
Abstract
DACT1 has been shown to participate in the development of many types of tumors; however, its role and precise molecular mechanisms in leukemia are unclear. In this study, we investigated the effect of DACT1 on KG-1α leukemia cells to further understand the mechanisms of DACT1-mediated tumor suppression. We transfected a DACT1 expression plasmid to upregulate DACT1 in KG-1α cells and analyzed the resulting phenotypic changes. The results demonstrated that DACT1 overexpression inhibited KG-1α proliferation, increased apoptosis, and arrested cells in the G0/G1 phase. Mechanistically, DACT1 overexpression inhibited Wnt/β-catenin signaling by reducing nuclear β-catenin levels in KG-1α cells. Furthermore, the viability of KG-1α cells transfected with DACT1 was significantly reduced when treated with daunorubicin. We also found that DACT1 reduced P-glycoprotein expression in KG-1α cells. These findings revealed an inhibitory role for DACT1 in leukemogenesis and provided evidence that DACT1 is an attractive target for the development of novel anti-leukemia therapies.
Introduction
Leukemia is one of the most common malignant tumors in the world and has harmful effects on human health. According to the latest statistics, approximately 370,000 new cases occur and 270,000 people die of leukemia annually, worldwide. 1 In recent years, worldwide leukemia incidences have shown a gradual upward trend. Chemotherapy plays an important role in leukemia treatment.2,3 Yet, most patients cannot be cured by chemotherapy because of drug resistance, and many patients have died because of chemotherapy failures.4,5
The Xenopus protein Dapper is a novel Dishevelled (DVL)-binding protein that has been shown to inhibit Wnt signaling by decreasing β-catenin activation. Dapper Antagonist of Beta Catenin Homolog 1 (DACT1), a homologue of Dapper, was first identified in 2003 and is located at the chromosomal region 14q23.1. 6 Many studies have confirmed that DACT1 is expressed both in lower vertebrates (such as zebrafish) and in higher vertebrates (such as humans), suggesting that it plays important physiological functions.7–10 Wnt signaling is a highly conserved fundamental signaling pathway that regulates cell proliferation, differentiation, and apoptosis.11–14 Wnt family ligands control three distinct signaling pathways, the Wnt/β-catenin pathway, Wnt/Ca2+ pathway, and Wnt planar cell polarity (PCP) pathway. Canonical Wnt/β-catenin signaling controls many biological processes, including playing a key role in the cancer development.15,16
A previous study by Yin et al. 17 reported that DACT1 antagonized Wnt/β-catenin signaling in breast cancer and inhibited breast cancer cell proliferation in vivo and in vitro. However, Yuan et al. 18 has put forward the opposite view, suggesting that DACT1 regulates the stability of β-catenin and is an important positive regulator of Wnt/β-catenin signaling. There are still controversies regarding the role of DACT1 in the Wnt signaling pathway. In this study, we investigated the role of DACT1 in KG-1α leukemia cells, further elucidating the role of DACT1 in tumor suppression.
Materials and methods
Cell lines and culture
KG-1α cells were maintained in RPMI-1640 (Gibco, Grand Island, NY, USA) at 37°C in a humidified incubator with 5% CO2. The culture medium was supplemented with 10% fetal bovine serum (Hyclone, Thermo Fisher Scientific, Waltham, MA, USA).
Transfection
Cells were transiently transfected using Attractene transfection reagent (QIAGEN, Hilden, Germany) according to the manufacturer’s instructions. Cells were transferred to six-well plates; each well contained 4 × 105 cells. Then, 100 µL serum-free RPMI-1640 medium, 1.2 µg DACT1, or vector plasmid was mixed with Attractene transfection reagent (4.5 µL/well) and incubated for 20 min at room temperature. The solution was then added to each well containing 1 mL Dulbecco’s Modified Eagle’s Medium (DMEM) (HyClone Laboratories Inc., Logan, UT, USA) and incubated for 6 h. After incubation, the medium was replaced with RPMI-1640 medium containing 20% serum, and cells were incubated at 37°C in a humidified incubator with 5% CO2.
Quantitative real-time polymerase chain reaction
Total RNA was extracted from cells using RNA pure total RNA fast isolation kit following the manufacturer’s protocols (BioTeke, Beijing, China). First-strand complementary DNA (cDNA) was synthesized using Super M-MLV Reverse transcriptase (BioTeke). The primers used in quantitative real-time polymerase chain reaction (RT-qPCR) reactions were as follows: DACT1 sense: 5′-CCGAACAAGCCGAAAGCAAGA and anti-sense: 5′-AATGGCGGAACACCGAGCA; β-catenin sense: 5′-GCCAAGTGGGTGGTATAGAG and anti-sense: 5′-GTGGGATGGTGGGTGTAAGA; β-actin sense: 5′-CTTAGTTGCGTTACACCCTTTCTTG and anti-sense: 5′-CTGTCACCTTCACCGTTCCAGTTT. RT-qPCR was performed using a SYBR Green reaction kit (TaKaRa, Dalian, China) on an Exicycler™96 thermal block (Bioneer, Daejeon, Korea). PCR reactions were amplified for 40 cycles. Each cycle consisted of denaturation for 10 s at 95°C, annealing for 20 s at 60°C, and polymerization for 30 s at 72°C.
Western blotting
Cells were harvested, washed with phosphate-buffered saline (PBS), and lysed in lysis buffer (Beyotime Institute of Biotechnology, Shanghai, China). Then, lysates were centrifuged at 12,000g for 10 min at 4°C, and supernatants were collected. Protein concentration was measured using the bicinchoninic acid (BCA) assay kit (Pierce Biotechnology, Thermo Fisher Scientific, Rockford, IL, USA) following the manufacturer’s instructions. Then, protein extracts were separated via polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride membranes (Millipore, Bedford, MA, USA). Protein-bound membranes were blocked in 5% skim milk solution and incubated with appropriate primary antibodies overnight at 4°C. Primary antibodies were diluted as follows: 1:2000 for DACT1 (Abcam, Cambridge, MA, USA) and 1:1000 for Bax, Bcl-2, β-catenin, Cyclin D1, C-myc, interleukin (IL)-8, and P-gp. Protein levels were normalized to β-actin (1:1000; Abcam). The next day, membranes were incubated with goat anti-rabbit peroxidase-conjugated secondary antibody (1:5000; Abcam). Finally, blots were analyzed using an ECL plus western blotting detection system (Beyotime Institute of Biotechnology).
Cell proliferation assay
The proliferation of KG-1α cells in 96-well plates was examined using Cell Counting Kit-8 (CCK-8; Beyotime Institute of Biotechnology) according to the manufacturer’s instructions. Briefly, 10 µL CCK-8 solution and 100 µL RPMI-1640 medium were added to each well and incubated for 1 h. The absorbance at 450 nm was measured using a spectrophotometer (Bio-Tek, Winooski, VT, USA).
Colony-formation assay
Cells were plated in dishes at a density of 400 cells/dish and colony formation was monitored for 14 days at 37°C in a humidified atmosphere with 5% CO2. The number of colonies containing 50 or more cells was then counted.
Annexin V apoptosis assay
Cells were trypsinized, washed with ice-cold PBS twice, and collected by centrifugation. Then, cells were resuspended in binding buffer, and Annexin V–fluorescein isothiocyanate (FITC) and propidium iodide (PI; BD Pharmingen, San Diego, CA, USA) were added to the fixed cells for 15 min in the dark at room temperature according to the manufacturer’s instructions. After incubation, cells were analyzed using a flow cytometer (BD Biosciences, San Jose, CA, USA).
Cell cycle analysis
Cell cycle analysis was performed using a Cell Cycle Analysis Kit (Beyotime Institute of Biotechnology) following the manufacturer’s instructions at 24, 48, and 72 h after transfection. Cells were washed with PBS and fixed with 70% cold ethanol for 2 h at 4°C. After washing, 100 µL RNase A was added to the cells for 30 min at 37°C, and then the cells were stained with PI for 30 min at 4°C in the dark. Results were collected using a flow cytometer.
Immunocytofluorescence
Cells were fixed in 3.7% paraformaldehyde for 15 min at room temperature, permeabilized with 0.1% Triton X-100 in PBS for 30 min, blocked in 10% normal goat serum (Solarbio, Beijing, China) with PBS for 15 min, and incubated with mouse monoclonal anti-β-catenin antibody (1:200; Abcam) overnight at 4°C. The next day, cells were washed three times with PBS and incubated with secondary goat anti-mouse IgG antibody (1:200; Beyotime Institute of Biotechnology) for 1 h at 37°C in the dark; DAPI (4′,6-diamidino-2-phenylindole) was used for nuclear counterstaining. Finally, coverslips were mounted with anti-fade mounting medium (Solarbio) and visualized with a fluorescence microscope (BX53; OLYMPUS, Tokyo, Japan).
Statistical analysis
Values are expressed as mean ± standard deviation (SD) from at least three independent experiments. Statistical analyses were carried out with the SPSS 20.0 statistical software package (SPSS Inc., Chicago, IL, USA). The significance of differences between groups was evaluated by Student’s t-test; a value of p < 0.05 was considered statistically significant.
Results
DACT1 overexpression inhibits KG-1α cell proliferation
As shown in Figure 1(a) and (b), DACT1 messenger RNA (mRNA) and protein levels were significantly increased in DACT1-transfected cells compared with control cells. To investigate the relationship between DACT1 overexpression and the viability of KG-1α cells, the CCK-8 cell-proliferation assay and colony formation assays were performed. Cell viability was significantly decreased at 24, 48, and 72 h after DACT1 transfection into KG-1α cells (p < 0.05) (Figure 1(c)). Colony-formation assays showed that DACT1 overexpression was associated with reduced colony formation (p < 0.05); DACT1-transfected cells had a lower colony formation ability compared with control cells (Figure 1(d)).

Tumor-suppressive functions of DACT1 in KG-1α cells. (a) The expression of DACT1 mRNA was measured with RT-qPCR. DACT1 mRNA expression was significantly upregulated in DACT1-transfected cells compared with the controls. (b) The expression of DACT1 protein was detected with western blot analysis. DACT1 protein expression was significantly increased in DACT1-transfected cells compared with the controls. (c) CCK-8 assay assessed the effect of DACT1 on cell proliferation in KG-1α, KG-1α/vector, and KG- 1α/DACT1 cells. (d) Colony-formation assay of DACT1 in KG-1α, KG-1α/vector, and KG-1α/DACT1 cells. Mean ± SD of three assays.
DACT1 overexpression increases KG-1α apoptosis
To further examine the effects of DACT1 overexpression on apoptosis, we performed an apoptosis assay using flow cytometry. The results demonstrated that DACT1-transfected cells exhibited higher apoptotic rates than control cells (Figure 2(a) and (b)). To evaluate the molecular mechanism responsible for DACT1-mediated apoptosis, levels of proteins involved in apoptosis were determined by western blot. The results showed that the pro-apoptotic protein Bax was upregulated and the anti-apoptotic protein Bcl-2 was downregulated in DACT1-transfected cells (Figure 2(c)).
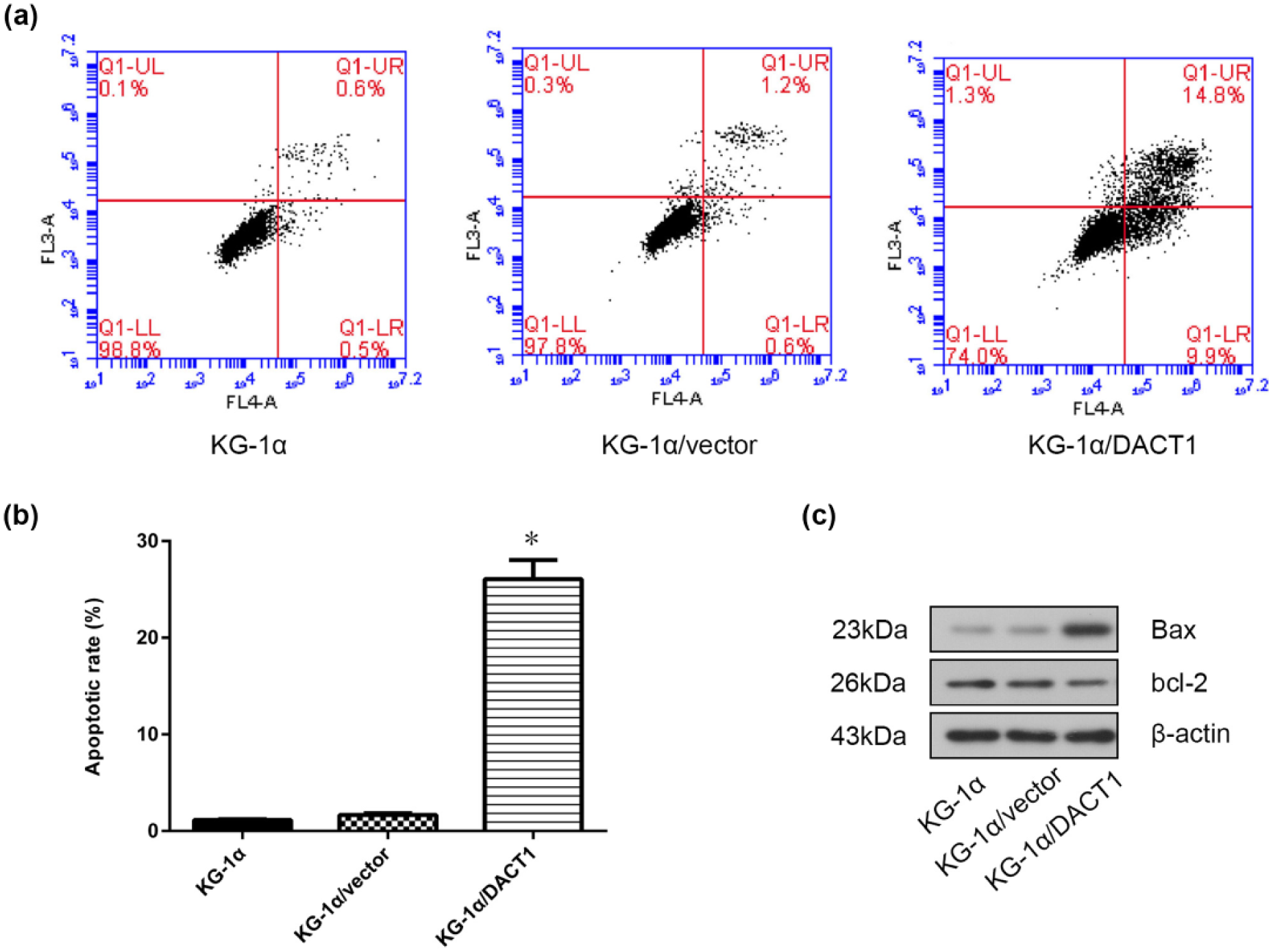
Effects of DACT1 overexpression on KG-1α cells apoptosis. (a) Cell apoptosis was determined with FACS analysis. DACT1 overexpression significantly induced apoptosis in KG-1α cells. The representative FACS analysis is shown. (b) Representative results from three independent experiments are shown. (c) Levels of apoptosis-related proteins including Bcl-2 and Bax were determined by western blotting analysis.
DACT1 overexpression arrests KG-1α cells in G0/G1
To determine whether DACT1 overexpression affects cell cycle progression, cells were subjected to flow cytometric analysis, and the percentages of cells in G0/G1, S, and G2/M were calculated. Our results showed that DACT1 overexpression led to a significant increase in the number of cells in G0/G1, suggesting a strong cell cycle arrest at the G0/G1 phase after transfection compared with control cells (Figure 3).

Effects of DACT1 overexpression on KG-1α cells cycle distribution. (a) Cell cycle distribution was determined with FACS analysis. The representative FACS analysis is shown. (b) Data showed G0/G1 arrest in KG-1α cells transfected with DACT1.
DACT1 overexpression inhibits Wnt/β-catenin signaling
To further investigate the role of DACT1 in Wnt signaling pathways, we performed western blot and immunocytofluorescence assays. Our results showed that DACT1-transfected cells had decreased β-catenin mRNA and protein expression and reduced expression of Wnt/β-catenin downstream target genes, including cyclin D1, c-myc, and IL-8 (Figure 4(a) and (b)). Results from the immunofluorescence assays suggested that DACT1 reduced nuclear β-catenin levels in KG-1α cells (Figure 4(c)).

DACT1 suppressed Wnt/β-catenin pathway. (a) β-catenin mRNA expression was significantly decreased in DACT1-transfected cells compared with the controls. (b) Western blotting analysis of β-catenin and target protein of Wnt/β-catenin pathway. (c) Subcellular location of β-catenin in KG-1α, KG-1α/vector, and KG-1α/DACT1 cells by immunofluorescence assays. Scale bars represent 50 µm.
DACT1 overexpression increases daunorubicin chemosensitivity in KG-1α cells
To investigate whether DACT1 is involved in KG-1α daunorubicin sensitivity, DACT1-transfected and control cells were incubated with 1 µg/mL daunorubicin for 24, 48, and 72 h, and cell viability was determined using CCK-8 cell-proliferation assays. These results showed that cell viability was significantly decreased in DACT1-transfected cells (p < 0.05). DACT1 overexpression also increased KG-1α cell chemosensitivity to daunorubicin (Figure 5(a)). To further investigate the mechanism through which DACT1 increased chemosensitivity, P-gp protein levels were examined in DACT1-overexpressing cells and showed downregulation compared with control cells (Figure 5(b)).

Effects of overexpression of DACT1 on chemosensitivity of KG-1α cells to daunorubicin. (a) Cell viability after 1 µg/mL daunorubicin treatment at 24, 48, and 72 h was measured by CCK-8 assay. Data are shown as mean ± SD of triplicate experiments. (b) Western blotting was used to assess P-gp expression in KG-1α, KG-1α/vector, and KG-1α/DACT1 cells.
Discussion
DACT1 has been reported to play a role in the development and progression of several human cancers. A recent paper showed that DACT1 was frequently downregulated in breast cancer. 17 DACT1 overexpression inhibits cell growth in vivo and in vitro, highlighting the crucial function of DACT1 in tumor suppression. However, the exact function and underlying mechanisms of DACT1 in leukemia are largely unknown.
In this study, we investigated the tumor suppression roles of DACT1 in KG-1α leukemia cells, finding that DACT1 negatively regulated KG-1α cell proliferation. This result was consistent with previous reports; Wang et al. 19 demonstrated that ectopic DACT1 expression in DACT1-silenced gastric cancer cell lines suppressed colony formation and retarded tumorigenesis in nude mice. However, in colon cancer, it was shown that DACT1 promoted cell proliferation in vitro and tumor growth in vivo and enhanced the migratory and invasive potential of colon cancer cells. 18
Our study showed that DACT1 overexpression led to KG-1α cell apoptosis. KG-1α cells transfected with DACT1 had a significantly higher percentage of apoptotic cells than the control group. In addition, DACT1-transfected cells had increased expression of pro-apoptotic proteins Bax and decreased expression of anti-apoptotic proteins Bcl-2, revealing that DACT1 may activate intrinsic apoptotic pathways. Cell cycle analysis showed that DACT1 overexpression induced KG-1α cells into a G0/G1 phase arrest. Taken together, these results suggested that DACT1 participates in the pathogenesis of KG-1α cells.
To determine the mechanisms of the tumor-suppressing effects of DACT1, we investigated the Wnt/β-catenin signaling pathway. It has been confirmed that Wnt signaling inhibits apoptosis and promotes cell growth and the development of multidrug resistance after chemotherapy.20–22 In the absence of Wnt ligands, free cytosolic β-catenin is marked for proteasomal degradation by the complex composed of Axin, APC, GSK3-β, and CK1 and cannot enter the nucleus to drive target gene transcription. In the presence of Wnt ligands, Wnt proteins combine with the cell surface receptor Frizzled and its co-receptor LRP5/6, leading to Axin degradation and the depolymerization of the degradation complex. Dvl is then recruited to the cell membrane, resulting in the phosphorylation of GSK3-β and an accumulation of cytoplasmic β-catenin, which can translocate to the nucleus and combine with LEF/TCF (T-cell factor/lymphoid enhancer factor) transcription factors to form the TCF/LEF/β-catenin complex. This complex specifically promotes the expression of downstream Wnt target genes such as c-myc, c-Myb, Cyclin D1, ETS, BMP4, EPHB2, EPHB3, CD44, MMP7, and IL-8. 23
Wnt/β-Catenin signaling has also been shown to contribute to the development of acute leukemia. Aberrant Wnt/β-catenin pathway activation has been implicated in leukemic transformation24,25 and shown to promote the proliferation and survival of leukemic cells in vitro. 26 β-catenin is a transcriptional regulator of canonical Wnt signaling after it has been imported into the nucleus. To identify whether DACT1 overexpression decreased nuclear β-catenin, we performed immunocytofluorescence assays. DACT1 overexpression decreased β-catenin mRNA and protein expression and the nuclear translocation of β-catenin. These data suggested that DACT1 expression was associated with decreased nuclear β-catenin. We next detected the expression of downstream Wnt pathway targets including c-Myc, Cyclin D1, and IL-8. The results showed that DACT1 overexpression inhibited the expression of Wnt targets, further explaining how DACT1 overexpression inhibited Wnt/β-catenin signaling.
Our results also showed that DACT1 increased KG-1α cell daunorubicin chemosensitivity. The viability of DACT1-overexpressing KG-1α cells was significantly reduced by daunorubicin, suggesting that DACT1 upregulation enhanced the sensitivity of KG-1α cells to daunorubicin. We further found that DACT1 reduced P-gp expression in KG-1α cells. In leukemia, expression of the product of ABCB1, P-gp, is associated with the development of drug resistance. 27 Recent reports have suggested that Wnt signaling is involved in multidrug resistance. The Wnt/β-catenin pathway was shown to regulate ABCB1 in many cancer types.28,29 Moreover, the study by Correa et al. 30 showed that the canonical Wnt/β-catenin pathway was involved in ABCB1 transcriptional activation in chronic myeloid leukemia and positively regulated ABCB1 and P-gp expression. β-catenin could bind to the ABCB1 promoter at the TCF-binding site. In this study, DACT1 overexpression decreased β-catenin nuclear translocation, suggesting that inhibiting canonical Wnt/β-catenin signaling leads to P-gp downregulation in KG-1α cells and an increase in the sensitivity of KG-1α cells to daunorubicin.
In conclusion, our findings revealed an inhibitory role for DACT1 in tumorigenesis of KG-1α cells and provided new insight into the molecular mechanisms underlying acute leukemia. Our results provide evidence that DACT1 is an attractive target for novel anti-leukemia therapy development and offers a promising option for gene therapy in the future.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.