
Abstract
To date, the sensitivity of the interferon gamma (IFN-γ) enzyme-linked immunosorbent assay (ELISA) to detect Johne's disease (JD) has been poor, especially in the early stages of disease. To improve the sensitivity of IFN-γ detection in the early stages of infection, an alternate assay needs to be developed. The enzyme-linked immunospot (ELISPOT) assay is a highly sensitive technique for the detection of cytokines and has the potential to improve the diagnosis of JD. Of the variables examined, choice of capture antibody and the method by which the peripheral blood mononuclear cells were isolated significantly affected the ability to enumerate IFN-γ–secreting cells. The ELISPOT assay was as sensitive as or better than the IFN-γ ELISA at detecting ovine JD and could also detect disease at early time points postinoculation. The IFN-γ ELISPOT could distinguish infected from unexposed animals; however, neither the IFN-γ ELISA nor the ELISPOT assay could distinguish between sheep experimentally infected with Mycobacterium avium subspecies paratuberculosis and those exposed to the bacterium but diagnosed as uninfected at necropsy.
Keywords
Introduction
Johne's disease (JD) in ruminants is a chronic enteritis caused by infection with Mycobacterium avium subspecies paratuberculosis (MAP). The disease is predominantly spread by the fecal-oral route, with infection occurring in animals at a young age. In sheep, clinical signs of the disease (weight loss and the occasional case of diarrhea) develop from 2 to 5 years of age. One of the confounding issues in the diagnosis of ovine Johne's disease (OJD) is the infecting strain of MAP. There are 2 main strains of MAP: cattle (C) and sheep (S). Typically, these infect their named host species, but this is dependant on the geographical location. For example, in sheep, the S strain is more prevalent in Australia and New Zealand, while the C strain is more common in India. 34
Typically, it is difficult to diagnose JD in ruminants during the early stages of the infection, as it is only when the animals shed MAP into the environment and clinical signs start to develop that the sensitivity of the diagnostic tests improves. Current antemortem testing for JD in sheep includes agar gel immunodiffusion, antibody enzyme-linked immunosorbent assay (ELISA), fecal culture, and polymerase chain reaction (PCR). Fecal culture is the current gold standard for the identification of infected animals, although the S strain has more fastidious culture requirements than the C strain of MAP. 42 An interferon gamma (IFN-γ) ELISA has also been evaluated, but as with the other immunological diagnostic assays for MAP infection, this has poor sensitivity in the early stages of infection (Stewart DJ, Stiles PL, Whittington RJ, et al.: 2002, Validation of the interferon-gamma for diagnosis of ovine Johne's disease: sensitivity and specificity field trials. In: Seventh International Colloquium on Paratuberculosis. International Association for Para-tuberculosis, Bilbao, Spain). 37
The traditional method for detection of IFN-γ in ruminants has been to stimulate whole blood lymphocytes with MAP antigens, remove the plasma, and carry out an ELISA on the plasma. Many research groups typically use commercially available kits for this type of test, such as the Bovigam assay. a The detection of IFN-γ for the diagnosis of JD using this ELISA has had limited success, depending on the animal species and the stage of disease. In sheep, the Bovigam assay has low sensitivity, in the range of 50–75% (Stewart DJ, et al.: 2002, Validation of the interferon-gamma for diagnosis of ovine Johne's disease). 32 In cattle, IFN-γ ELISAs have a high rate of nonspecific reactions when detecting MAP infection in animals younger than 24 months, due to nonspecific IFN-γ production by natural killer cells, 27,28 although this phenomenon has not been observed in sheep.
Experimental inoculations of Merino sheep.
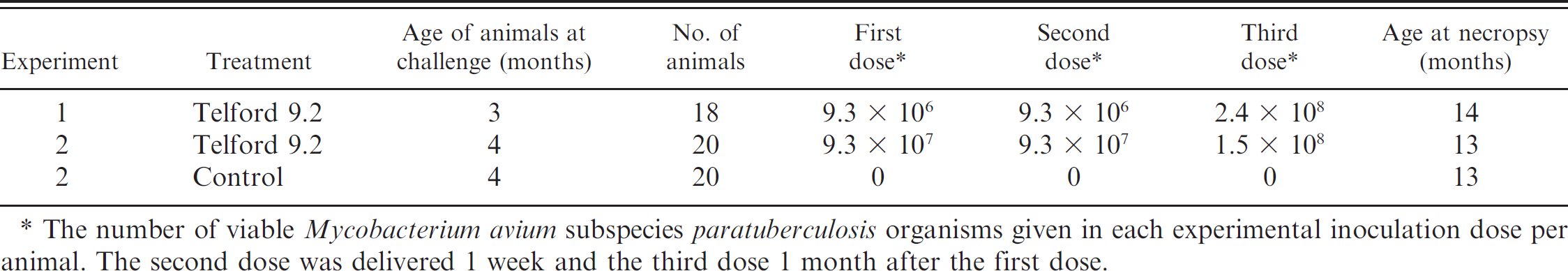
The number of viable Mycobacterium avium subspecies paratuberculosis organisms given in each experimental inoculation dose per animal. The second dose was delivered 1 week and the third dose 1 month after the first dose.
In recent years, tuberculosis (TB) detection in humans has improved through the use of IFN-γ assays based on the enzyme-linked immunospot (ELISPOT) technique rather than traditional approaches such as intradermal skin tests. 22 The ELI-SPOT assay detects the number of IFN-γ–producing cells rather than the total amount of IFN-γ as is measured by traditional cytokine ELISA assays. ELISPOT has been reported to be 10–200 times more sensitive in the detection of cytokines than the ELISA. 38 In addition, the ELISPOT has been developed as an alternative technology for diagnosing TB, as it has the advantage of being able to discriminate between different forms of TB, such as latent or active infections. 10 As these factors would also be important in the diagnosis of JD, an IFN-γ ELISPOT was developed for the detection of MAP infection in sheep. The ELISPOT was then compared with IFN-γ ELISA methods for the detection of MAP-infected sheep.
Methods
Animals
All animals used in the current study were used with the approval of the University of Sydney Animal Ethics Committee. The Merino sheep (Ovis aries) were maintained at pasture unless otherwise stated.
Vaccinated sheep. Two sheep were vaccinated with a single dose of a vaccine for MAP subcutaneously b (1 ml) to the anterior part of the neck as instructed by the manufacturer.
Naturally exposed or JD-free sheep. Animals were selected from farms in New South Wales, Australia, on which OJD had been previously identified or found to be free of the disease. The diseased animals were transported to the University of Sydney OJD quarantine facility and held for 2 weeks prior to necropsy. The animals were fed lucerne chaff and grain once daily with water supplied ad libitum. The OJD-free property was identified by previous monitoring in which 176 animals from the flock were tested by pooled fecal culture 40 and ELISA 15 with negative results. All individuals also tested negative for MAP in direct quantitative PCR examination of feces 20 ; 36 individuals in addition to those used in the present trial were culled over a 2-year period and found to be free of MAP infection by culture and histopathology at necropsy. Sheep from the OJD-free property were transferred to the university on the day of the necropsy.
Experimentally infected sheep. Mycobacterium avium subspecies paratuberculosis strain Telford 9.2, a pure clonal culture at passage level 5 (including its primary isolation from sheep feces), was reconstituted from lyophilized stock and inoculated into a BACTEC 12B c vial supplemented with egg yolk and mycobactin J (MJ). 42 After culture in BACTEC to achieve maximal growth, the vial was subcultured to 7H10+MJ 42 slopes and incubated for 4–6 weeks at 37°C. Telford 9.2 is an IS1311 Sstrain, IS900 restriction fragment length polymorphism type S1. 25,26
The slopes were harvested and single-cell suspensions prepared in phosphate buffered saline (PBS) with 0.1% v/v Tween 80. Enumeration was by visual counting in a Thoma-ruled counting chamber and by end-point titration in BACTEC medium, using the standard 3-tube most probable number (MPN) method. 31 The visual counts were used to prepare the inocula at 109 MAP organisms/ml of undiluted suspension. One-milliliter aliquots of this stock suspension were diluted in 10 ml of PBS for each of the animals to be infected, as shown in Table 1.
The remaining undiluted suspension (stock) was retained at 4°C to repeat the inoculation 1 week later. Another MPN was set up on the day of the second dose from the stock suspension stored at 4°C to confirm the viable dose after storage. IS900 and IS1311 PCR assays conducted on the stock solution were used to confirm that the infectious material was MAP S strain. A fresh suspension was prepared for the third inoculation dose, which was given to the animals 1 month after the first dose.
Blood-sampling regimes
For comparisons of antibodies and preparation methods for the ELISPOT, blood was taken as required from the 2 vaccinated sheep. Blood samples from OJD-free animals were taken either just prior to necropsy or, in 1 experiment, at 12 months of age and then again at 17 months of age, which was just prior to necropsy. Naturally infected sheep were blood sampled just prior to necropsy. Experimentally infected sheep were sampled every 2–4 months until necropsy.
Necropsy
All animals were euthanized using an intravenous injection of barbiturate into the jugular vein. The intestinal tract from the duodenum to the rectum was removed from the animal and placed in a clean tray. Between animals, the trays were cleaned with warm water and Trigene 2 d disinfectant. Samples were then taken from the following sections of the small intestine: terminal ileum and posterior, mid-distal, middle, mid-proximal, and anterior jejunum. The ileocecal lymph node and jejunal lymph nodes corresponding to the sections of jejunum sampled were also collected.
Histology
Tissue samples were placed in 10% buffered formalin for 48–72 hr, embedded in paraffin, and sectioned at 5 μm. Duplicate sections were stained with hematoxylin and eosin or the Ziehl–Neelsen stain. Sections were then graded using criteria previously outlined. 30 Any section graded with a score of 1 or higher was considered to be positive for OJD.
Isolation of MAP from feces and tissue
Samples were frozen at −80°C until they were processed. Fecal and tissue samples were processed for BACTEC culture as described previously. 42 Briefly, after decontamination, samples were inoculated into BACTEC vials and cultured for 12 weeks at 37°C. Culture-positive samples were then confirmed by PCR and restriction enzyme analysis as previously described. 41,42
Purification of peripheral white blood cells
Ficoll density gradient centrifugation. A 10-ml heparinized vacutainer of blood was centrifuged at 1,455 × g for 20 min. The buffy coat of white blood cells was removed, layered onto 2 ml of Ficoll-Paque e (δ = 1.077), and centrifuged at 754 × g for 30 min with no brake. The cloudy layer of white blood cells was removed and washed twice by centrifugation in 10 ml of PBS. The cells were resuspended in 5 ml of 10% fetal calf serum f made up in Roswell Park Memorial Institute (RPMI)-1640 f supplemented with L-glutamine (2 mmol), 2-mercaptoethanol (58 μmol), and penicillin/streptomycin f (100 μ/ml; complete media). The live cells were enumerated using Trypan blue and suspended at 2.5 × 106 viable cells/ml.
Crude white blood cell buffy coat preparations. A 10-ml heparinized vacutainer of blood was centrifuged at 1,455 × g for 20 min. The buffy coat of white blood cells was removed and washed twice by centrifugation in 10 ml of PBS. The cells were enumerated using Turk solution and resuspended in complete media as stated above at 2.5 × 106 viable cells/ml.
Red blood cell lysis. A 10-ml heparinized vacutainer of blood was centrifuged at 1,455 × g for 20 min. The buffy coat of white blood cells was removed and placed into 5 ml of red blood cell lysis buffer (NH4Cl 8.3 g, KHCO3 1 g, ethylenediamine tetra-acetic acid 3.72 g, and 10 mol of NaOH 0.4 ml in 1 liter of MQH20). After a 5-min incubation at room temperature, 5 ml of PBS was added to the buffy coat/lysis buffer mixture and centrifuged at 233 × g for 10 min. The cells were washed with PBS before enumeration using Trypan blue and resuspended in complete media as stated above at 2.5 × 106 viable cells/ml.
ELISPOT assay to detect IFN-γ
After optimization of the antibody concentrations and examination of various substrates, the following ELISPOT methodology was used in all assays unless otherwise stated. ELISPOT plates g were coated with 50 μl of 2 μg/ml of a mouse monoclonal antibovine IFN-γ, either IFN6.19 (immunoglobulin G2a clone 6.19) or IFN-CC330 (Serotec, clone CC330) as determined by the experiment, and incubated overnight at 4°C. The plates were then hand washed 6 times with PBS to remove excess antibody. Purified white blood cells at a concentration of 2.5 × 106 cells/ml were diluted to concentrations of 1.25 × 106 cells/ml and 6.25 × 105 cells/ml, with 100 μl of each dilution being placed into adjacent wells as required in all experiments. Fifty microliters of culture medium, either alone (unstimulated) or with MAP antigen h (316v, 30 μg/ml), and purified protein derivative from M. avium i (PPDA; 30 μg/ml) or pokeweed mitogen j (PWM; 10 μg/ml) was added to the wells. The plates were incubated for 18 to 24 hr at 37°C in air with 5% CO2.
At the end of the incubation, the plates were hand washed 6 times using PBS to remove the cells, and 50 μl of the secondary antibody, a biotin-conjugated mouse monoclonal antibovine IFN-γ (clone CC302 k ), was added at a concentration of 0.5 μg/ml diluted in PBS. The plates were then incubated at 37°C for 1 hr, washed in PBS, and 50 μl of alkaline phosphatase streptavidin 1 (1 μg/ml) diluted in PBS added. The plates were then incubated at 37°C for 1 hr, washed 5 times in PBS, and 100 μl of 5-bromo-4-chloro-3-indolyl phosphate (BCIP) substrate 1 was added and incubated for 1 hr at room temperature. The BCIP substrate was then removed and the plate washed 6 times in tap water, then allowed to dry and read in a KS ELISPOT reader. m
Results are presented as the number of antigen-specific IFN-γ–producing cells/105 white blood cells by taking the number of antigen-reactive cells and subtracting the number of unstimulated IFN-γ–producing cells/105 white blood cells. The test was considered to be valid if the number of PWM-specific IFN-γ–producing cells/105 white blood cells was greater than 15. Pokeweed mitogen stimulation was used for initial monoclonal capture antibody testing in the ELISPOT assay.
IFN-γ ELISA
Blood stimulations. Heparinized blood (0.5 ml) was placed into wells and stimulated with 0.5 ml of MAP 316v antigen at 20 μg/ml. The negative control consisted of blood with 0.5 ml of culture medium, while the positive control had 0.5 ml of media with PWM added (10 μg/ml). After 48 hr of culture at 37°C in air supplemented with 5% CO2, the plasma supernatant was collected and stored at −20°C.
Bovigam ELISA. a An analysis of IFN-γ was performed using the standard protocol outlined for the Bovigam ELISA kit. Results are presented as the optical density (OD) of the MAP antigen-stimulated sample – the OD of the negative control (medium-stimulated) sample.
In-house IFN-γ ELISA. ELISA plates n were coated with 50 μl of 1.5 μg/ml mouse monoclonal anti–IFN-γ antibody (IFN 6.19) and incubated overnight at 4°C. Frozen plasma supernatant samples were thawed and brought to room temperature. The plates were machine washed 5 times using wash buffer (PBS with 0.05% Tween 20) prior to adding 50 μl of PBS to appropriate wells followed by 50 μl of the plasma supernatant sample. The plates were incubated at room temperature for 1 hr and then washed 5 times using wash buffer. The secondary antibody (mouse monoclonal anti–IFN-γ antibody biotin-conjugated clone CC302; 50 μl) was added at a concentration of 0.5 μg/ml diluted in PBS. The plates were incubated at room temperature for 1 hr and then machine washed 5 times using wash buffer. Fifty μl of horseradish peroxidase–streptavidin 1 was added (0.01 μg/ml in PBS). The plates were incubated at room temperature for 1 hr and machine washed 5 times using wash buffer. TMB (3,3′,5,5′-tetramethylbenzidine) substrate o (100 μl) was added to each well as required, and the plate was incubated in the dark for 30 min. The reaction was then stopped with 100 μl of 2 mol sulphuric acid. The OD was measured in an ELISA reader p at 450 nm. Results are presented as the OD of the MAP antigen-stimulated sample – the OD of the medium-stimulated sample.
Results
Comparisons of antibodies and preparation methods for ELISPOT
After the initial antibody concentration optimization, the sensitivity of detecting spots in the IFN-γ ELISPOT assay was found to be affected by several parameters. These included the choice of capture antibody and the method of isolating the white blood cells. The ELISPOT reader was optimized for counting spots from the IFN-γ ELISPOT assay using the IFN-CC330 capture antibody. The same settings were used to compare the 2 capture antibodies (IFN-CC330 and IFN6.19) and the different white blood cell isolation methods.
In a direct comparison of the 2 capture antibodies, on average 6-fold more spots were resolved with IFN6.19 compared with IFN-CC330, measured across a range of test conditions including antigen-and mitogen-stimulated cells from various sheep. The difference was highly significant (paired t-test P < 0.001), with spots being more pronounced when using the IFN6.19 antibody, which enabled the ELISPOT reader to detect them more readily.
The initial optimization of the IFN-γ ELISPOT assay was completed on peripheral blood mononuclear cells (PBMC) isolated using Ficoll-Paque density gradient centrifugation. This was the standard method to which the red blood cell lysis buffer and crude white blood cell buffy coat isolation methods were compared. The effect of the cell isolation method was compared using both the IFN-CC330 and IFN6.19 capture antibodies, as shown in Table 2. Both red blood cell lysis and crude buffy coat methods produced significantly fewer spots than the Ficoll isolation method regardless of the capture antibodies used (Table 2). Wells containing buffy coat-prepared cells had a red-brown tinge in the center due to red blood cell contamination, with increased background color when higher concentrations of cells were used (Fig. 1). When cells were prepared by red blood cell lysis, there was no background color, but there were fewer spots across the range of cell dilutions tested (Fig. 1). The Ficoll isolation method was therefore used for subsequent experimental trials.
Effect of mononuclear blood cell isolation method on the outcome in the ovine interferon gamma (IFN-γ) enzyme-linked immunospot (ELISPOT) assay for 2 different anti-IFN-γ capture antibodies.
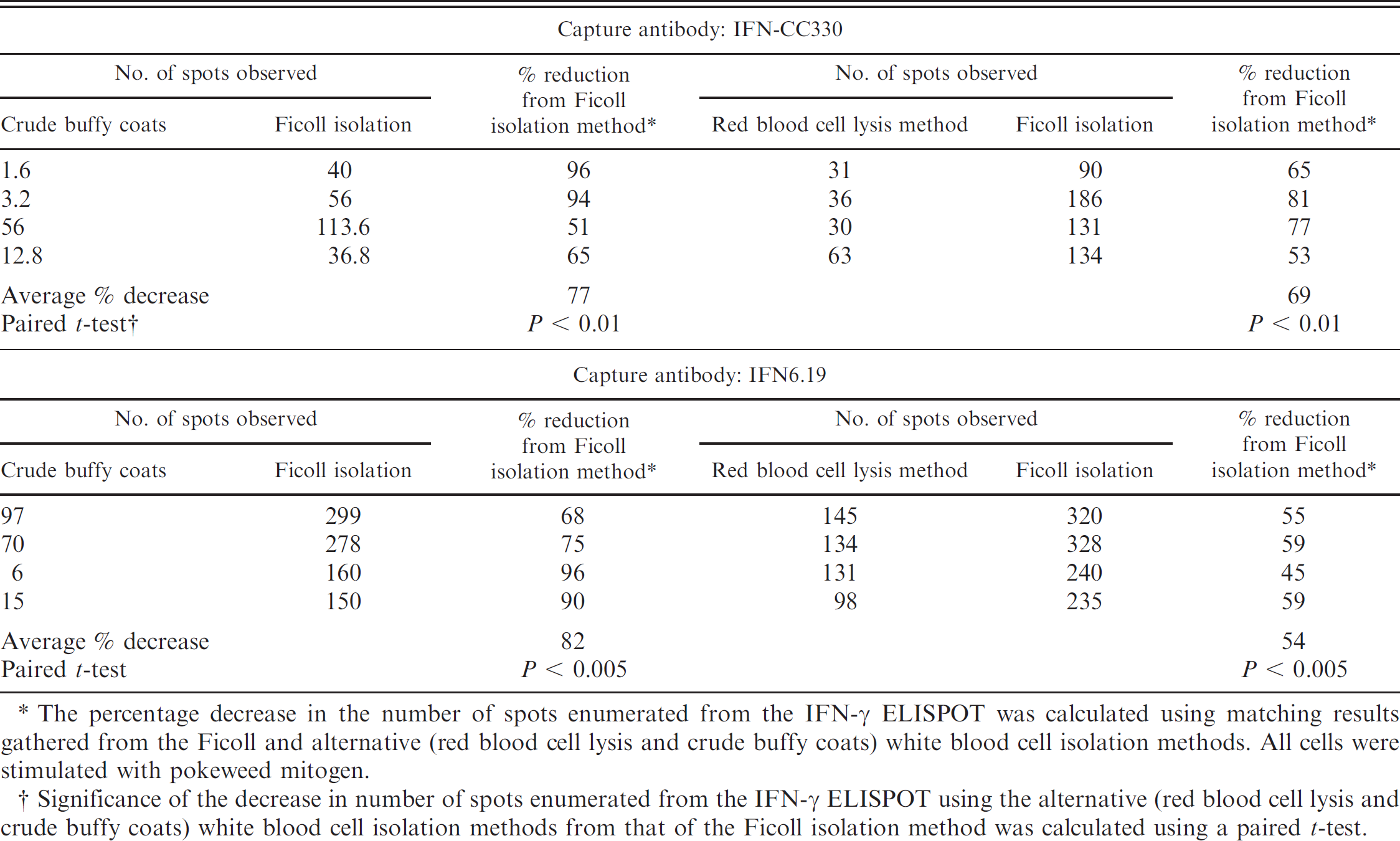
The percentage decrease in the number of spots enumerated from the IFN-γ ELISPOT was calculated using matching results gathered from the Ficoll and alternative (red blood cell lysis and crude buffy coats) white blood cell isolation methods. All cells were stimulated with pokeweed mitogen.
Significance of the decrease in number of spots enumerated from the IFN-γ ELISPOT using the alternative (red blood cell lysis and crude buffy coats) white blood cell isolation methods from that of the Ficoll isolation method was calculated using a paired t-test.
OJD-free animals
A group of 10 Merino ewes older than 3 years were selected from a farm shown to be free of MAP. The 10 animals tested negative for OJD by histological analysis and culture of MAP from their intestinal tissues. Although these animals were classified as OJD negative, 7 of the 10 animals had MAP-specific responses in the range of 1 to 10 reactive cells per 105 white blood cells (Fig. 2). However, 6 of the 10 animals had a stronger response to PPDA than to MAP antigen. Using these data, a preliminary cut point of 10 MAP reactive cells per 105 white blood cells was used to identify strong responders in the remainder of the study.
In another experiment, MAP antigen–specific responses in animals not exposed to MAP were compared over time. Figure 3 shows the change in MAP antigen–specific responses from a group of 20 uninfected animals from 12 to 17 months of age using the IFN-γ ELISPOT and ELISA. The sheep were later confirmed to be free of infection through tissue culture and histological analysis. While 11 of the animals consistently had low numbers of MAP antigen–specific cells over this time period, 9 animals had an increasing number; however, this increase was not significant (paired t-test). The in-house IFN-γ ELISA showed that most animals had an antigen-specific change of less than 0.1 OD. The average change for both assays over time was nonsignificant.
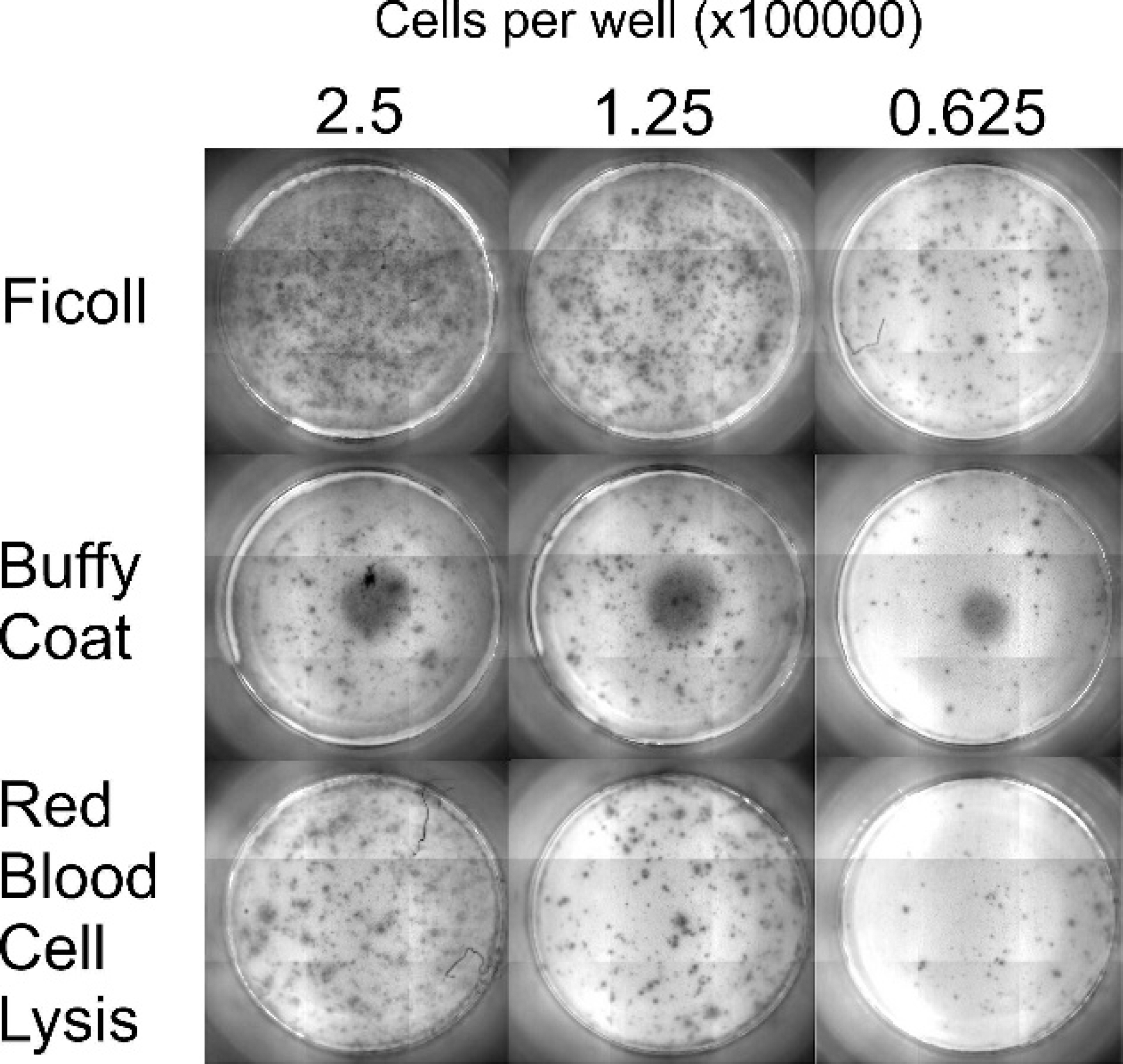
Images of the spots produced when using different white blood cell purification procedures. The capture antibody IFN-CC330 was used, while Ficoll-Paque density gradient centrifugation, isolation of the buffy coat via centrifugation, or lysis of the red blood cells from buffy coat preparations was used to purify the cells. All cells were stimulated with pokeweed mitogen.
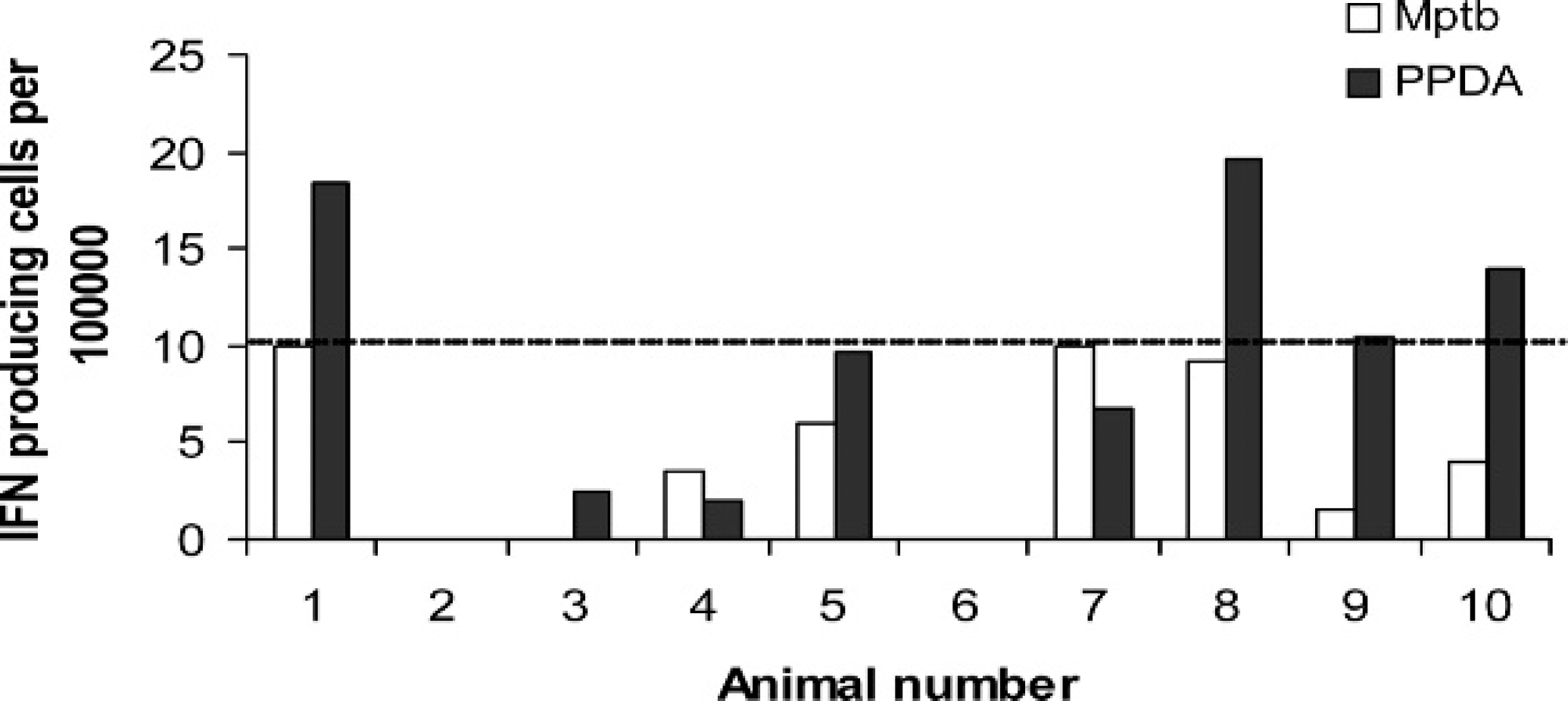
Interferon gamma (IFN-γ) enzyme-linked immunospot (ELISPOT) assay responses from ovine Johne's disease (OJD)-free sheep. Peripheral blood mononuclear cells (PBMC) were isolated from 10 sheep older than 3 years. The animals were found to be free of OJD by histological analysis and tissue culture of intestinal tissues. The assay used the IFN6.19 capture antibody and shows the antigen-specific responses to Mycobacterium avium subspecies paratuberculosis (MAP) and purified protein derivative from Mycobacterium avium (PPDA). The PBMC were isolated via Ficoll density gradient centrifugation. The dotted line indicates an arbitrary cutoff at which animals are considered to have a strong response to the MAP antigen.
Diagnosis of naturally infected animals
A group of animals from a farm found previously to have sheep with OJD were tested. The IFN-γ ELISPOT assay detected strong cytokine production (>10 MAP antigen–specific IFN-γ–producing cells per 105 white blood cells) from 8 of 10 MAP culture–positive animals. The 2 IFN-γ assays gave different results in 5 of the animals: sheep nos. 981, 986, 990, and 993 were strong responders in the ELISPOT only (Table 3). Sheep no. 981, which was MAP culture negative, also had a strong IFN-γ ELISPOT response (Table 3). The remaining 3 MAP tissue culture–negative animals had minimal IFN-γ responses in both the ELISPOT and Bovigam ELISA assays.
Longitudinal study of experimentally infected animals
Sheep experimentally inoculated, using a standardized infection model, with a clonal strain of MAP were monitored until the first signs of clinical disease developed in the flocks. MAP-specific IFN-γ–producing cells were detected by ELISPOT as early as 3 months postinoculation. The strength of this response varied over time and between the different experimental infection trials (Fig. 4A, 4C). Sampling of experimentally infected animals over time consistently showed that sheep inoculated with MAP had more antigen-specific IFN-γ–producing cells than uninoculated control animals (Fig. 4C).
Results from the MAP-specific in-house IFN-γ ELISA on the experimentally infected animals in trial 1 resembled the profile of the ELISPOT assay (Fig. 4A, 4B). In trial 2, the IFN-γ profiles of the 2 assays varied over time (Fig. 4C, 4D). The ELISA assay suggested progressively increasing total IFN-γ levels, while the ELISPOT assay detected higher responses in the infected sheep at necropsy. A drop in IFN-γ–producing cells was seen between 4 and 8 months postinoculation by ELISPOT that was not evident by ELISA. In trial 2, the ELISPOT assay was able to distinguish between tissue culture–positive and –negative animals at necropsy.
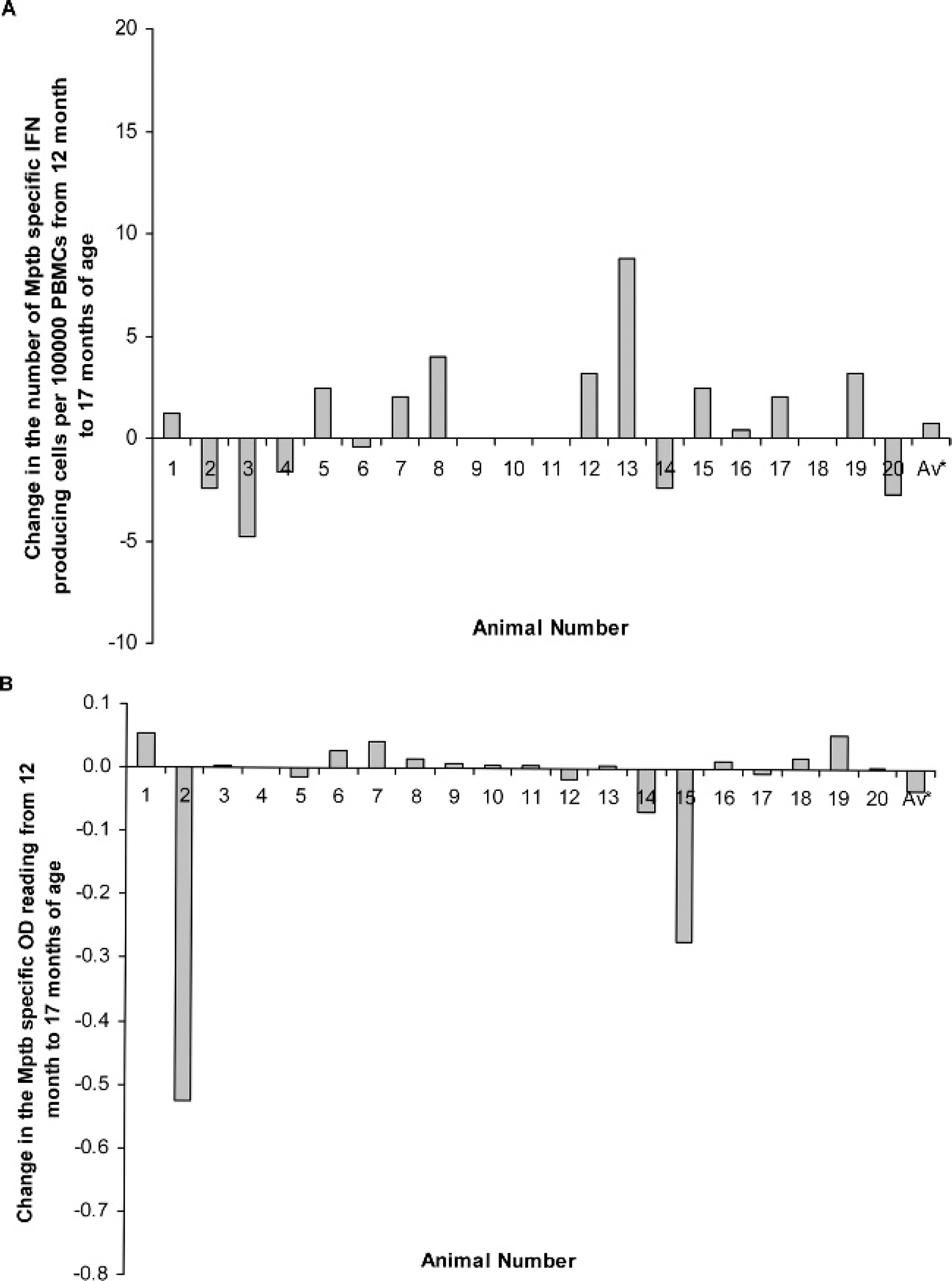
Change in Mycobacterium avium subspecies para-tuberculosis–specific interferon gamma (IFN-γ) responses in ovine Johne's disease–free sheep using an enzyme-linked immunospot (ELISPOT) assay and enzyme-linked immunosorbent assay (ELISA). Twenty sheep were monitored from 12 to 17 months.
Discussion
Results from the current study demonstrate for the first time the use of an IFN-γ ELISPOT assay applied to the diagnosis of JD in ruminants. Results from these experiments indicate that using the IFN-γ ELISPOT assay to detect MAP-infected sheep may provide a method as sensitive as or more sensitive than the traditional IFN-γ ELISA.
Interferon gamma (IFN-γ) results and infection status as shown by Mycobacterium avium subspecies paratuberculosis (MAP) tissue culture and histology from a group of animals from a farm known to be infected with ovine Johne's disease.*
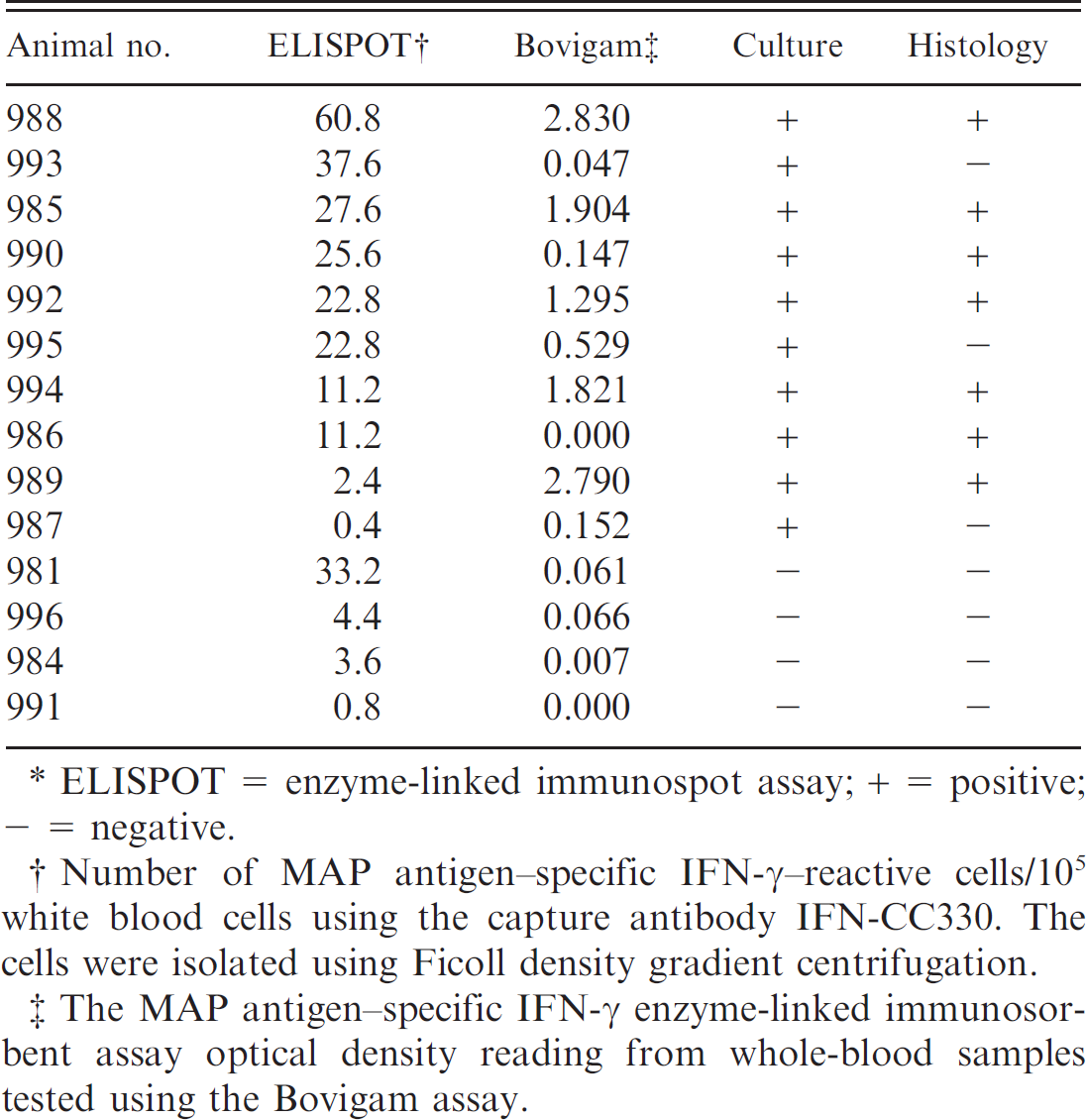
ELISPOT = enzyme-linked immunospot assay; + = positive; – = negative.
Number of MAP antigen–specific IFN-γ–reactive cells/105 white blood cells using the capture antibody IFN-CC330. The cells were isolated using Ficoll density gradient centrifugation.
The MAP antigen–specific IFN-γ enzyme-linked immunosorbent assay optical density reading from whole-blood samples tested using the Bovigam assay.
Important aspects that affect an ELISPOT assay include the affinity of the capture and detection antibodies, the activity of the enzyme conjugates, and the substrates used to generate the spots. 19 While there are a limited number of reagents available to veterinary researchers, 3 antibovine mouse monoclonal antibodies were found to be cross-reactive to ovine IFN-γ. Only 1 of the antibodies was commercially biotin conjugated (clone CC302), and this enabled evaluation of 2 different capture antibodies (IFN6.19 and IFN-CC330). As stated previously, 19 the darker the spots and the lighter the background, the more accurate the quantification. The IFN6.19 monoclonal antibody was selected as the optimal capture antibody as it produced superior spots and improved quantification, possibly due to this antibody's having a higher affinity for ovine IFN-γ.
The method by which the PBMC were prepared was also important. This method was assessed because possible future development of the assay will require high throughput and low cost. Ficoll-Paque density gradient centrifugation produced the most easily measured spots by the KS ELISPOT reader. Red blood cell lysis of buffy coat also gave well-defined spots, although fewer of them. Unfortunately, the easiest cell separation method examined, crude buffy coats, led to increased nonspecific background noise, and fewer spots were enumerated. A reduction in the number of spots when using the red blood cell lysis and buffy coat methods compared with Ficoll separation was not unexpected, as the latter removes most of the granulocytes, leading to the enrichment of the mononuclear cell population that produce IFN-γ. Neutrophils comprise 10–50%, eosinophils 0–10%, and basophils 0–3% of the total leucocyte population in sheep blood. 13 The IFN-γ–producing cells are most likely to be T cells, 2,36 so granulocytes dilute the number of IFN-γ–producing cells, resulting in fewer spots being recorded. While it may be possible to increase the number of cells added to increase the sensitivity from the red blood cell lysis and buffy coat methods, this may result in increased nonspecific background noise.
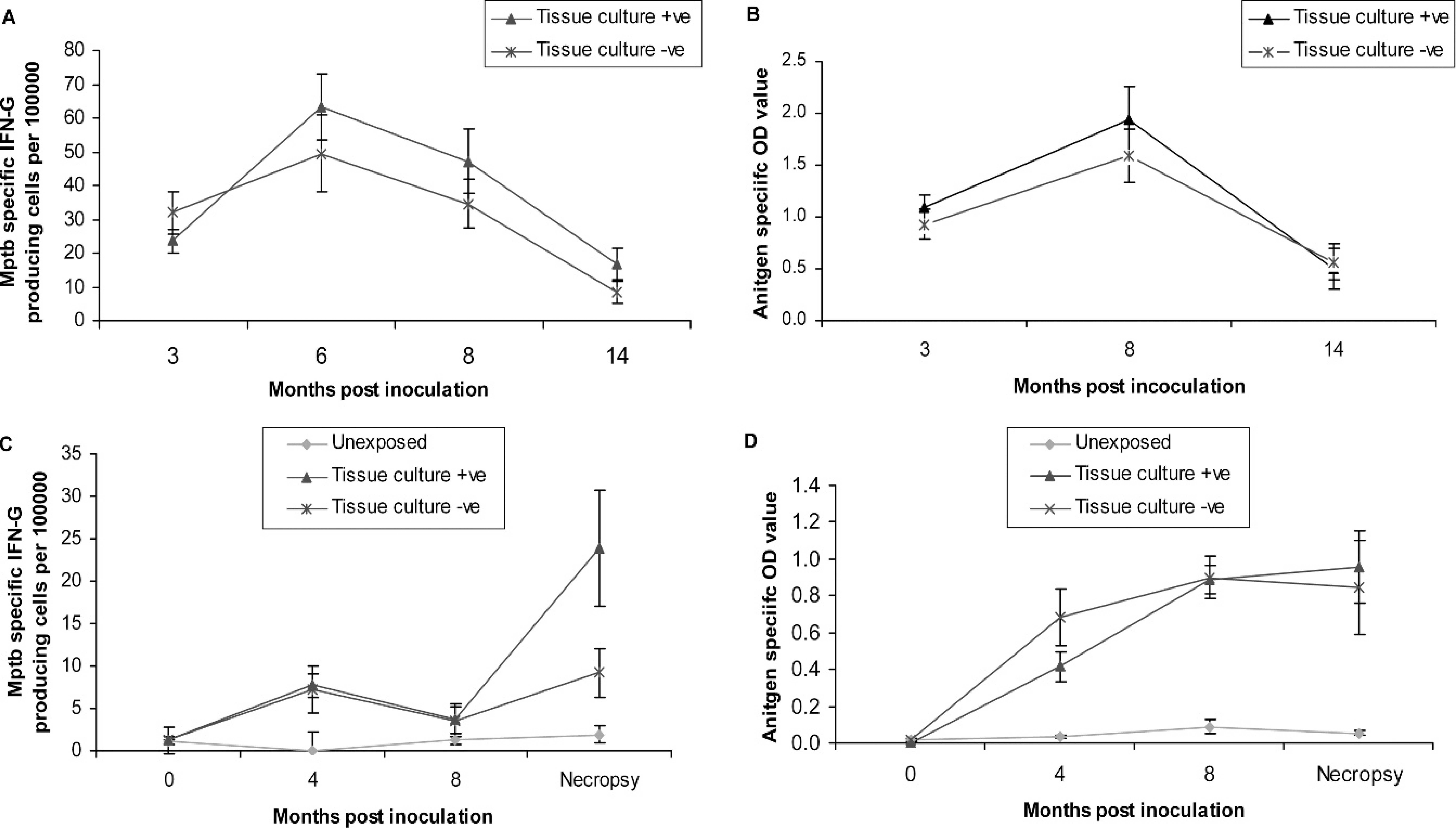
Mycobacterium avium subspecies paratuberculosis (MAP)–specific interferon gamma (IFN-γ) enzyme-linked immuno-spot (ELISPOT) assay responses from experimentally infected animals over time.
The number of MAP-specific IFN-γ–reactive cells in sheep infected with MAP appears to be very low, in the range of 10 to 100 IFN-γ–producing cells per 105 white blood cells. Differences between the 2 methods may be due to few cells producing large amounts of IFN-γ, and this could be an indication of disease status. Previous studies showed that as cattle and sheep move from subclinical to clinical JD, there is typically a drop in the cell-mediated immune response, 21,35 although in sheep, the IFN-γ response does not always decline. 3 As fewer cells may produce IFN-γ but at a higher level per cell, there may be a role for assays that measure total IFN-γ as well as the number of IFN-γ–producing cells. The cutoff value to determine uninfected from infected animals needs to be examined in more detail with larger numbers of animals.
The difference between the different types of IFN-γ assays (ELISA and ELISPOT) in identifying animals as being low or high IFN-γ responders (Table 3, Fig. 4) was not surprising and may aid further discrimination of diseased animals. The assays measure different outputs (the total amount of IFN-γ released into the plasma for the ELISA compared with the total number of IFN-γ–secreting cells measured in the ELISPOT). The different assay outputs could be used as part of an improved array of diagnostic assays to diagnose JD in the early stages of subclinical infection.
One sheep (no. 981; Table 3) that had been exposed to MAP naturally had a high number of MAP IFN-γ–producing cells, but this animal was negative by tissue culture and histology and, as such, apparently free of infection. This response can be explained from observations made in experimentally infected sheep. Animals that are inoculated with MAP may overcome the infection. 4,5,16,39 Exposed animals that had negative culture results from tissues (Fig. 4) have possibly eliminated the bacteria or are controlling the infection to a level lower than can be detected by microbiological culture of tissue. In trial 2, the number of MAP-specific IFN-γ–producing cells in animals with no detectable infection at 13 to 14 months after inoculation was higher than in the unexposed controls but lower than that of sheep with active infection at the time of necropsy. This indicates that in its current form, the IFN-γ ELISPOT assay provides evidence of exposure and may have the potential to discriminate active from past infection.
Unexposed sheep with no evidence of MAP infection by histology or tissue culture had low levels of MAP antigen–specific IFN-γ–producing cells. In general, these animals had a larger number of M. avium–specific cells per 105 white blood cells. This could be an indication of exposure to environmental mycobacteria and the poor specificity of the MAP antigen used. The use of crude antigens has been identified as a problem for the immunological diagnosis of JD in many studies. 1,7–9,23,24,43 There is a need for defined specific antigens that could enable extension of the assay to identify animals likely to be immune and those with a latent infection that will eventually succumb to disease. It may enable identification of sheep that go on to develop a specific lesion type. Research carried out on TB has shown that specific antigens such as region of difference 1 (RD1) including culture filtrate protein 10, early secreted target 6, and other proteins such as Rv2031c (also known as HspX, alpha crystalline, or the 16-kDa antigen) used in an IFN-γ ELISPOT tests can differentiate between active and latent TB patients and people who have been vaccinated with Bacillus Calmette-Guérin. 10,11,14,22 However, the use of a single RD1 antigen resulted in a high degree of specificity but low sensitivity. Improved results are seen when multiple RD1 antigens are used. 29 Numerous studies have been carried out to identify new antigens that may be used in the currently available and new MAP immunodiagnostic tests. 1,7,9,17,23,24 To date, these recently identified antigens have examined only humoral and not cell-mediated immune responses.
There are a number of documented technical concerns related to the use of ELISPOT assays. These include interoperator variation, especially with regard to the enumeration of the cytokine-secreting cells and the handling of blood samples. Interoperator variation was overcome in these studies by using a standardized protocol, assay-specific controls, a standard automated ELISPOT reader, and software parameters. 18,33 Other issues relating to blood sample handling apply to both the traditional IFN-γ ELISA and the ELISPOT assays and result in a reduction of the signal. Issues include the interval between blood collection and processing and the storage temperature of blood. The addition of stabilization reagents to the blood tube at sampling 12 or the addition of certain cytokines or antibodies at the time of stimulation (Jungersen G, Grell SN, Clemensen A, Howard CJ: 2005, Interleukin-12 potentiation of the interferon-gamma test rescues day old blood samples for diagnosis of paratuberculosis PPD specific cellular mediated immune responses. 8th International Colloquium on Paratuberculosis. Copenhagen, Denmark) 6 may overcome this problem. These refinements can be investigated in the ELISPOT assay for MAP.
The IFN-γ ELISPOT assay has the potential to improve the diagnosis of JD in animals, as shown in the current study. The assay can detect infected animals at a similar or better rate than the IFN-γ ELISA. This is the first successful application of an IFN-γ ELISPOT assay to detect naturally and experimentally MAP-infected ruminants. The ELI-SPOT assay could be an important tool as part of a diagnostic assay array to provide increased information on an animal's disease status. Investigations comparing the traditional IFN-γ ELISA with the ELISPOT assay and other methods of detecting cytokines are warranted.
Acknowledgements
The authors would like to thank Nicole Cater, Natalie Schiller, Anna Waldron, Craig Kristo, and Nobel Toribio for all their help with animal sampling and completing the numerous assays. The authors would also like to thank Meat and Livestock Australia for their generous funding and support for this work.
Footnotes
a.
Prionics AG, Schlieren-Zurich, Switzerland.
b.
Gudair®, Pfizer Australia Pty Ltd., West Ryde, New South Wales, Australia.
c.
BD, North Ryde, New South Wales, Australia.
d.
Medichem International, Sevenoaks, Kent, UK.
e.
GE Healthcare Bio-Sciences AB, Uppsala, Sweden.
f.
Gibco BRL, Grand Island, NY.
g.
Millipore Corp., Billerica, MA.
h.
Elizabeth Macarthur Agricultural Institute, Menangle, New South Wales, Australia.
i.
CSL Ltd., Victoria, Australia.
j.
Sigma-Aldrich, St. Louis, MO.
k.
AbD Serotec, Kidlington, Oxford, UK.
l.
Vector Laboratories Inc., Burlingame, CA.
m.
Carl Zeiss MicroImaging GmbH, Gottingen, Germany.
n.
Nunc, Roskilde, Demark.
o.
Pierce Biotechnology Inc., Rockford, IL.
p.
Multiscan Biochromatic Type 348, Labsystems, Helsinki, Finland.