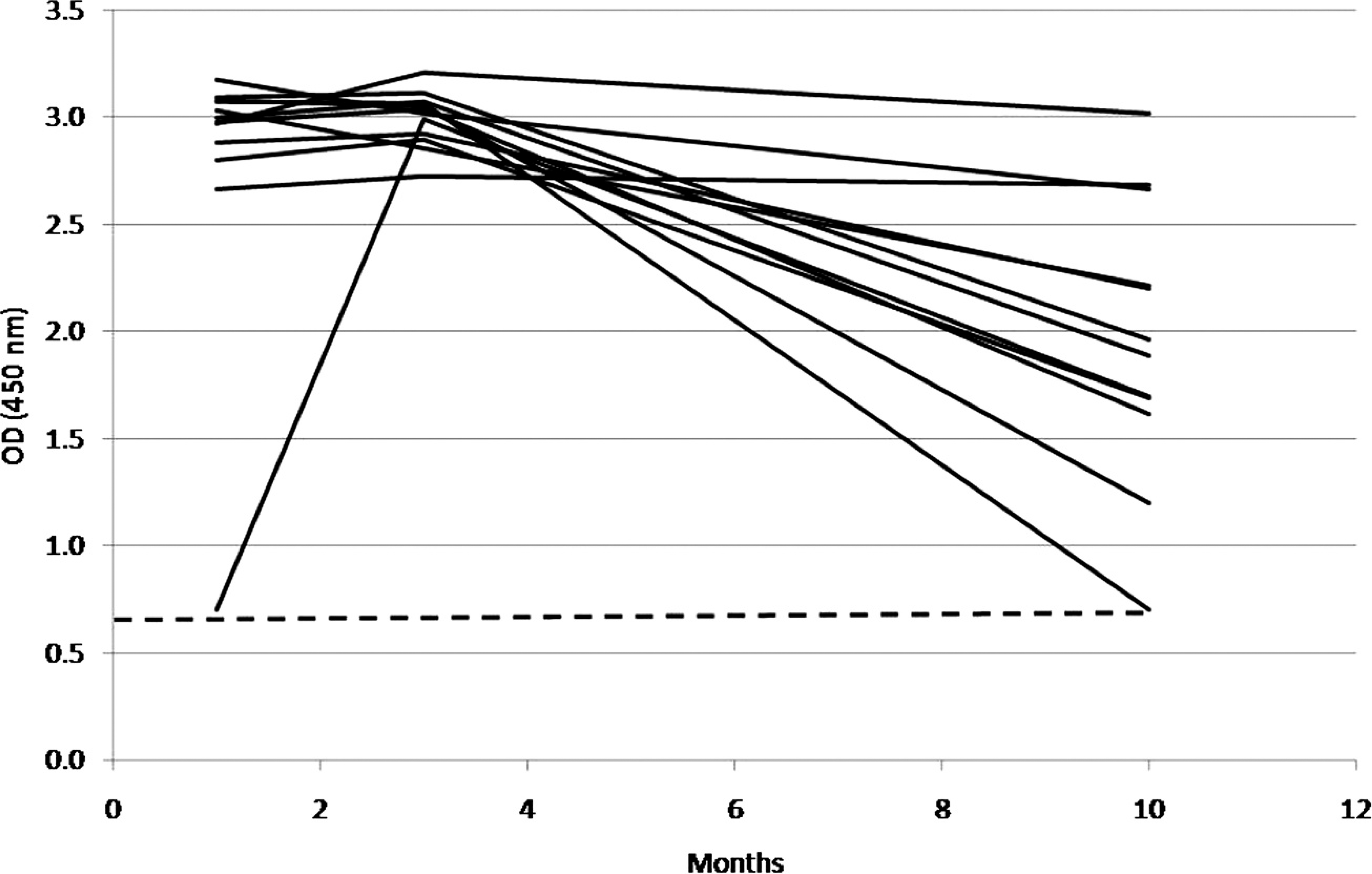
Detection of antibodies in 12 Swiss sheep 1, 3, and 10 months after abortion due to an infection with Salmonella Abortusovis. The dashed line indicates the selected enzyme-linked immunosorbent assay cutoff. OD = optical density.
The ROC analysis of the ELISA results revealed an optimum cutoff value at 0.7 OD units. When applying this cutoff value, the diagnostic sensitivity and specificity of the ELISA, using 45 negative sera and 44 positive sera previously classified by immunoblotting, were 97.7% (95% confidence interval [CI]: 88.0–99.9%) and 100.0% (95% CI: 92.1–100.0%), respectively (Fig. 1).
No reaction could be detected between S. Abortusovis antigens and S. diarizonae O61–specific antibodies on immunoblots. Furthermore, S. diarizonae O61 serum did not yield significantly higher OD values on ELISA than the negative reference serum and stayed well below the cutoff value. Therefore, cross-reactions with S. diarizonae O61 were considered unimportant.
Only 12 sheep from which samples were available for all 3 time points were used for the longitudinal study of the humoral immune response to S. Abortusovis. Using the ELISA, positive antibody levels were present throughout the 10-month observation period (Fig. 2). One month after abortion, serum samples of 11 of 12 animals revealed high antibody levels with OD values of approximately 3.0 (cutoff: 0.7), whereas the serum of 1 sheep was not as strongly positive with an OD value of 0.7. Two months later, this animal had also reached a high antibody level with an OD value of approximately 3.0, whereas the other animals maintained their high antibody levels. Ten months after abortion, antibody levels decreased markedly in 9 animals. In contrast to this, 3 animals had constantly high antibody levels throughout the entire observation period. In contrast, using the agglutination assay, 8 animals reached a positive agglutinating antibody titer of 1:320 1 month after abortion, and 4 animals reached that point 2 months later. Ten months after abortion, no positive agglutinating titer could be detected in any of the 12 animals.
Discussion
To detect S. Abortusovis antibodies after an abortion during a longer period of time, an ELISA that allows detection of IgG was modified and validated. Whereas previous publications dealing with humoral immune response to S. Abortusovis used the agglutination test, which mostly detects IgM present during the first few months after infection, the newly developed ELISA allowed confirmation of the presence of specific antibodies in ewes for at least 10 months after abortion. In contrast, no positive antibody titers could be detected at this time using the agglutination assay, showing a clearly higher sensitivity of the ELISA in samples collected months after infection.
Comparative analysis of LPS did not reveal any differences of immunoblot patterns between strains from Germany, France, and Switzerland, although genomic differences could be demonstrated between these isolates by pulsed-field gel electrophoresis. 1 Thus, it appears that LPS of S. Abortusovis are conserved antigens. Therefore, the ELISA established in the current project might not only be useful for seroepidemiological studies in Switzerland but also on a more global scale. Nevertheless, formal registration of this assay for testing individual animals would make a more comprehensive validation necessary.
Because the positive control sera originated from sheep with spontaneous disease the interval from infection and abortion to bleeding and thus the immune status varied between animals. Negative control sera were taken from animals without abortion history (based on the information of sheep owners) but with contact to the natural flora of sheep and possibly other pathogens. Use of these imperfect positive and negative control sera could yield poorer test characteristics than those that might be obtained by using ideal control sera sets. Because the test was intended for use in a survey in which several animals per flock would be sampled and tested, false-negative results would lead to an underestimation of the prevalence, especially at the flock level. Therefore, emphasis was put on high specificity, and an animal level test sensitivity of 98% was accepted as sufficient.
Infection with S. diarizonae O61 is the most frequently diagnosed Salmonella infection in sheep in Switzerland. Infections with other Salmonella strains are rare events and not associated with abortion (Raymond Miserez, personal communication, 2009). Therefore, it was important to rule out cross-reactions with this serotype. Although the newly developed ELISA is sensitive enough to allow detection of seropositive animals during a longer period of time, no significant cross-reactions between antibodies of S. diarizonae O61 and S. Abortusovis antigens could be observed either by immunoblot or by ELISA. Furthermore, as the animals in the negative reference cohort were not experimental sheep but originated from regular farms in Switzerland, they had contact with the bacterial flora of normal sheep and their habitat. Therefore, a significant cross-reaction problem would most likely have been detected in the validation process.
In 11 of 12 animals analyzed by ELISA, high antibody levels could already be observed about 1 month after abortion caused by S. Abortusovis infection. One animal with an initially low antibody level reached a level comparable with that of the other ewes by the time of the second sampling, suggesting that it had been infected later than its herdmates. Although 10 months after abortion all animals were still seropositive, antibody levels were markedly lower in most animals.
Therefore, antibody levels may not correlate with protective immunity because animals that aborted due to a S. Abortusovis infection are protected for several years 9 suggesting the presence of a long-lasting complementary protective cellular immune response. Indeed, 2–3 months after abortion the number of animals showing a reaction to an intradermal allergic test measuring the cellular immune response had been higher than the number of animals reacting positively in the agglutination test. 11 A protective booster effect that is induced by reactivation of a latent infection during gestation could also be responsible for this long-lasting immunity considering that pregnancy may influence antibody levels. A previous study 12 demonstrated that antibody levels induced by subcutaneous vaccination with a live attenuated strain of S. Abortusovis increased at the time of lambing but decreased between 2 pregnancies. Whether this holds true in natural infections has not been determined. Despite the long-lasting immunity, animals may remain asymptomatic carriers 7 and may infect naïve animals occasionally. Because the use of the ELISA test described in the current study is able to detect antibodies against S. Abortusovis for at least 10 months after abortion, this test is a reliable and sensitive tool, and therefore, could be applied for monitoring the spread of infections and assessing an epidemiological situation.
Acknowledgements
The authors acknowledge Patrick Boujon for excellent technical support, Joachim Frey for helpful discussions, and Lisa Harwood for the critical review. The authors thank Dominique Gauthier for providing strains from France, Monika Engels and Nicole Borel for providing negative control serum samples, and Christine Daves and the sheep owners for their supportive collaboration and for collecting samples. This study was kindly supported by the Federal Veterinary Office (project 01.07.16) and the Galli-Valerio foundation, Lausanne, Switzerland.
Footnotes
a.
IDEXX Switzerland AG, Liebefeld-Bern, Switzerland.
b.
Tritirachium album-Proteinase, AppliChem GmbH, Darmstadt, Germany.
c.
Millipore Corp., Billerica, MA.
d.
Sigma-Aldrich, St. Louis, MO.
e.
Roche Diagnostics, Rotkreuz, Switzerland.
f.
Nunc™; Thermo Fisher Scientific, Roskilde, Denmark.
g.
Merck, Darmstadt, Germany.