
Abstract
The present report describes the analysis of 4 Deerpox virus isolates from California, Oregon, and Ontario, Canada. All 4 isolates were associated with cutaneous crusting lesions. Examination of selected samples by electron microscopy demonstrated that the viruses were morphologically similar to orthopoxviruses. Phylogenetic analysis of the A21 gene, which is found in all poxviruses, indicated that the 4 isolates form a lineage distinct from other members except for those belonging to the genus Cervidpoxvirus of the subfamily Chordopoxvirinae. Members of the Cervidpoxvirus lineage encode a set of genes not found in other poxviruses. These include homologs of genes encoding interleukin 1 receptor antagonists (IL-1Ra) and C-type lectin-like receptors (CTLR). In the current investigation, genes encoding homologs of IL-1Ra and CTLR were amplified from all the isolates and were found to be closely related to orthologs found in the Cervidpoxvirus genus, which further supports the inclusion of these isolates in the Cervidpoxvirus genus.
Introduction
Although 8 genera have been recognized within the Chordopoxvirinae subfamily of the Poxviridae family, sequence data from 2 closely related genomes (DPV-83 and DPV-84; 95% nucleotide sequence identity) of Deerpox virus (DPV) isolated from 2 Wyoming mule deer (Odocoileus hemionus hemionus) fawns suggested that they might be members of a distinct genus related to the Capripoxvirus (e.g., Sheeppox virus) and Suipoxvirus (e.g., Swinepox virus) genera. 1,8 In addition, the DPV genomes were found to encode several genes that may be involved in regulation of the immune system, which have not been reported in other members of the Poxviridae family. These genes are thought to modulate pathogenesis and are predicted to encode proteins similar to cellular endothelins, interleukin 1 receptor antagonists (IL-1Ra), C-type lectin-like receptors (CTLR), and the major histocompatibility complex-1 (MHC-1). One of the DPV strains (strain W-848-83 or DPV-83) encoded an open reading frame (ORF) similar to transforming growth factor β1(TGF-β1). Homologs of TGF-β1 are not found in other mammalian Chordopoxvirinae viruses, although distantly related homologs are encoded by avipoxviruses. Because of these distinctive features, the DPV isolates have been placed in a new genus, Cervidpoxvirus (CPV; by the International Congress for the Taxonomy of Viruses; E. Lefkowitz, pers. comm.).
Orthopoxvirus-like agents have been reported in reindeer, mule deer, and black-tailed deer. 2,6,8 In addition, Parapoxvirus, another genus of Chordopoxvirinae, has also been reported to infect a variety of cervids. 3,4,6,8 However, parapoxviruses cause skin lesions in a variety of animals (including sheep, goats, cattle, rodents, and pinnipeds) in addition to cervids. 3–6,8 The present report describes the characterization of 4 poxvirus isolates from diseased cervids located in California (DPV-CA), Oregon (DPV-OR1 and DPV-OR2), and Ontario, Canada (DPV-RD). In order to determine the etiology of these diseases, samples were propagated in cultured cells, then a gene conserved in all poxviruses (A21) and 2 genes specific to DPV (IL-1Ra and CTLR) were amplified and sequenced. From this data, it was determined that all the viruses contain homologs of IL-1Ra and CTLR and belong to the Cervidpoxvirus lineage.
Material and methods
Virus isolates
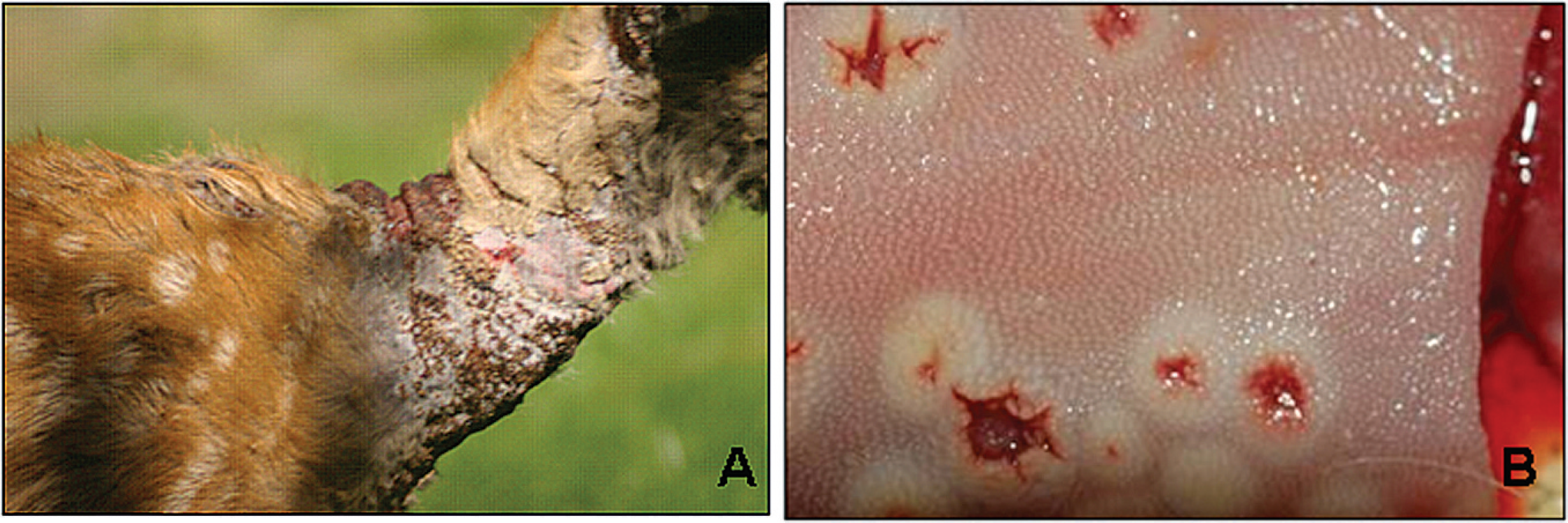
DPV-CA was from a deer rehabilitation facility north of San Francisco in Sonoma County, California. During the investigation of a recurring skin disease at the facility in 1994, isolates were recovered from biopsy material and scabs from healing cutaneous lesions on the neck of black-tailed deer (Odocoileus hemionus columbianus) fawns 2–5 months of age (Fig. 1A). The first Oregon isolate (DPV-OR1) was from skin collected during the necropsy of a wild mule deer (Odocoileus hemionus) fawn from Union county in northeastern Oregon in August 2005. This fawn had cutaneous lesions of the face and neck. The second Oregon isolate (DPV-OR2) was from the carcass of a male yearling black-tailed deer found near McMinnville in northwestern Oregon in July 2008. This animal was necropsied at the Oregon State University Veterinary Diagnostic Laboratory (OSUVDL; Corvallis, OR) and was found to have cutaneous lesions similar to those seen on the California fawns as well as oral lesions (Fig. 1B). The Canadian isolate (DPV-RD) was obtained in September 2008 using crusts and biopsy material from a 5-month-old female reindeer (Rangifer tarandus tarandus) housed at the Toronto Zoo (Ontario, Canada). Lesions on this fawn were located around the lips, muzzle, and eyelids. Diseased animals with similar symptoms have been observed periodically in the young stock of this captive Canadian herd since 1980, when the condition was first recognized and linked to infection with an orthopox-like agent. 2
Cell culture and virus isolation
Primary cultures of black-tail deer cells were derived from testicular tissue of a 7–9-month-old animal. The testes were aseptically removed, trypsinized, and cultured in minimal essential medium (MEM) + 10% fetal bovine serum (FBS). They were cryopreserved in 1999 after 1–3 passages. Vero (African green monkey kidney epithelial) cells were maintained at 37°C in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% FBS, a penicillin (100 U/ml), and streptomycin b (100 μg/ml) with 5% CO2 in a humidified incubator.
Viral isolates were isolated from skin tissue and scabs collected via biopsy or necropsy. The tissues were homogenized in 10% DMEM supplemented with penicillin (200 U/ml), and streptomycin b (200 μg/ml), and centrifuged at 700 × g for 10 min. The supernatants were inoculated onto primary black-tail deer cells. Positive isolation was confirmed by the observation of cytopathic effects (CPE) in the cultured cells and with electron microscopic examination of negatively stained specimens.
The virions were propagated in Vero cells as described above. Confluent cell monolayers were infected with plaque-purified virus at a multiplicity of infection of 0.1. Virus was harvested when more than 90% of the cells showed CPE. The infected flasks were subjected to 2 freeze–thaw cycles. The medium harvested from infected Vero cells was cleared of cells and cell debris by centrifugation at 9,000 × g for 30 min. The virions were then centrifuged in an ultracentrifuge c at 20,000 × g for 1 hr. The pellets were suspended in phosphate buffered saline (PBS) and frozen at −70°C until required.
Electron microscopy of virions
Virus particles purified by ultracentrifugation of infected cell suspensions, as described above, were adsorbed to Formvar d -coated carbon-stabilized copper grids by floating grids on ∼20 μl drops of the sample spotted on parafilm. The grids were then blotted dry with Whatman filter paper e and immediately floated on ∼20 μl drops of 2% phosphotungstic acid (PTA; pH 6.9) in water for 30 sec. Excess PTA was removed by side-blotting, and the grids were allowed to air dry. Images were obtained with an electron microscope. f
Purification of viral DNA
Virus particles purified, as described above, were extracted twice with 1:1 phenol:chloroform, a and once with 25:24:1 phenol:chloroform:isoamyl alcohol. a DNA was precipitated using 2 volumes of 100% ethanol and 1/10 of 3 M sodium acetate, washed once in 70% ethanol, and resuspended in TE buffer (10 mmol Tris-HCl, 1 mmol ethylenediamine tetra-acetic acid [EDTA], pH 8.0).
Primers and polymerase chain reaction amplification
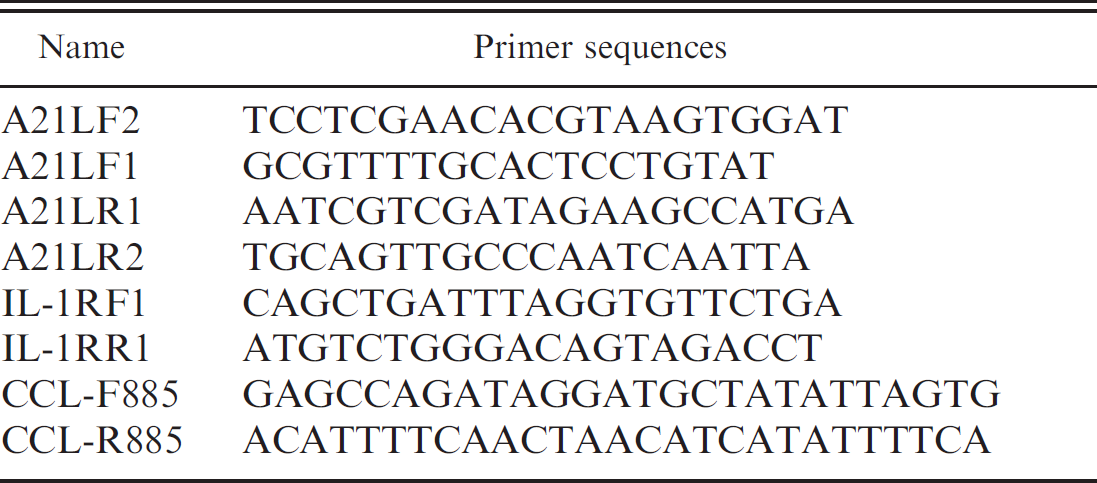
The oligonucleotide primer sets used for polymerase chain reaction (PCR) are listed in Table 1. Selection of the sequences used to design these primers for PCR was based on the known genomic information of mule deer pox isolates, W83 and W84 1,8 (GenBank accession nos. AY689436 and AY689437, respectively). They were selected by the Primer 3 program (http://frodo.wi.mit.edu/). Each PCR reaction contained 1 × buffer, g 1 × en-hancer, a 0.4 mmol each deoxyribonucleotide triphosphate (dNTP), a 1 mmol MgS04, a 1.25 U Econo Taq DNA polymerase, g 0.4 nmol of each primer, and 0.5 μg total DNA. The PCR was conducted at 94°C for 2 min and 30 cycles of 94°C for 30 sec, 50°C for 45 sec, and 72°C for 75 sec, finishing with a delay step of 72°C for 5 min. The resulting products were separated on a 1% agarose gel in 1 × TAE buffer (40 mM Tris-acetate, 1 mM EDTA).
. Primers used to amplify gene A21, and the interleukin 1 receptor (IL-1R) and C-type lectin (CCL) homologs.
DNA sequencing and analysis
Sequences were determined by direct sequencing of PCR products purified with a commercial PCR clean-up kit. a Further data were generated using primers designed from the initial sequence. All sequencing was carried out by the Center for Genome Research and Biocomputing (CGRB) at Oregon State University (Corvallis, OR). h Phylogenetic analysis of the concatenated amino acid sequences of A21, IL-1R, and CTL gene was carried out using MacVector 9.5.2 i software with the following settings: neighbor joining; best tree; tie breaking = systematic distance; gaps distributed proportionally; bootstrap (1,000 replicates).
Results
Growth of virus in cell culture
When the 4 isolates were grown in primary black-tail deer cells, CPE was observed at 3–5 days postinfection. The CPE was characterized by enlarged cytoplasm containing many inclusion bodies and cell detachment at 7–10 days postinfection. All isolates were subcultured in Vero cells in which they produced very small plaques. Isolates DPV-CA and DPV-OR1 yielded titers of 1 × 105 plaque-forming units (PFU)/ml, and isolates DPV-OR2 and DPV-RD produced slightly higher titers of 2–3 × 105 50% tissue culture infective dose (TCID50)/ml.
Electron microscopy
Virions of both DPV-CA and DPV-OR1 had an ovoid morphology at approximately 200 nm × 230 nm, which is similar to orthopoxviruses (Fig. 2). Similar results were obtained for DPV-OR2 and DPV-RD.
Molecular phylogeny
In order to determine the relationship of these isolates to other members of the Poxviridae family, gene A21 was examined. Gene A21 is present in all poxvirus genomes. It is comprised of 117 amino acids in the prototypic Vaccinia virus, forms a component of the entry fusion complex, and is an essential protein for cell-to-cell spread. 7 Therefore, orthologs of gene A21 from the 4 isolates were PCR amplified, sequenced, and analyzed. From this data, all the viruses were found to form a distinct lineage most closely related to the CPV genus (Fig. 3). In addition, all isolates were found to be closely related to one another and to the published DPV isolates. The A21 sequences of DPV-83 and DPV-84 are identical, as are those of DPV-CA and DPV-OR2. The A21 sequence of the most divergent isolate was DPV-OR1, and displayed 97.4% amino acid identity to DPV-CA.
To investigate the relationships among the isolates, 2 CPV-specific genes were characterized. The genes included homologs of CTLR and IL-1Ra. In members of the Poxviridae family, these genes have only been reported in CPV strains and appear to be a defining feature of this genus. 1 In these analyses, the viral genes were compared with homologous proteins from both human (Homo sapiens) and cattle (Bos taurus). The CTLR sequences formed a group closely related to the previously described sequence from DPV, showing more than 80% amino acid sequence identity (Fig. 3). The DPV-OR1 isolate appeared to form a well-supported distinct lineage, as did DPV-RD and the 2 Wyoming isolates (DPV-83/84). Another lineage appeared to be shared by DPV-CA and DPV-OR2. In contrast, the DPV proteins showed approximately 48% and 31% identity to the homologs from B. taurus and H. sapiens, respectively. The IL-1Ra sequences (Fig. 3) showed a phylogenetic tree similar to the CTLR sequences, though the sequences were somewhat more conserved than CTLR, with greater than 86% amino acid sequence identity. The DPV CTLR sequences show about 40% identity to the B. taurus and H. sapiens homologs. The differentiation of these isolates was not well supported, based on the IL-1Ra sequences.
The DPV-CA and DV-OR2 isolates are very closely related. In addition to their identical A21 amino acid sequences, their IL-1Ra sequences are also identical, and their CTLR sequences only differ by 1 amino acid. Overall, the DPV-CA and DPV-OR2 isolates show a 2-nucleotide (nt) difference in the 1,467 nt encoding these 3 genes. Because of this close relationship, they have been combined in Figure 3 to simplify the diagram.
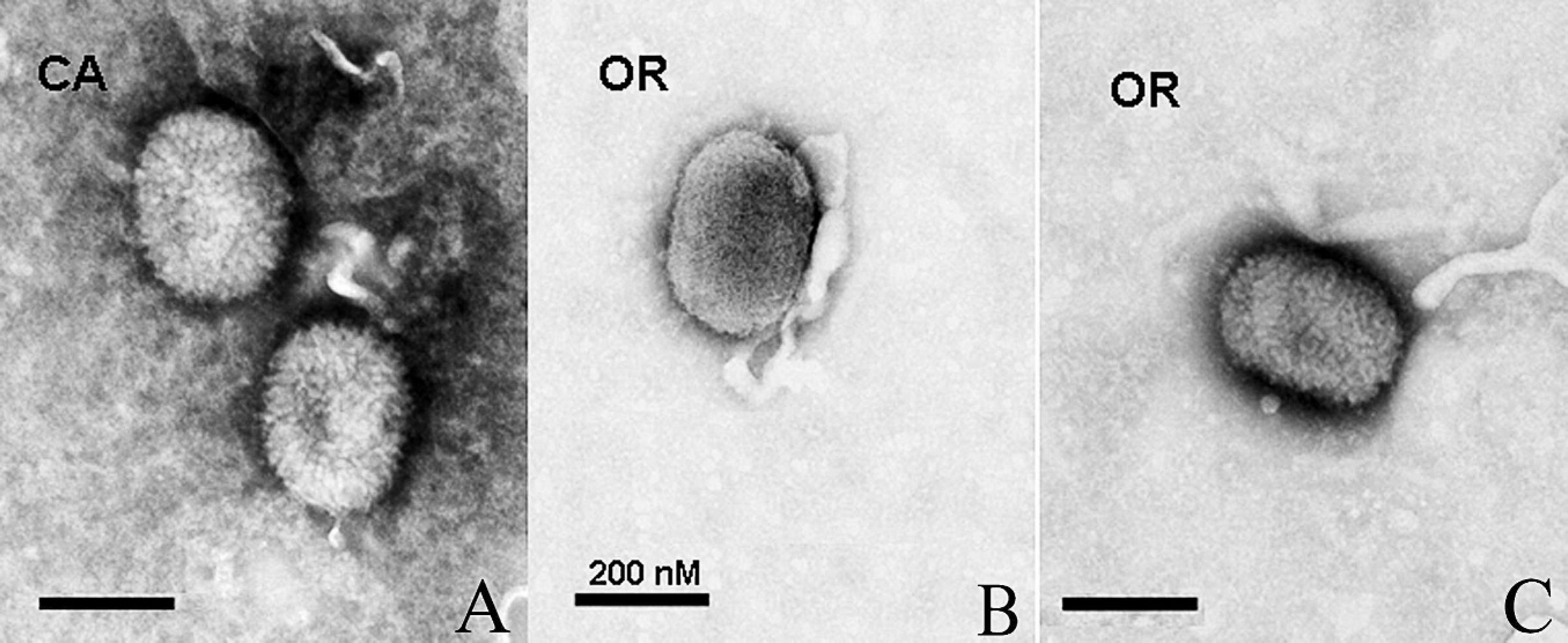
Electron micrographs of Deerpox virus (DPV) virions isolated from California and Oregon (DPV-CA and DPV-OR1, respectively) harvested from infected Vero cells and negatively stained.
A homolog of TGF-β1 was found in the genome of the DPV-83 strain originally characterized, but not in DPV-84, 1 and its presence was investigated in the DPV-CA and DPV-OR1 isolates. A sequence was PCR amplified and characterized from the DPV-CA isolate. The sequence showed 7 differences from the DPV-83 isolate over the 236 amino acids identified. However, the sequence of TGF was not identified in the DPV-OR1 isolate, despite the use of multiple primer pairs. Although this suggested that a homolog of TGF was not present in the DPV-OR1 isolate, it could have been caused by a number of phenomena (e.g., sequence divergence). Therefore, the investigation was not continued (data not shown). The GenBank accession numbers submitted by the authors for A21, CTLR, and IL-1Ra from DPV-CA, DPV-OR1, and DPV-RD are FJ590510–FJ590518, respectively.
Discussion
The previous characterization of DPV was based on isolates obtained from free-ranging deer from 2 locations 40 km apart in Wyoming. The viruses were closely related, and molecular evolutionary analyses suggested that they are members of a distinct genus of the Chordopoxvirinae subfamily. This was additionally supported by the presence in their genomes of several genes homologous to immunomodulatory genes that are not found in other members of the Poxviridae family, 1 which has led to their inclusion in a new genus, Cervidpoxvirus. The 4 isolates characterized in the present study demonstrated a close phylogenetic relationship to the Wyoming isolates and also contained at least 2 of the novel genes associated with CPV, confirming that they likely represent additional members of this genus. A notable aspect of the present investigation is the wide area from which these viruses were isolated, extending the range from the original Wyoming isolates to > 1,000 km of the western United States (separating the DPV CA, and OR1 and OR2 isolates) and to eastern Canada.
The original source of the reindeer isolate is unclear. When the infection was first identified in the reindeer population at the Toronto Zoo, the herd included individuals acquired from the Assiniboine Park Zoo in Winnipeg, Manitoba, as well as stock from Helsinki, Finland. There were also other cervids at the zoo, including white-tailed deer, wapiti, and moose, but they were kept in separate enclosures. The disease has periodically recurred in a mild form in juvenile reindeer since the original diagnosis in 1980 2 and usually is resolved spontaneously in 1–2 weeks. Similar lesions have not been observed in other animals with the exception of woodland caribou, which are the same species as reindeer. The caribou were housed in a separate noncontiguous location from other cervids during the 1980s but are no longer kept by the zoo.
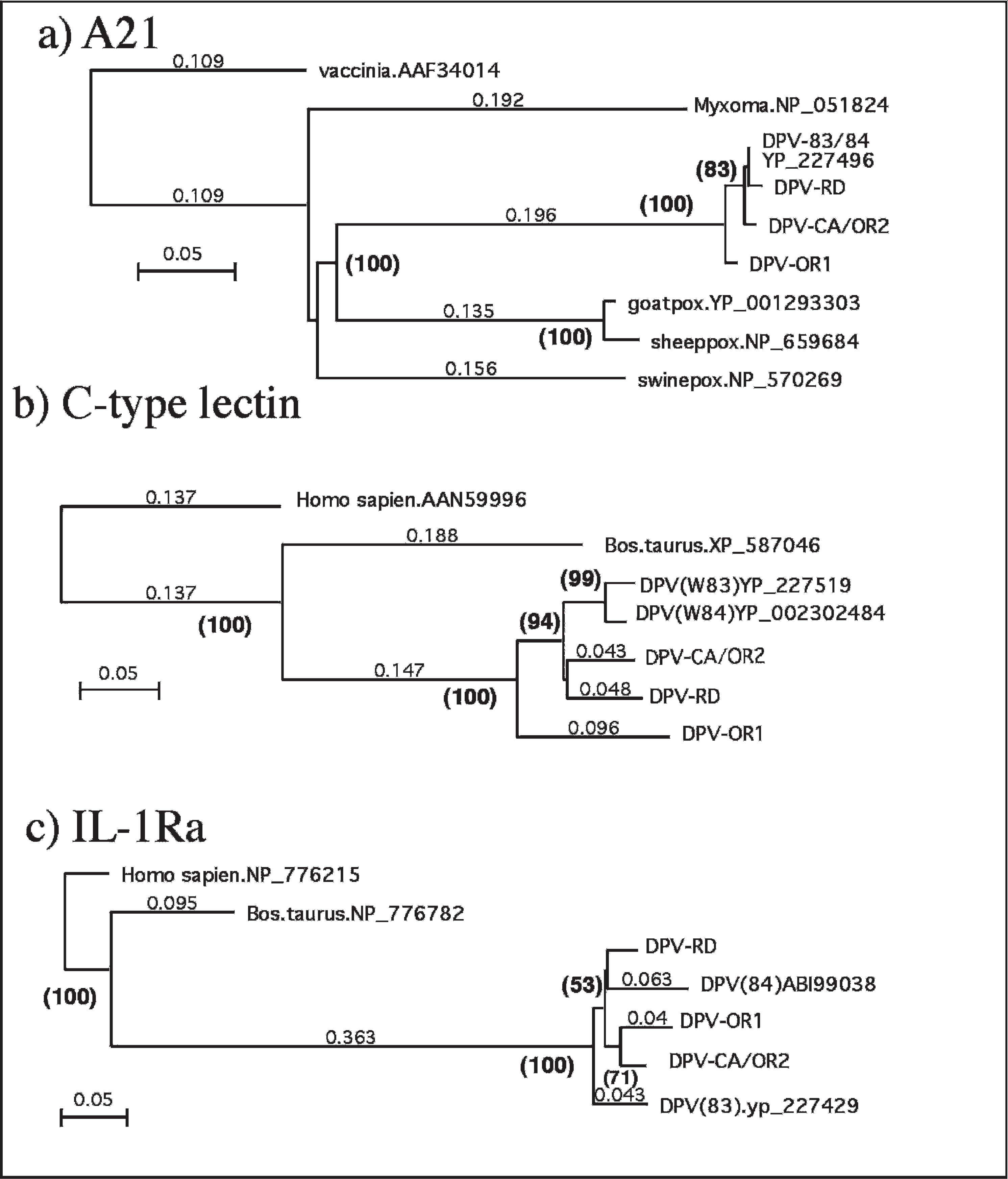
Phylogenetic analysis of selected predicted Deerpox virus (DPV) proteins: A21, C-type lectin, and interleukin-1 receptor antagonist (IL-1Ra). A21 is an open reading frame present in all members of the Poxviridae family. C-type lectin and IL-1Ra homologs have been identified only in DPV isolates. The Homo sapiens and Bos taurus data are nonviral and are included to indicate the relatedness to other members of these protein families.
An unusual feature of the CPV infections is that so far they have only been identified in a limited number of cervids (O. hemionus and reindeer), which belong to the same subfamily, Capreolinae. This is in contrast to members of the Parapoxvirus genus that have been reported from a wide variety of animals, including cervids, ungulates, rodents, and pinnipeds. 3,6,8 The apparent narrow host range of the DPV isolates combined with the presence in their genome of several novel immunomodulatory genes suggest that a feature of the host physiology may have selected for the inclusion and maintenance of these genes, as is typical of well-adapted host–pathogen relationships. Two of the viruses characterized in the present study were from animals kept in confined conditions where cutaneous lesions are easily seen. Although DPV can cause severe infections of young animals that are readily detectable, the infections may be asymptomatic under natural conditions or with healthy animals. Recent experiments involving infection of black-tailed deer fawns support this view. Of 6 animals exposed to the virus, only 1 animal showed symptoms of systemic infection, while the others developed only localized cutaneous lesions and mild transient pyrexia (Bildfell et al., unpublished observations). Further understanding of CPV infections awaits characterization of more isolates, investigation of the role played by the novel genes present in the genomes in the pathophysiology of the infections, and a better understanding of the distribution of the virus in wild cervid populations.
Acknowledgements
The authors thank Marjorie Davis and Wildlife Fawn Rescue for their assistance and for the gross photograph. The following authors contributed equally to this work: Megan Moerdyk-Schauwecker and Kathleen Eide.
Footnotes
a.
Invitrogen Corp., Carlsbad, CA.
b.
Sigma-Aldrich, St. Louis, MO.
c.
Beckman Coulter, Fullerton, CA.
d.
Electron Microscopy Sciences, Hatfield, PA.
e.
VWR International, Bristol, CT.
f.
Philips Electronic Instruments, Mahwah, NJ
g.
Lucigen Corp., Middleton, WI.
h.
ABI PRISM™ 3730 Genetic Analyzer, BigDye™ Terminator v. 3.1 Cycle Sequencing Kit, ABI PRISM™ 3730 Data Collection Software v. 3.0, ABI PRISM™ DNA Sequencing Analysis Software v. 5.2; Applied Biosystems, Foster City, CA.
i.
MacVector Inc., Cary, NC.