
Abstract
The objectives of this study were to determine and compare the in vitro enrofloxacin susceptibility of 94 Pseudomonas aeruginosa isolates obtained from enrofloxacin-treated and untreated mice and that of 40 Pasteurella pneumotropica strains and also to assess the efficacy and effects of enrofloxacin treatment of laboratory mice. The minimum inhibitory concentrations (MICs) of enrofloxacin against all the Ps. aeruginosa isolates were in the range of 1 to 4 μg/ml, whereas those against all the P. pneumotropica strains were less than 0.5 μg/ml. The mutation frequency in 54% of the Ps. aeruginosa isolates on treatment with enrofloxacin ranged from 10−6 to 10−8; however, none of the P. pneumotropica strains could grow on medium containing more than 3 μg/ml enrofloxacin. Comparison of in vitro enrofloxacin susceptibilities suggested that enrofloxacin was effective for eliminating P. pneumotropica but not for eliminating Ps. aeruginosa for which the MIC of enrofloxacin was more than 1 μg/ml. These results indicated that the enrofloxacin susceptibility of P. pneumotropica was higher than that of Ps. aeruginosa, and that the enrofloxacin treatment might not affect the susceptibility of Ps. aeruginosa.
Pasteurella pneumotropica is an opportunistic pathogen of laboratory rodents and is frequently isolated from their upper respiratory tract, lungs, and digestive tract. 2 This bacterium does not significantly affect immunocompetent animals; however, it often causes lethal pneumonia in immunodeficient animals. 10 Therefore, preventing the propagation off. pneumotropica and monitoring infections are important issues in the care and use of laboratory rodents. 2 Although hysterotomy and embryo transfer are known to be some of the most effective treatment measures for the elimination of P. pneumotropica from infected laboratory rodents, they are time-consuming and require special facilities and equipment. 6 Antimicrobial treatment has frequently been recommended as an alternative method for the elimination of P. pneumotropica from laboratory rodents. 6,10,13,20 Ueno et al. 20 reported that the administration of enrofloxacin (a fluoroquinolone) was effective in eliminating this pathogen from infected animals. Enrofloxacin is generally considered to target DNA gyrase and topoisomerase IV that are functional enzymes involved in controlling the topological state of DNA during replication and transcription in prokaryotes. 9 Enrofloxacin is widely used to treat various bacterial infections in animals. 12,15 However, fluoroquinolone-resistant bacteria have been isolated from various animals in recent studies; Pseudomonas aeruginosa, in particular, is well known to exhibit reduced susceptibility to fluoroquinolones, including enrofloxacin. 1,4,11,14,18,19 Ps. aeruginosa is ubiquitously found in the environment and is also an opportunistic pathogen of humans and animals. 11,14,19 This microorganism is frequently isolated from laboratory rodents along with P. pneumotropica and often causes various diseases in immunodeficient animals. 2 Therefore, it is also important to monitor and eliminate Ps. aeruginosa infections in immunodeficient animals. 2 The use of enrofloxacin to eliminate P. pneumotropica in laboratory rodents is thought to affect enrofloxacin susceptibility and occurrence of Ps. aeruginosa. It is important to asses the efficacy of enrofloxacin treatment in the elimination of P. pneumotropica and Ps. aeruginosa from laboratory rodents; however, there have been no reports on the relationship between the enrofloxacin susceptibility of these organisms and their elimination from host animals.
The objectives of this study were to isolate Ps. aeruginosa from enrofloxacin-treated and untreated mice and to compare the minimum inhibitory concentration (MIC) of enrofloxacin against the Ps. aeruginosa isolates and the P. pneumotropica strains. Furthermore, this study was also conducted to determine the in vitro mutation frequencies of these organisms.
In a previous study, of the 94 Institute of Cancer Research (ICR) mice, a 47 were orally administrated enrofloxacin for the elimination of P. pneumotropica, whereas the other 47 mice were left untreated, and the enrofloxacin-treated mice resulted in successful bacterial elimination. 20 Subsequently, Ps. aeruginosa was isolated from those mice in the following manner. Cecal swabs from 94 ICR mice were streaked on nalidixic acid cetrimide (NAC) agar, b and the isolates were identified as Ps. aeruginosa based on colony pigment, Gram staining, and catalase activity.
Thirty-four wild-type strains and 6 reference strains of P. pneumotropica were used in this study. The wild-type strains were isolated from the upper respiratory tract of laboratory mice and rats and a wild mouse; 16,17 the reference strains were ATCC 35149, CCUG 998, CCUG 26450, CCUG 26451, CCUG 26453, and CCUG 36632. Strain ATCC 35149 was obtained from the American Type Culture Collection, and the other 5 strains were obtained from the Culture Collection of the University of Göteborg.
The Ps. aeruginosa isolates were maintained in LuriaBertani medium and the P. pneumotropica strains, in 5% horse-blood agar. To determine the MICs and mutation frequencies, cation-adjusted Mueller-Hinton (CAMH) broth c and CAMH supplemented with 0.05% yeast extract and tryptone (CAMH-YT) were used for Ps. aeruginosa and P. pneumotropica, respectively. The MIC of enrofloxacin was determined by using the standard protocol of the microdilution method; 5 CAMH and CAMH-YT broth were used for Ps. aeruginosa and P. pneumotropica, respectively.
The mutation frequency of the bacterial strains was determined according to the methods of Jumbe et al. 8 with minor modifications. Briefly, 100 μl of CAMH (CAMHYT) broth containing approximately 109 precultured bacterial cells was inoculated on CAMH (CAMH-YT) agar supplemented with enrofloxacin at final concentration of 6 μg/ml or on agar devoid of enrofloxacin. The colonies that grew on the enrofloxacin-supplemented agar after incubation at 37°C for 72 hours were considered to be strains that had mutated because of exposure to enrofloxacin. The mutation frequency was calculated by dividing the number of colonies on the enrofloxacin-supplemented agar by that on the agar devoid of enrofloxacin.
MICs of enrofloxacin and the corresponding numbers of Ps. aeruginosa isolates and P. pneumotropica strains.
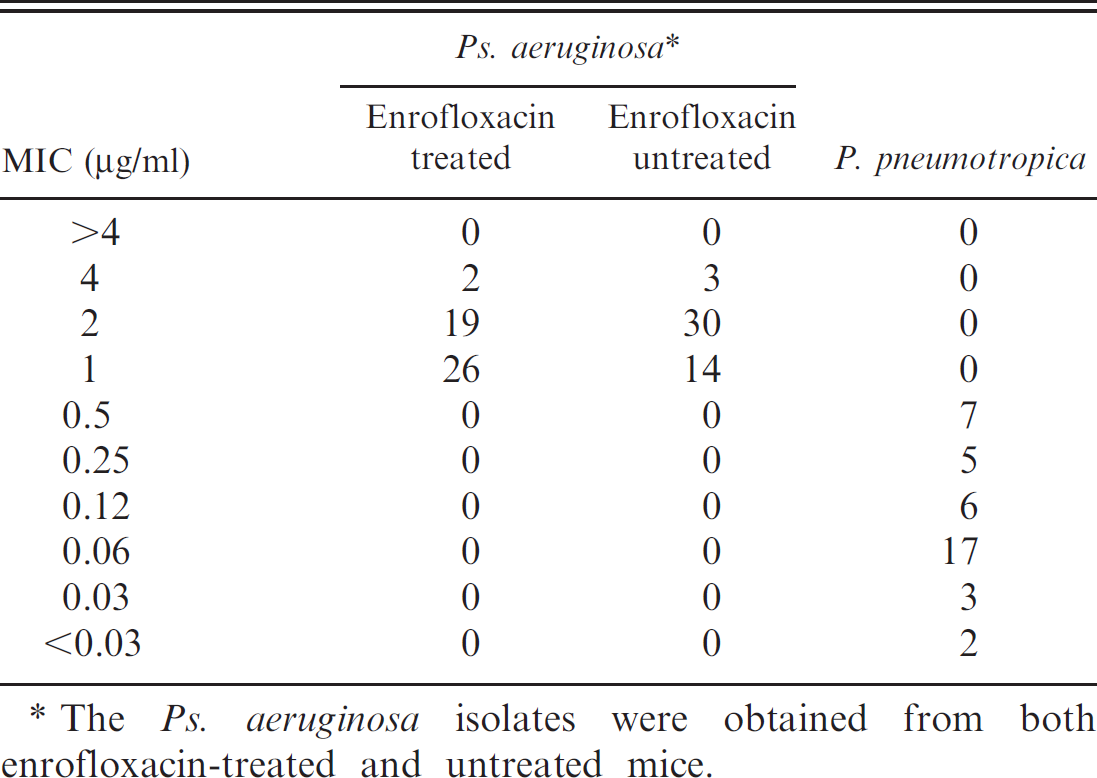
The Ps. aeruginosa isolates were obtained from both enrofloxacin-treated and untreated mice.
The MICs of enrofloxacin against the Ps. aeruginosa isolates from the treated mice were not higher than those against the isolates from the untreated mice (Table 1), and all the Ps. aeruginosa isolates were assumed to be susceptible to enrofloxacin. The MICs of enrofloxacin were also determined for the P. pneumotropica strains, including the 6 reference strains. The MICs of enrofloxacin against all the P. pneumotropica strains in this study were less than 0.5 μg/ml, indicating that P. pneumotropica is highly susceptible to enrofloxacin. The average MIC against the P. pneumotropica strains was more than 20-fold higher than that against the Ps. aeruginosa isolates. Wild-type P. pneumotropica strains have been reported to differ in terms of both phenotypic and genotypic characteristics. 16,17 Namely, the P. pneumotropica strains used in this study were selected from each of 5 groups based on host animals and 16S ribosomal DNA sequence. 17 However, no significant differences were observed among them with regard to their susceptibility to enrofloxacin. Thus, the use of enrofloxacin may be an effective treatment system for eliminating the various P. pneumotropica strains from infected rodents. In contrast, enrofloxacin treatment failed to eliminate Ps. aeruginosa from the mice. Thus, a higher concentration of orally administered enrofloxacin may be required for the elimination of Ps. aeruginosa from the digestive tract than that required for the elimination of P. pneumotropica.
Exposure of bacterial cells to antimicrobial agents has led to reduced susceptibility of the bacteria to these agents. In particular, Ps. aeruginosa, is one of the organisms that exhibit reduced susceptibility to various antimicrobial agents, including fluoroquinolones. 3 In addition, a few wild-type P. pneumotropica strains isolated from mice and rats have been reported to exhibit resistance to enrofloxacin. 7 Further, the mutation frequencies of the Ps. aeruginosa isolates and P. pneumotropica strains were determined by exposing them to enrofloxacin in vitro. The strains that could grow on CAMH or CAMH-YT agar containing 6 μg/ml enrofloxacin were considered to be those that had developed mutations in the enrofloxacin target genes. This is because the MICs against organisms in which the point-mutated genes encoding DNA gyrase, topoisomerase IV, and multidrug efflux pumps are over-expressed are 4− to 8−fold higher than those against the parent organisms. 8 Table 2 lists the numbers that correspond to the mutation frequencies of the Ps. aeruginosa isolates from both the enrofloxacin-treated and untreated mice. Of all the Ps. aeruginosa isolates, 46% did not grow in the enrofloxacin-supplemented medium after 72 hours incubation; the mutation frequencies of these isolates were estimated to be less than −8 log. On the other hand, 54% of the total isolates were able to grow on the enrofloxacin-supplemented medium, and 62% of these isolates that grew on the medium were obtained from the enrofloxacin-treated mice. Moreover, the isolate that exhibited the highest mutation frequency (−5.5 log) was obtained from an enrofloxacin-treated mouse. These results might indicate that more isolates from the enrofloxacin-treated mice than from the untreated mice exhibited increased mutation frequency despite the fact that similar MIC values were observed for isolates from both groups of mice. The mutation frequency of the P. pneumotropica strains was also determined; none of these strains could grow on the medium containing 6 μg/ml enrofloxacin. For the P. pneumotropica strains, the enrofloxacin concentration was increased gradually from 0.75 to 6 μg/ml, and the strains that grew on the medium containing 0.75 μg/ml enrofloxacin were subcultured on the medium containing 3 μg/ml enrofloxacin. Of all the P. pneumotropica strains, 35% were able to grow on the medium containing 0.75 μg/ml enrofloxacin; the mutation frequencies of these strains ranged from −5 to −8 log (data not shown). However, none of the subcultured P. pneumotropica strains could grow on the medium containing more than 3 μg/ml enrofloxacin. These results indicated that the P. pneumotropica strains were highly susceptible to enrofloxacin and that a prolonged period may be required for these strains to exhibit reduced enrofloxacin susceptibility. Based on restriction analysis of 16S ribosomal DNA, Sasaki et al. 16 reported that P. pneumotropica strains could be divided into 4 groups. All the isolates obtained from rats belonging to one of these groups (designated as a-4) could grow on medium containing 0.75 μg/ml enrofloxacin, and they exhibited mutation frequencies in the range of −5 to −7 log; whereas the strains belonging to the other groups exhibited varying mutation frequencies. Hansen and Velschow 7 reported that many P. pneumotropica isolates from rats had a lower susceptibility to enrofloxacin than those from mice. Thus, the mutation frequencies of wild-type P. pneumotropica strains to antimicrobial agents may vary with the host animal. Recent studies revealed that the phenotypic and genotypic characteristics of wild-type P. pneumotropica strains are diverse and complex 16,17 ; therefore, it is necessary to confirm the susceptibility of newly isolated P. pneumotropica strains to enrofloxacin.
Ps. aeruginosa isolates obtained in the medium containing 6 μg/ml enrofloxacin and their corresponding mutation frequencies.
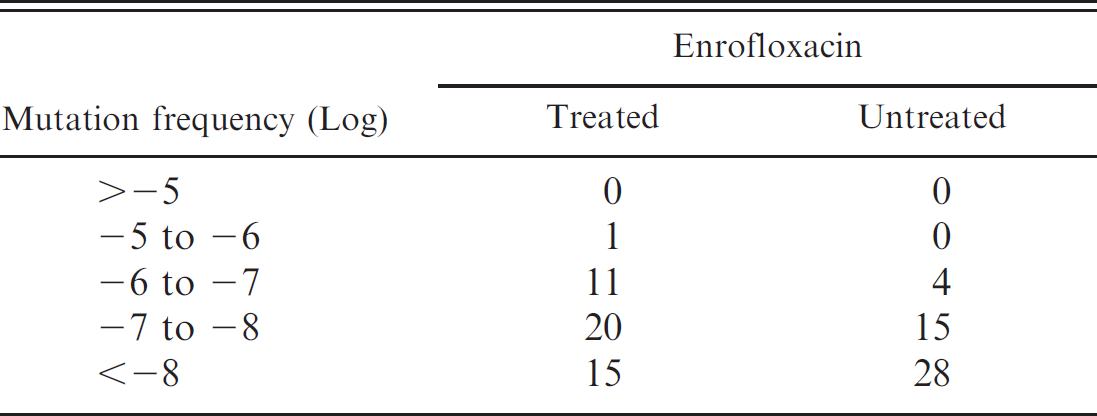
In this study, it was clarified that P. pneumotropica is more susceptible than Ps. aeruginosa to enrofloxacin. Although the increased mutation frequency of the Ps. aeruginosa isolates after treatment with enrofloxacin was found, there were no significant differences in their MIC values between the enrofloxacin-exposed and unexposed isolates. The mechanisms underlying the increase in the mutation frequency of Ps. aeruginosa should be investigated in future studies.
Acknowledgement. This study was partly supported by a Grant-in-Aid from the Ministry of Education, Culture, Sports, Science and Technology, Japan.
Footnotes
a.
ICR mice, Clea, Tokyo, Japan.
b.
NAC agar, Nissui, Tokyo, Japan.
c.
Cation-adjusted Mueller-Hinton broth, BD, Cockeysville, MD.