
Abstract
Fibroblast growth factor receptors are growth factor receptor tyrosine kinases, exerting their roles in embryogenesis, tissue homeostasis, and development of breast cancer. Recent genetic studies have identified some subtypes of fibroblast growth factor receptors as strong genetic loci associated with breast cancer. In this article, we review the recent epidemiological findings and experiment results of fibroblast growth factor receptors in breast cancer. First, we summarized the structure and physiological function of fibroblast growth factor receptors in humans. Then, we discussed the common genetic variations in fibroblast growth factor receptors that affect breast cancer risk. In addition, we also introduced the potential roles of each fibroblast growth factor receptors isoform in breast cancer. Finally, we explored the potential therapeutics targeting fibroblast growth factor receptors for breast cancer. Based on the biological mechanisms of fibroblast growth factor receptors leading to the pathogenesis in breast cancer, targeting fibroblast growth factor receptors may provide new opportunities for breast cancer therapeutic strategies.
Introduction
Breast cancer was the most commonly diagnosed cancer and the leading cause of cancer deaths in women worldwide. An estimated 1.7 million cases and 521,900 deaths occurred in 2012, and it accounts for 25% of all cancer cases and 15% of all cancer deaths among females. 1 The etiology of breast cancer is generally considered as multiple genetic predisposition combined with environmental factors. Importantly, the fibroblast growth factor/fibroblast growth factor receptor (FGF/FGFR) signaling pathway appears to play crucial role in the development and progression of breast cancer. 2 As necessary components in this pathway, FGFRs are infrequently mutated but frequently overexpressed in breast cancer. FGF signaling is susceptible to subversion by cancer cells though modulating key cell proliferation, differentiation, and survival.3,4 The FGFR family includes five key members: FGFR1–4 and FGFR5 (also known as FGFRL1). FGFR1–4 are typical growth factor receptor tyrosine kinases containing an extracellular domain, a trans-membrane domain and a cytoplasmic domain, while FGFR5 lacks the intracellular kinase domain. Moreover, FGFR1–4 play important roles both in physiological and pathological processes of human tissues and organs, but FGFR5 has many unclear functions. 5
Genome-wide association studies (GWAS) have identified a significant association between some genetic variants and breast cancer risk.6–10 Among these variants, an intronic variant in the FGFR2 gene is currently identified as one of the most important genetic susceptibility locus in breast cancer. 10 Furthermore, several single-nucleotide polymorphisms (SNPs) in FGFR2 were confirmed to have a high correlation with breast cancer risk. 11 Moreover, recent case-control studies also indicate that some SNPs in FGFR3 and FGFR4 genes were closely related to breast cancer risk.12,13 In this article, we will focus on the main FGFRs genetic polymorphisms and their possible physiological functions that are related with breast cancer. The potential roles and pathogenesis of FGFRs in breast cancer have also been discussed. In addition, we will summarize the recent advances and challenges in targeting FGFRs for the prevention and treatment of breast cancer.
Structure and physiological function of FGFRs
The FGFR family includes five key members linked to multiple intracellular downstream signaling pathways: FGFR1–4 and FGFR5 (also known as FGFRL1). FGFR1–4 are typical growth factor receptor tyrosine kinases containing an extracellular domain, a single-pass trans-membrane domain and a carboxy-terminal cytoplasmic domain. 5 The extracellular portion comprises three immunoglobulin (Ig)-like domains (IgI, IgII, and IgIII) with the acidic box, which is a run of glutamic acid and aspartic acid residues in the IgI–IgII linker region. 14 IgI and the acid box in the amino-terminal portion are involved in autoinhibition, while the IgII and IgIII domains are believed to be necessary for ligand binding. 15 The intracellular portion consists of a juxta-membrane domain, a split tyrosine kinase domain, and a carboxy-terminal tail (Figure 1(a)). 16 This receptor system contains heparan sulfate proteoglycans and related heparin-like molecules, which are necessary for FGF–FGFR binding and FGFR activation. 17 Like FGFR1–4, the N-terminal extracellular region of FGFR5 includes a putative signal peptide, three typical Ig-like domains, six cysteines, and an acidic box. 18 The C-terminal region of FGFR5 contains a trans-membrane domain but lacks an intracellular kinase domain (Figure 1(b)). FGFR5 has few clear functions; however, it could form heterodimers combining with other FGFRs and affect signal transduction. 19 In addition, alternative splicing of exons 8 and 9 encoding the IgIII extracellular fragment of FGFR1–3 leads to the expression of two distinct receptor isoforms (IIIb and IIIc). These isoforms have different ligand binding specificity for IIIb and IIIc isoforms predominantly expressed in the epithelium and mesenchymal tissues, respectively. 20 However, the binding specificity of isoform variants can also transform when referring to cancers. Moreover, FGFR1–2 also have the third isoform named IIIa, which may produce truncated and secreted receptors. Hence, FGFR1–2 may have an autoinhibitory function and unable to transduce signals in FGF/FGFR signaling pathway. 21 FGFR4 is reported to possess only one isoform that is similar to the IIIc variant of FGFR1–3.

Structure of the fibroblast growth factor receptor (FGFR) family of receptor tyrosine kinases. (a) FGFR1–4 are typical growth factor receptor tyrosine kinases containing an extracellular domain, a single-pass trans-membrane domain and a carboxy-terminal cytoplasmic domain. The extracellular portion is comprised of three immunoglobulin (Ig)-like domains (IgI, IgII, and IgIII) with the acidic box, which is a run of glutamic acid and aspartic acid residues in the IgI–IgII linker region. IgI and the acid box in the amino-terminal portion are involved in autoinhibition while the IgII and IgIII domains are believed necessary for ligand binding. The intracellular portion is consisted of a juxta-membrane domain, a split tyrosine kinase domain, and a carboxy-terminal tail. (b) FGFR5 includes a putative signal peptide, three typical Ig-like domains, six cysteines, and an acidic box. The C-terminal region of FGFR5 contains a trans-membrane domain but lacks an intracellular kinase domain.
With respect to the function, FGFR1–4 not only act as inducers of mesoderm participating in various processes during embryonic development but also play important roles in organogenesis, especially in the nervous system, the limbs, the midbrain, and the lungs. 22 In addition, FGFR1–4 appear to be involved in maintaining homeostasis in normal human tissues and regulating tissue repair, angiogenesis, and inflammation in the adult organism. 23 Similarly, FGFR1–4 have been implicated in diverse cellular processes including cell growth, survival, differentiation, and migration. Besides, they also take part in pathological processes, including angiogenesis, wound healing, and cancer. 24 In cancer cells, when FGFRs bind to different FGF ligands, FGFRs can lead to the abnormal upregulation of the Ras-dependent mitogen-activated protein kinase (MAPK), Ras-independent phosphoinositide-3-kinase-proteinkinase B/Akt (PI3K-PKB/Akt), and signal transducer and activator of transcription (STAT)-dependent signaling pathways, which are closely associated with the development of multiple cancers (Figure 2). 25

FGFR signaling network. When FGFRs bind to different FGF ligands, it causes receptor dimerization and tyrosine domains transphosphorylation of FGFR. This process leads to upregulation of four signaling pathways: MAPK, PI3K/AKT, STAT, and PLCγ. These pathways finally regulate transcription of target genes, which are associated with cancer cell proliferation, differentiation, invasion, and survival. While PLCγ signaling pathway act as a negative regulator, which is different from other pathways.
Apart from these common functions, FGFR1–4 possess their own characteristics. As a co-receptor, FGFR1 is required during adeno-associated virus 2 infection for successful viral entry into the host cell. 26 FGFR4 is involved in myogenesis and muscle regeneration with a particular autophosphorylation rate and tyrosine kinase activity compared to other FGFRs. 27 Moreover, although FGFR5 has many unclear functions, recent studies indicated that FGFR5 may provide a binding site for one or more FGFs which regulates pancreatic function. 18
Genetics of FGFR genes in breast cancer
Since FGFRs play a vital role in the pathology of cancer, and these functions may result from different FGFR pathway aberrations: (1) receptor overexpression caused by gene amplification or post-transcriptional regulation; (2) FGFR mutations producing receptors which exhibit a reduced dependence on ligand binding for activation; (3) translocations leading to the expression of FGFR-fusion proteins with constitutive FGFR kinase activity; (4) isoform switching and alternative splicing of FGFR substantially alters ligand specificity increasing the range of FGFs, and finally leads to the stimulation of tumor cells; (5) the overexpression of FGF in cancer or stromal cells and the upregulation of FGFs from the extracellular matrix lead to paracrine/autocrine activation of the pathway. Table 1 shows common FGFR genomic deregulations in breast cancer, and we can see that breast cancer was mainly caused by amplification in FGFR2 and FGFR4, the details are discussed subsequently.
Reported association studies of FGFR variants with breast cancer.

FGFR: fibroblast growth factor receptor; SNP: single-nucleotide polymorphism; OR: odds ratio; CI: confidence interval; GWAS: Genome-wide association studies; SEARCH: population-based study of breast cancer in UK; EPIC-Norfolk: European Prospective Investigation into Cancer in Norfolk cohort study; NHS: Nurses’ Health Study cohort; PLCO: the Prostate, Lung, Colorectal and Ovary Cancer Screening Trial Cohort; CPS-II: the American Cancer Society Cancer Prevention Study-II; NA: not available.
Breast cancer appears to have familial aggregation, which is consistent with variations in genetic susceptibility to the disease. 31 Beyond all the known breast cancer susceptibility genes (less than 25%), more than 75% genetic variants of the familial risk of the disease remain unexplored which may confer more risks. 32 The GWAS identified several genomic locations in the FGFR2 gene as potentially associated with breast cancer. The first identified SNP is rs2981582 which lies in intron 2 of FGFR2 (also known as CEK3), and it was proved as a novel breast cancer susceptibility loci. The team of Douglas demonstrated that multiple haplotypes carrying the minor allele of rs2981582 could increase the risk of breast cancer. 10 Then, some other SNPs were revealed to link to breast cancer risk. SNPs rs1219648 and rs2420946 were in intron 2 of FGFR2 encoding a receptor tyrosine kinase, which is associated with the development and neoplasia of mammary gland. All four (additional variants rs11200014 and rs2981579) SNPs in FGFR2 have common characteristics in terms of their correlation with breast cancer risk, in line with the high degree of linkage disequilibrium. 11 Further studies found that rs2981582 was more strongly associated with estrogen receptor (ER)-positive than ER-negative breast cancer. Moreover, this SNP was more closely correlated with progesterone receptor–positive, low grade, and node-positive breast cancer. 28 Recently, a Chinese study team enrolled 1049 breast cancer patients and 1073 cancer-free controls. They assessed whether polymorphisms of FGFR2 are also associated with breast cancer risk in Chinese women and whether these associations are stronger in women with a reproductive history suggestive of greater exposure to endogenous estrogens. Their results revealed that rs2981582C/T, rs1219648A/G, and rs2420946C/T were significantly associated with increased breast cancer risk in a dose-dependent manner and indicated that genetic variants in FGFR2 may contribute to breast cancer occurrence in Chinese women, possibly through pathways related to estrogen and/or progesterone. 29 Currently, another study group analyzed the relationship between 14 tag SNPs and breast cancer risk in the Han Chinese population, including 185 breast cancer patients and 199 healthy women controls of the different types of breast cancer and menopausal status. They found rs2981579 in the FGFR2 gene was associated with breast cancer susceptibility. 30 These findings strongly indicated that FGFR2 is a breast cancer susceptibility gene.
To identify whether common variation in other FGFRs also affects breast cancer risk, many case-control studies were carried out in diverse populations recently. A study conducted on northern Chinese population suggested that SNPs rs1966265 and rs351855 in FGFR4 gene were strongly associated with breast cancer risk. After further analyses of the disease characteristics, SNP rs351855 was proved to be related with lymph node–positive breast cancer in a dose-dependent effect of the minor allele. 13 However, little evidence of association was found between breast cancer risk with SNPs in the FGFR1, FGFR3, FGFR4, and FGFRL1 genes in another multicenter case-control study. The strongest evidence in European women was for rs743682 in FGFR3, while it is still markedly inferior to that observed for SNPs in FGFR2. 12
The potential roles of FGFRs in breast cancer
FGFR1 in breast cancer
Amplification of the chromosomal region of FGFR1 (8p11–12) has been detected in approximately 10% of human breast cancers, predominantly in ER-positive cancers, and it has a pejorative effect on overall survival in breast cancer.3,33 In addition, amplification and high expression of FGFR1 in non-transformed human mammary cells would result in morphological transformation characterized by epidermal growth factor–independent cell growth, anchorage-independent cell proliferation and survival, loss of cell polarity, and epithelial-to-mesenchymal transition. 34 FGFR1 maps to the proximal A2 core of 8p11–12, and its amplification correlates to reduced metastasis-free survival. Recently, it has been reported that activation of FGFR1 may drive invasive behavior of breast cancer cells. 33 FGFR1 amplifications happen more frequently in the invasive components of tumors and more commonly in invasive breast cancer than ductal carcinoma in situ. 35 In vitro and in vivo studies have suggested that FGFR1 amplification and overexpression would lead to oncogene addiction and modulate invasive cell behaviours. 20
In addition, it is reported that amplification of 8p11–12 appears in approximately 15% of human breast cancer, and this region of amplification is significantly associated with disease-specific survival and distant recurrence in breast cancer patients.33,36–38 Interestingly, dozens of papers have presented compelling evidence for the role of many genes on the 8p11 amplification in breast cancer, including ZNF703,39–41 WHSC1L1,42–44 LSM1, 45 and others. Considering the same location for FGFR1, it is reasonable to assume that FGFR1 may also influence fundamental breast cancer processes. As FGFR1 amplification contributes to carcinogenesis in mammary cells and drives resistance to endocrine therapy, 46 it may be a potential therapeutic target in patients with 8p11–12 amplification.
FGFR2 in breast cancer
In previous studies, GWAS has identified FGFR2 as a breast cancer susceptibility gene and SNPs in the second intron of FGFR2 were associated with an increased risk of developing breast cancer.10,11 The expression and amplification of FGFR2 have long been known to be elevated in 5%–10% of breast tumors. 47 Moreover, it is reported that the “at risk” minor allele increased the risk of developing breast cancer by 1.26-fold in heterozygote and 1.63-fold in homozygote. Although the increase in risk is relatively small, the population carrying at least one copy of the minor allele highly accounts for 40% of risk. 3 Interestingly, the minor allele showed a stronger association with the development of ER-positive than ER-negative breast cancer. 28 In addition, it is reported that the most strongly associated variants were located in intron 2 of the FGFR2 gene,10,11 which shows a high degree of conservation in mammals and contains several putative transcription factor binding sites. Recently, a somatic missense mutation in exon 14 of FGFR2, p.K660N (c.1980G>C) was also revealed to be related to breast tumor development. Further study showed that the p.K660N mutation was primarily mediated by FGFR2-IIIb isoform. 48 Moreover, it remains necessary to discover the concrete mechanisms about the phenotypic outcome of impaired FGF signaling caused by breast cancer mutations in the FGFR2 gene.
Triple-negative breast cancers (TNBCs) have a relatively poor prognosis and lack effective treatment with current targeted therapies. Recently, FGFR2 amplification and overexpression have been implicated in approximately 4% of TNBCs. 49 In addition, both cytoplasmic and nuclear FGFR2 are overexpressed in invasive ductal carcinoma (IDC) compared to normal tissue, and resulted in worse outcomes in terms of overall survival and disease-free survival. 50 Moreover, FGFR2 amplification seems to promote breast tumorigenicity through maintenance of breast tumor-initiating cells. 51 Thus, this tyrosine kinase receptor may be a novel therapeutic target in a subset of TNBCs and IDCs with FGFR2 gene amplification.
FGFR3 in breast cancer
The results from Kuroso et al.’s study have not shown that FGFR3 expression is significantly associated with specific clinicopathological or molecular parameters in invasive breast cancer. However, it demonstrated that elevated levels of FGFR3 were significantly related to poor overall survival in human breast cancer, 52 and we can use FGFR3 as a candidate marker for a poor prognosis. In addition, recent studies also indicated that FGFR3 may be a novel breast cancer susceptibility gene, and FGFR3 may function via a common molecular pathway causing breast cancer. Therefore, FGFR3 was confirmed to be a candidate therapeutic target in FGFR3-associated breast cancer. 53
FGFR4 in breast cancer
FGFR4 G388R is a prevalent germ-line SNP in which glycine is replaced for arginine at codon 388. And the situation occurs in approximately half of the population. Both homozygous and heterozygous carriers of this SNP are overexpressed in a subset of patients with lymph node–positive breast cancer. 54 Although FGFR4 G388R was not reported to increase the incidence of cancer, it has been shown a significant association with poor prognosis, including resistance to chemotherapy and early disease relapse in node-positive breast cancer. 55 In terms of oncogenic function, expression of FGFR4 G388R potentially promotes tumor cell motility and invasion in breast cancer cell lines. 54 Moreover, further studies found that FGFR4 G388R also contributes to breast cancer progression and development of pulmonary metastases in a mouse mammary carcinoma model. 56
FGFRs as a potential therapeutic target for breast cancer
FGFR inhibitors
FGFRs are known to modulate various cellular functions from embryogenesis to adult tissue homeostasis. Moreover, FGFR mutations, gene amplifications, and chromosomal translocations have been identified to be related to the tumor initiation and progression of breast cancer. To date, lots of effective trials have been conducted to explore the rationale and potential roles of FGFR inhibitors in breast cancer (Table 2).
Potential therapeutics target to FGFRs for breast cancer.
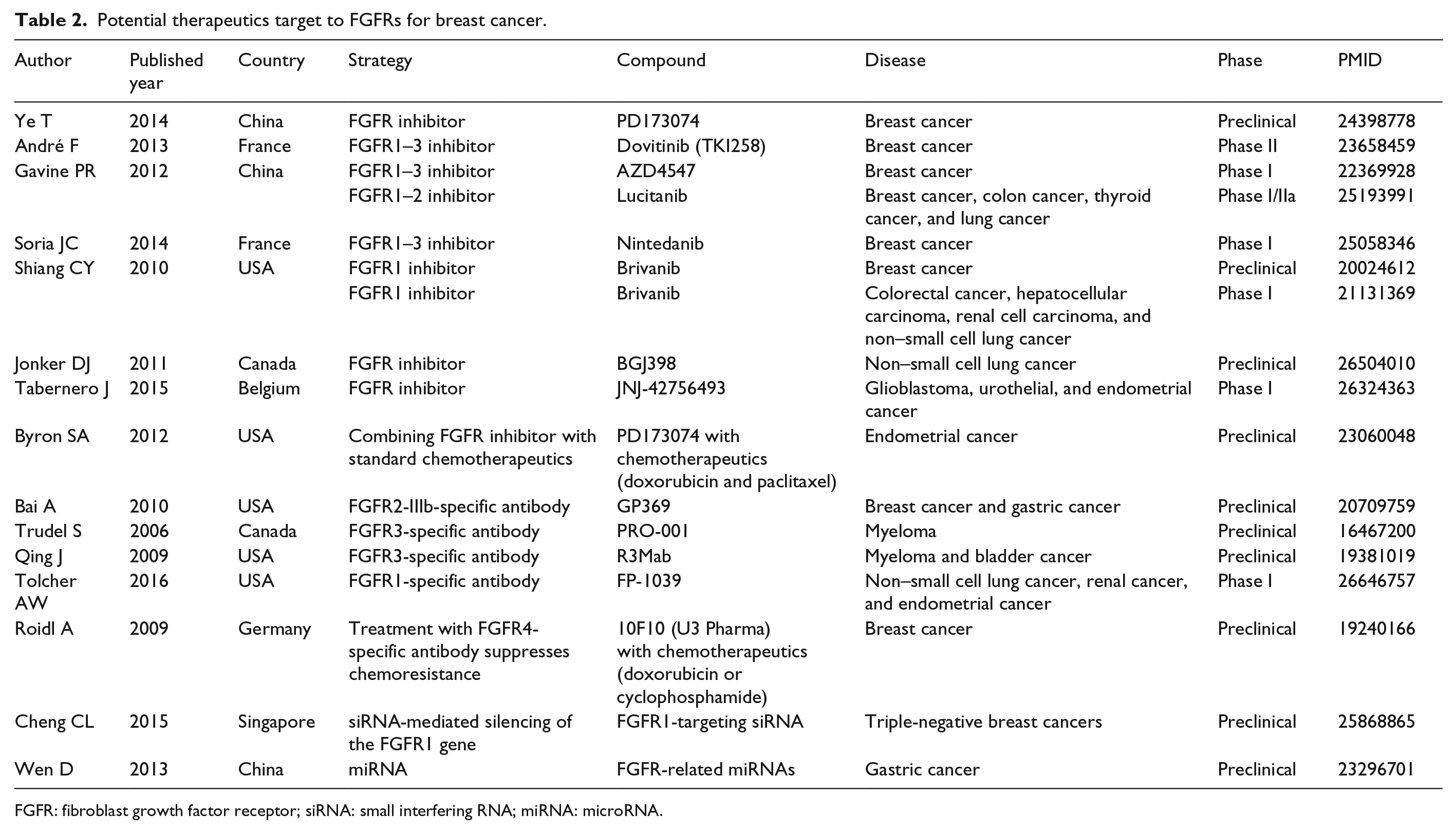
FGFR: fibroblast growth factor receptor; siRNA: small interfering RNA; miRNA: microRNA.
It has been demonstrated that FGFR inhibitor PD173074 inhibited the viability of several human breast cancer cells and 4T1 murine mammary tumor cells. PD173074 also decreased the ability of 4T1 cell migration and invasion in vitro. Moreover, PD173074 significantly delayed breast tumor growth and metastasis to the lung without obvious side effects in 4T1 tumor-bearing mice. 57 So, PD173074 may provide a promising therapeutic approach for the treatment in breast cancer patients.
In addition, other FGFR inhibitors are further tested in clinical trials and most of them are not specific to FGFRs or other kinases. Dovitinib (TKI258) is an oral tyrosine kinase inhibitor (TKI) against FGFR1–3, vascular endothelial growth factor receptor 1–3, and platelet-derived growth factor receptor with IC50 approximately 10 nmol/L in vitro. The results of phase I studies indicated that dovitinib effectively targeted FGFR at safe therapeutic doses. Then, a phase II trial of dovitinib showed modest anti-tumor effects in patients with metastatic breast cancer harboring FGFR1 amplification but not in non-amplified patients. 58 Furthermore, trials of dovitinib in combination with endocrine therapies, as the alternative chemotherapy for patients with metastatic breast cancer, are currently underway (NCT01528345, NCT01262027, NCT01484041). Similarly, AZD4547 is also a well-tolerated oral TKI against both FGFR1–3 and a range of other related kinases, such as kinase insert domain receptor, insulin-like growth factor, PI3Ka, and AKT. Based on the phase I clinical studies, AZD4547 possessed potent anti-tumor activity against FGFR-deregulated tumors, including breast cancer. 59 This agent is further being investigated in patients with ER-positive breast cancer either alone or plus endocrine therapies (NCT01795768, NCT01202591, NCT01791985). Other FGFR TKIs under preclinical or clinical trials were tested in breast cancer patients, for example, lucitanib (NCT02053636), nintedanib (NCT01658462), orantinib (NCT00798252), brivanib (NCT00798252), BGJ398 (NCT01004224, NCT01928459), and JNJ-42756493 (NCT01703481). According to their roles in hindering tumor progression, such as delaying metastasization and decreasing immunosuppression, 2 FGFR inhibitions certainly will be a potential therapeutic target for breast cancer.
Other therapeutics targeted to FGFRs
Human antibody, small interfering RNA (siRNA), and synthetic microRNA (miRNA) appear to be potent methods for the treatment of breast cancer (Table 2). Some research groups proved that FGFR-specific human antibodies functioned as a new therapeutic strategy for the lack of selectivity exhibited by many small molecule FGFR inhibitors. GP369, a FGFR2-IIIb-specific antibody was utilized to treat breast cancer cell line via overexpressing FGFR2. This study exhibited dose-dependent roles of GP369 in decreasing cell proliferation. In vivo studies, treatment with GP369 also inhibited tumor growth in breast cancer xenograft models, and it was well tolerated with no visible signs of toxicity. 60 In addition, many antibodies by modulating FGFRs may display potential therapeutic roles in breast cancer. PRO-001, a selective anti-FGFR3 antibody, showed a significant inhibition of tumor growth in a xenograft model of t(4;14)+ multiple myeloma. 61 Similarly, treatment of both t(4;14)+ multiple myeloma and bladder cancer (containing mutant FGFR3) with another anti-FGFR3 antibody (R3Mab) in mouse xenograft models also significantly restrained tumor growth. 62 Along with more understanding of FGFR3 in breast cancer, PRO-001 and R3Mab may be applied to the treatment of breast cancer in the future. Furthermore, FP-1039 containing the extracellular domain of human FGFR1c, has displayed its potential anti-cancer effects in non–small cell lung cancer, renal cancer, and endometrial cancer cell lines with upregulated FGFRs or FGF ligands. Although FP-1039 was proved to be favorable safety in an initial phase I clinical trial, 63 it needs lot of further studies to investigate whether these antibodies could finally be applied to treat breast cancer in clinic.
The siRNA emerges to be a popular technology for RNA silencing therapy of cancers because of its stability and lower toxicity. Recent studies have identified that siRNA silencing of the FGFR1 gene in MDA-MB-231 TNBC cells significantly inhibited FGFR1 messenger RNA (mRNA) expression, which was closely related to suppression of cell migration. 64 This provided us further evidence that FGFR-targeting siRNA may act as a novel therapeutic method in TNBCs, even more types of breast cancer. Synthetic miRNA was also a potent technology for the treatment of malignancies. Considering the repressive roles of FGFR-related miRNAs in gastric cancer 65 and hepatocellular carcinoma, 66 we hypothesize that these miRNA may also influence FGFRs and inhibit the growth of breast cancer.
Several members of the FGF/FGFR family of signaling molecules have been proved to be associated with chemoresistance in various types of malignancies, including breast cancer. In breast cancer cells, an association has been reported between amplification of FGFR4 with the resistance to doxorubicin. And, the inhibition with a FGFR4 antagonistic antibody was proved to suppress this chemoresistance. 67 In addition, Byron et al.’s 68 study have indicated a potential therapeutic benefit in combining an FGFR inhibitor with standard chemotherapeutic agents in endometrial cancer therapy particularly in patients with FGFR2 mutation positive tumors. Therefore, we also consider that inhibition of FGFRs activation (FGFR inhibitors, antibody, siRNA, and miRNA) will increase the anti-tumor effects when combined with conventional chemotherapeutic agents in breast cancer with FGFRs mutations.
Conclusion and future perspectives
FGFRs are growth factor receptor tyrosine kinases, exerting their roles in embryogenesis, tissue homeostasis, and development of breast cancer. To date, some SNPs in FGFRs have been identified to be significantly associated with breast cancer risk, and these findings ought to be further confirmed in more racial and ethnic groups. In the future, more in-depth genetic screenings should be conducted to find functional variants of FGFRs that are related to breast cancer. According to the biological mechanisms, FGFRs take part in the pathogenesis of breast cancer; several promising FGFR TKIs, specific human antibodies, and related RNA technologies have been developed, and some of them are in early phases of clinical trials. A more comprehensive understanding of FGFRs will helpfully contribute to the development of novel strategies for the prevention and treatment of breast cancer. In addition, as we know, FGFRs have a physiological role in the normal breast and in breast cancer, and it is important to reveal the role of FGFRs in other breast disease, such as hyperplasia of mammary glands or mammitis. However, to date, there are few studies to explore its impact in other breast disease. Hence, this field also needs to be explored in the future.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.