
Abstract
Aberrant expression of microRNAs correlates with the development and progression of human cancers by targeting downstream proteins. MiR-1202 is downregulated in ovarian cancer and clear cell papillary renal cell carcinoma; however, its role in glioma remains unknown. The purpose of this study was to determine the expression and the role of miR-1202 and to elucidate its regulatory mechanism in glioma. We used quantitative real-time polymerase chain reaction to measure miR-1202 expression in both glioma tissues and cell lines. The findings showed that the miR-1202 expression decreased dramatically in clinical glioma tissues and cell lines, and miR-1202 expression was inversely correlated with the expression of Rab1A. Using bioinformatics and luciferase reporter assays, we identified Rab1A as a novel and direct target of miR-1202. In vitro, overexpression of miR-1202 inhibited glioma cell proliferation and induced endoplasmic reticulum stress and apoptosis through targeting Rab1A, whereas suppression of miR-1202 promoted cell proliferation and inhibited endoplasmic reticulum stress and apoptosis. Similarly, silencing Rab1A with small interfering RNA also suppressed glioma cell growth and induced endoplasmic reticulum stress and apoptosis. Taken together, our data indicate that miR-1202 suppresses proliferation and induces endoplasmic reticulum stress and apoptosis through targeting and inhibiting Rab1A in glioma cells. These results suggest miR-1202 as a potential therapeutic target for the treatment of glioma patients.
Introduction
Glioma is the most prevalent and malignant primary tumor in the central nervous system, with a rate of 3–6 per 100,000 population in China.1–3 It is characterized by high proliferative, invasive, and migration abilities. Despite the improvements in therapeutic intervention with combinations of surgery, chemotherapy, radiotherapy, immunotherapy, gene therapy, and other novel biological therapies, the survival rate of patients remains poor. 4 Therefore, it is quite urgent to explore the molecular mechanisms of glioma and develop novel and more effective therapeutic strategies. Increasing evidences have demonstrated that microRNAs (miRNAs) regulate various oncogenes and tumor suppressor genes and participate in the tumorigenesis and progression of glioma.5–8
MiRNAs are small, endogenous, noncoding RNA molecules that are 18–25 nucleotides in length and negatively regulate gene expression at a post-translational level through binding to the 3′-untranslated region (3′-UTR) of target messenger RNAs (mRNAs), leading to the degradation or translational repression of target mRNAs.9–12 Through mediation of their targets’ expression, miRNAs regulate numerous biologic processes such as cell proliferation, cell cycle progression, apoptosis, differentiation, migration, invasion, stress resistance, and cell motility.13–15 Accumulating evidence has revealed that miRNAs were aberrantly expressed in human cancers, including glioma, and they played oncogenic or tumor suppressive roles.16,17 In particular, miR-1202 downregulation has been shown to play a key role in ovarian cancer and clear cell papillary renal cell carcinoma.18,19 We aimed to clarify the expression of miR-1202 in glioma and its involvement in glioma carcinogenesis. Next, we analyzed the miR-1202 targets by bioinformatics software and found that miR-1202 can target Rab1A.
Rab proteins are a family of small GTPases that function in intracellular vesicular trafficking. Rab proteins are localized on specific intracellular compartments and participate in distinct steps in membrane trafficking pathways. 20 They govern membrane transport and regulate cell signaling pathways through binding to effector proteins. As a member of the Rab family, Rab1A functions as a Golgi-resident Rab to vesicle trafficking from endoplasmic reticulum (ER) to Golgi apparatus. 21 Suppression of Rab1A leads to the accumulation of protein within ER, which induces endoplasmic reticulum stress (ERS) signaling pathway referred to unfolded protein response (UPR). UPR coordinates protein synthesis and degradation, cell cycle progression, and apoptosis. 22 If UPR is not effective in decreasing ERS, cell apoptosis is induced.
In this study, we aimed to investigate the biological function of the miR-1202 in glioma cells. We found that miR-1202 was frequently downregulated in glioma tissues. Furthermore, overexpression of miR-1202 significantly inhibited proliferation and induced ERS and apoptosis of glioma cells. Bioinformatics analysis and dual luciferase reporter assays demonstrated that Rab1A is a novel target of miR-1202. Knockdown of Rab1A by shRab1A induced ERS and apoptosis. In addition, shRab1A suppresses the growth of glioma U87 cells. Overall, our data suggest that miR-1202 inhibits proliferation and induced ERS and apoptosis of glioma cells by directly targeting Rab1A.
Materials and methods
Human tissue samples
A total of 39 human glioma samples were obtained from adult patients who were admitted and diagnosed at Second Affiliated Hospital at Xi’an Jiaotong University. Nine normal brain samples were collected from people with traumatic brain injury, for whom a partial resection of brain tissues was conducted for decreasing the intracranial pressure. The samples were immediately frozen in liquid nitrogen and stored at −80°C. Informed consent was obtained from each patient before specimen collection. This study was approved by the Ethics Committee of Xi’an Jiaotong University Health Science Center.
Cell lines and cell culture
The human glioma U87, U251, U373, A172, and LN229 cell lines were obtained from the Environment and Genes Related to Diseases Key Laboratory of Education Ministry, Xi’an Jiaotong University Health Science Center, China. Primary normal human astrocytes (NHAs) were purchased from the Cell Bank (Shanghai Genechem Co., Ltd., Shanghai, China). The cells were cultured in RPMI-l640 medium (Invitrogen, Carlsbad, CA, USA) supplemented with 10% (v/v) fetal bovine serum (Gibco BRL, Grand Island, NY, USA) containing 50 U/mL penicillin, 50 µg/mL streptomycin, and 2.0 mmol/L glutamine and were incubated at 37°C with 5% CO2.
Quantitative real-time polymerase chain reaction analysis
Total RNA was extracted from tissues or cell lines using TRIzol reagent (Invitrogen, USA) according to the manufacturer’s instructions. RNA was reverse transcribed to complementary DNA (cDNA) with a Reverse Transcription Kit (Takara, Dalian, China). Quantitative real-time polymerase chain reaction (qRT-PCR) analysis was carried out using Power SYBR Green PCRmastermix (Takara, China). qRT-PCR was conducted using the iCycler iQ Multicolor qRT-PCR Detection System (Bio-Rad, Hercules, CA, USA). The procedures were performed according to the manufacturers’ instructions. The results were normalized to U6 or β-actin expression. The primer sequences were as follows: miR-1202 reverse-transcribed primer, 5′-GTCGTATCCAGTGCGTGTCGTGGAGTCGGCAATTGCACTGGATACGACGGGGGAG-3′; miR-1202 forward, 5′-ATCCAGTGCGTGT CGTG-3′; miR-1202 reverse, 5′-TGCTGTGCCAGCTGCAGT-3′; U6 reverse-transcribed primer, 5′-CGCTTCACGAATTTGCGTGTCAT-3′; U6 forward, 5′-GCTTCGGCAGCACATATACTAAAAT-3′; U6 reverse, 5′-CGCTTCACGAATT TGCGTGTCAT-3′; Rab1A forward, 5′-GGGAAAACAATCAAGCTTCAAA-3′; Rab1A reverse, 5′-CTGGAGGTGATTGTTCGAAAT-3′; β-Actin forward, 5′-TGGCACCCAGCACAATGAA-3′; β-Actin reverse, 5′-CTAAGTCATAGTCCGCCTAGAAGC A-3′.
Immunohistochemistry
Immunohistochemical study was performed on paraffin-embedded glioma tissues using a mouse monoclonal anti-Rab1A antibody (1:100; Santa Cruz, CA, USA). The tissues were sectioned at 4 µm and prepared on glass slides. The slides were deparaffinized according to established guidelines and quenched with 3% hydrogen peroxide for 5 min. They were rinsed in distilled water followed by Tris-buffered saline. Then, heat-induced antigen retrieval was performed with citrate buffer for 30 min after cooling. The sections were incubated with mouse anti-Rab1A antibody for 4 h. All sections were processed based on the peroxidase–antiperoxidase method (Dako, Hamburg, Germany). The peroxidase reaction was visualized through incubating the sections with 3,3′-diaminobenzidine (DAB). The sections were counterstained with hematoxylin solution, examined through light microscopy. Two independent pathologists scored Rab1A protein immunostaining based on staining intensity and positive cells’ percentage. Rab1A expression <5% was considered negative, and ≥5% was considered positive (overexpression).
Luciferase reporter assay
The 3′-UTR of human Rab1A mRNA was constructed with synthetic oligonucleotides and cloned in between the XhoI and SacI sites of the pmirGLO Dual-Luciferase miRNA target expression vector (Promega, Madison, WI, USA). PmirGLO-Rab1A-3′-UTR vector was co-transfected with miR-1202 into the HEK293T cell line, with pmirGlO vector as their control. Then, the cells were harvested for luciferase assays 24 h after transfection. Reporter activity was examined with the Dual-Luciferase Assay System (Promega).
Plasmid constructs, miR-1202 inhibitor synthesis, and transfection
Hsa-miR-1202 precursor was interpolated into pcDNA6.2-GW/EmGFPmiR vector according to the manufacturer’s instructions (named miR-1202). Scramble miRNA was interpolated into the same vector and used as negative control (named miR-1202-Ctrl). The sequence of negative control miRNA is 5′-AGGTACGAAACGCTAAGAAT-3′. Cell transfection was performed with Lipofectamine 2000 (Invitrogen) according to the manufacturer’s instructions. Small interfering RNA (siRNA) oligonucleotides were used as miR-1202 inhibitors (named anti-miR-1202) and purchased from GenePharma (Shanghai, China). The sequence of anti-miR-1202 was 5′-CUCCCCCACUGCAGCUGGCAC-3′. Scramble siRNA was used as negative control (named anti-miR-1202-Ctrl), and the sequence was 5′-CAGUACUUUUGUGUAGUACAA-3′. siRNA oligonucleotides were transfected into glioma U87 cells with Lipofectamine 2000 according to the manufacturer’s guidelines.
siRNA synthesis and transfection
siRNA was used for Rab1A gene silencing. Human Rab1A siRNA (sense 5′-GGAAACCAGUGCUAAGAAUT-3′, antisense 5′-AUUCUUAGCACUGGUUUCCTT-3′) and negative siRNA (NC-siRNA, sense 5′-UUCUCCGAACGUGUCACGUTT-3′, antisense 5′-ACGUGACACGUUCGGAGAATT-3′) were pre-designed and chemically synthesized by GenePharma Corporation (Shanghai, China). Lipofectamine 2000 was used to optimize siRNA transfection. siRNA and Lipofectamine 2000 were initially formed with 250 nM siRNA and then diluted to 60 nM in the plated cells in future experimental procedures.
Cell proliferation assay
U87 cells (5000 cells/well) were seeded into 96-well plates with 200 µL RPMI-l640 medium in each well and incubated for 1 day. The cells were then treated with miR-1202-Ctrl vector, miR-1202 vector, anti-miR-1202-Ctrl, anti-miR-1202, NC-siRNA (60 nM), or Rab1A siRNA (60 nM) for 1, 2, and 3 days, respectively. Cell activity was measured at a wavelength of 490 nm with the MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide) assay FLUOstar OPTIMA (BMG Labtech, Ortenberg, Germany). Each experiment contained five replicates, and the results were collected as the mean of three independent experiments.
Cell cycle analysis
Cells at 3 × 105 cells per well were cultured in six-well plates for 1 day and treated by miR-1202-Ctrl, miR-1202, anti-miR-1202-Ctrl, anti-miR-1202, NC-siRNA, or Rab1A siRNA for 2 days. At the end of culture, cells were harvested by trypsinization and fixed in 75% icecold ethanol overnight at 4°C. The cells were then washed with phosphate-buffered saline (PBS) and stained with 50 µg/mL propidium iodide (PI) containing 50 µg/mL RNase A for 20 min at room temperature. The cells were examined by fluorescence-activated cell sorting (FACSCalibur; BD Biosciences, San Jose, CA, USA), and the cell cycle populations were analyzed by ModFit software. In each independent experiment, three parallel wells were made, and the experiments were carried out in triplicate.
Cell apoptosis analysis
Cells were seeded into six-well plates at a density of 3 × 105 cells per well for 1 day and treated by miR-1202-Ctrl, miR-1202, anti-miR-1202-Ctrl, anti-miR-1202, NC-siRNA, or Rab1A siRNA for 2 days. Then, the cells were harvested and washed twice with PBS. Cell apoptosis analysis was performed with Annexin V FITC Apoptosis Detection Kit (Invitrogen) according to the manufacturer’s instructions and examined using a flow cytometer. The apoptosis populations were analyzed by ModFit software.
Western blot analysis
Total protein was extracted using radioimmunoprecipitation assay (RIPA) lysis buffer from cells harvested 48 h after transfection or tissue samples. Protein was separated in 10% sodium dodecyl sulfate (SDS) polyacrylamide gels and transferred to a nitrocellulose membrane. Membranes were blocked for 1 h in 5% non-fat dry milk in TBST (10 mM Tris–HCl and 0.05% Tween 20). Next, the membranes were incubated with primary antibodies overnight at 4°C and with secondary antibody for 1 h at room temperature. The primary antibodies included mouse monoclonal anti-Rab1A (1:1000, Santa Cruz Biotechnology), mouse monoclonal anti-Bcl-2 (1:1000, Santa Cruz Biotechnology), mouse monoclonal anti-Bax (1:1000, Santa Cruz Biotechnology), rabbit polyclonal anti-GRP78 (1:1000, Santa Cruz Biotechnology), and mouse monoclonal anti-β-actin (1:5000, Santa Cruz Biotechnology). The membranes were incubated with enhanced chemiluminescence (ECL) (Amersham, Little Chalfont, Buckinghamshire, UK) for chemiluminescence detection. The blots were scanned and the band density was measured with Syngene GBox (Syngene, Cambridge, UK). The experiments were carried out in triplicate.
Statistical analysis
Each experiment was repeated at least three times independently. The quantitative data were presented as mean ± standard error of mean (SEM). All statistical analyses were performed using SPSS14.0 software. Student’s t test and Pearson’s chi-square test were used according to the data characteristics. Survival curves by Rab1A status were estimated using the Kaplan–Meier and compared by log-rank test. Differences were considered statistically significant at p < 0.05.
Results
MiR-1202 is downregulated and is reciprocally related to the overexpressed Rab1A in glioma tissues
To analyze the expression levels of miR-1202 in glioma, qRT-PCR was conducted in nine normal brain tissues and 39 glioma samples. Results showed that the expression of miR-1202 was lower in glioma samples than normal brain tissues (p < 0.01) and decreased with the increasing degree of malignancy in glioma (low-grade glioma vs high-grade glioma, p < 0.05; Figure 1(a)). Additionally, miR-1202 expression levels in five glioma cell lines (U87, U251, U373, A172, and LN229) were also decreased compared with the NHAs (Figure 1(b)). Since miR-1202 expression in U87 cells was the lowest, this cell line was chose to be used in the following experiments. The expression of Rab1A mRNA in glioma samples was higher compared with normal brain tissues (p < 0.01) and increased with the increasing degree of malignancy in glioma (low-grade glioma vs high-grade glioma, p < 0.01; Figure 1(c)). The effect of miR-1202 on Rab1A was assessed based on the data obtained from qRT-PCR. A significant inverse correlation was identified between Rab1A and miR-1202 (Figure 1(d); n = 39, r = −0.893, p < 0.001, Pearson’s correlation). A dramatic increase in Rab1A protein was found in glioma samples as compared with normal brain tissues and increased with the increasing degree of malignancy (Figure 1(e) and (f); p < 0.01). In addition, the overall survival status was statistically different between Rab1A overexpression group and Rab1A-negative group (Figure 1(g); p < 0.01).
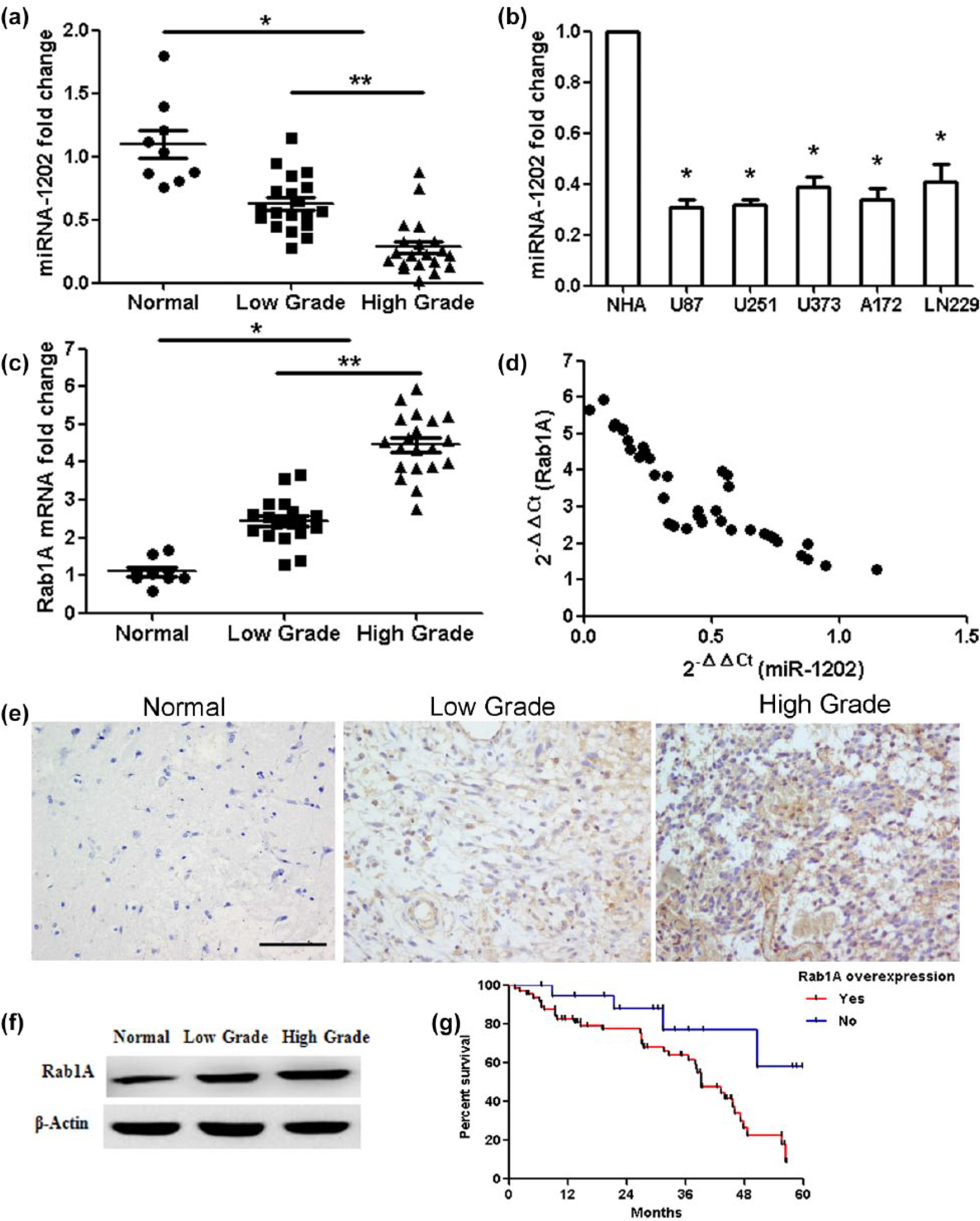
Expressions of miR-1202 and Rab1A, and the correlation between miR-1202 and Rab1A in glioma. (a) MiR-1202 expression in glioma samples compared with normal brain tissues (*p < 0.01), miR-1202 expression with the increasing degree of glioma (low grade vs high grade; **p < 0.05). (b) MiR-1202 expression was analyzed in glioma U87, U251, U373, A172, LN229, and normal human astrocyte (NHA). (c) Rab1A expression in glioma samples compared with normal brain tissues (*p < 0.01), Rab1A expression with the increasing degree of glioma (low grade vs high grade; **p < 0.01). (d) miR-1202 and Rab1A levels were inversely correlated. 2−ΔΔCt values of miR-1202 and Rab1A were subjected to Pearson’s correlation analysis (n = 39, r = −0.893, p < 0.001, Pearson’s correlation). (e) Immunohistochemistry analysis showed that Rab1A protein was positively expressed in the low-grade and high-grade glioma tissues but was negative in normal brain tissues. Bar = 100 µm. (f) Rab1A protein level was measured by western blotting. (g) The 5-year overall survival curves depending on Rab1A overexpression (blue line: Rab1A overexpression, red line: negative Rab1A expression; p < 0.05).
Rab1A is a direct target of miR-1202 in glioma cells
Rab1A was predicted as one of the candidate target genes of miR-1202 by three algorithms (PicTar, miRBase Target, and TargetScan). The Rab1A-encoded mRNA contains a 3′-UTR element ranging from dinucleotide 2261 to 2281 bp, which is stringently complementary to the seed sequence of miR-1202 (Figure 2(a)). This indicates that miR-1202 would directly target Rab1A. The wild-type Rab1A-3′-UTR construct in combination with miR-1202 or control was transfected into HEK293T cells. Compared with control, miR-1202 led to a significant decrease in luciferase activity of the Rab1A-3′-UTR construct (p < 0.01). After the conserved targeting regions for miR-1202 recognition were mutated, the luciferase activity of the reporter gene was restored (Figure 2(b)). The regulatory function of miR-1202 on Rab1A was determined at both mRNA and protein levels. The results showed that the expression of miR-1202 in U87 cells transfected with miR-1202 overexpression was upregulated and compared with cells transfected with the control (Figure 2(c); p < 0.01). Overexpression of miR-1202 significantly downregulated Rab1A mRNA and protein levels in U87 cell lines. Consistently, inhibition of miR-1202 led to an increased expression of Rab1A at both mRNA and protein levels (Figure 2(d) and (e); p < 0.01). These data demonstrated that Rab1A is a target gene of miR-1202 in glioma cells.

MiR-1202 directly targets the Rab1A gene. (a) Bioinformatics predicted interactions of miR-1202 and their binding sites at the 3′-UTR of Rab1A. (b) The luciferase reporter plasmid–containing mutant or wild-type Rab1A-3′-UTR was co-transfected into HEK293T cells in combination with miR-1202 or control. Luciferase activity was determined by the dual luciferase assay. (c) miR-1202 expression was determined after miR-1202 overexpression or anti-miR-1202 treatment. (d) The level of Rab1A mRNA was examined after miR-1202 overexpression or anti-miR-1202 treatment. (e) The expression of Rab1A protein was analyzed by western blotting. β-Actin was used as control. *p < 0.01, compared with miR-1202-Ctrl group; p < 0.01, compared with anti-miR-1202-Ctrl group, n = 3.
Overexpression of miR-1202 inhibits proliferation and induces ERS and apoptosis in glioma U87 cells by targeting Rab1A
As miR-1202 was expressed at a lower level in glioma samples compared with normal brain tissues, we hypothesized that miR-1202 might act as a tumor suppressor and that restoration of miR-1202 expression could regulate cell proliferation and apoptosis in glioma cells. MiR-1202 vector or miR-1202-Ctrl vector were transfected into glioma U87 cells. MTT assay was used to measure cell activity. We found that the proliferation of cells transfected with miR-1202 vector was decreased significantly compared with that of cells transfected with miR-1202-Ctrl vector (Figure 3(a); p < 0.01). Since the cell cycles are involved in the regulation of cell proliferation, we examined the processes using a flow cytometer 2 days after transfection. The results showed that cell cycles were arrested significantly at G1/G0 phase in miR-1202 group (Figure 3(b); p < 0.01). In addition, cell apoptosis analysis showed that the proportion of early apoptosis and late apoptosis increased remarkably in miR-1202 group (Figure 3(c); p < 0.01). To explore whether miR-1202-dependent Rab1A inhibition activates ERS, we examined the expression of glucose-regulated protein 78 (GRP78), which belongs to ER molecular chaperones and is considered a marker of ERS. The protein expression of GRP78 significantly increased in cells transfected with miR-1202 vector. The protein expressions of Rab1A and Bcl-2 were lower in miR-1202 group compared with miR-1202-Ctrl group, but the expression of Bax was higher (Figure 3(d)). These data suggest that miR-1202 targets and suppresses Rab1A, induces ERS, and results in apoptosis in glioma cells.

MiR-1202 overexpression inhibits proliferation and induces apoptosis in glioma U87 cells. (a) MTT assay showed that miR-1202 overexpression decreased the cell activity at 2 and 3 days. (b) The flow cytometry analysis of cell cycle was visualized via PI staining. The data show the percentages in the G1/G0, S, and G2 phases. (c) The flow cytometry analysis of apoptosis was visualized using Annexin V/PI staining. The data showed the percentages of early apoptosis and late apoptosis. (d) The protein expressions of Rab1A, GRP78, Bcl-2, and Bax were examined by western blot in miR-1202 group. *p < 0.01, compared with miR-1202-Ctrl group, n = 3.
Inhibition of miR-1202 promotes proliferation and suppresses ERS and apoptosis in glioma U87 cells by targeting Rab1A
Human glioma U87 cells were transfected with miR-1202 antisense (anti-miR-1202). MTT assay showed that anti-miR-1202 promoted the proliferation of glioma cells (Figure 4(a); p < 0.01). The number of the S-phase cells significantly increased in the anti-miR-1202 group compared with anti-miR-1202-Ctrl group; meanwhile, the number of the G1/G0 phases cells remarkably decreased (Figure 4(b); p < 0.01). There was no significant difference among the proportion of early apoptosis and late apoptosis in different treatment groups (Figure 4(c)). Rab1A and Bcl-2 protein expressions increased in cells transfected with anti-miR-1202, while GRP78 and Bax protein expressions decreased (Figure 4(d)).
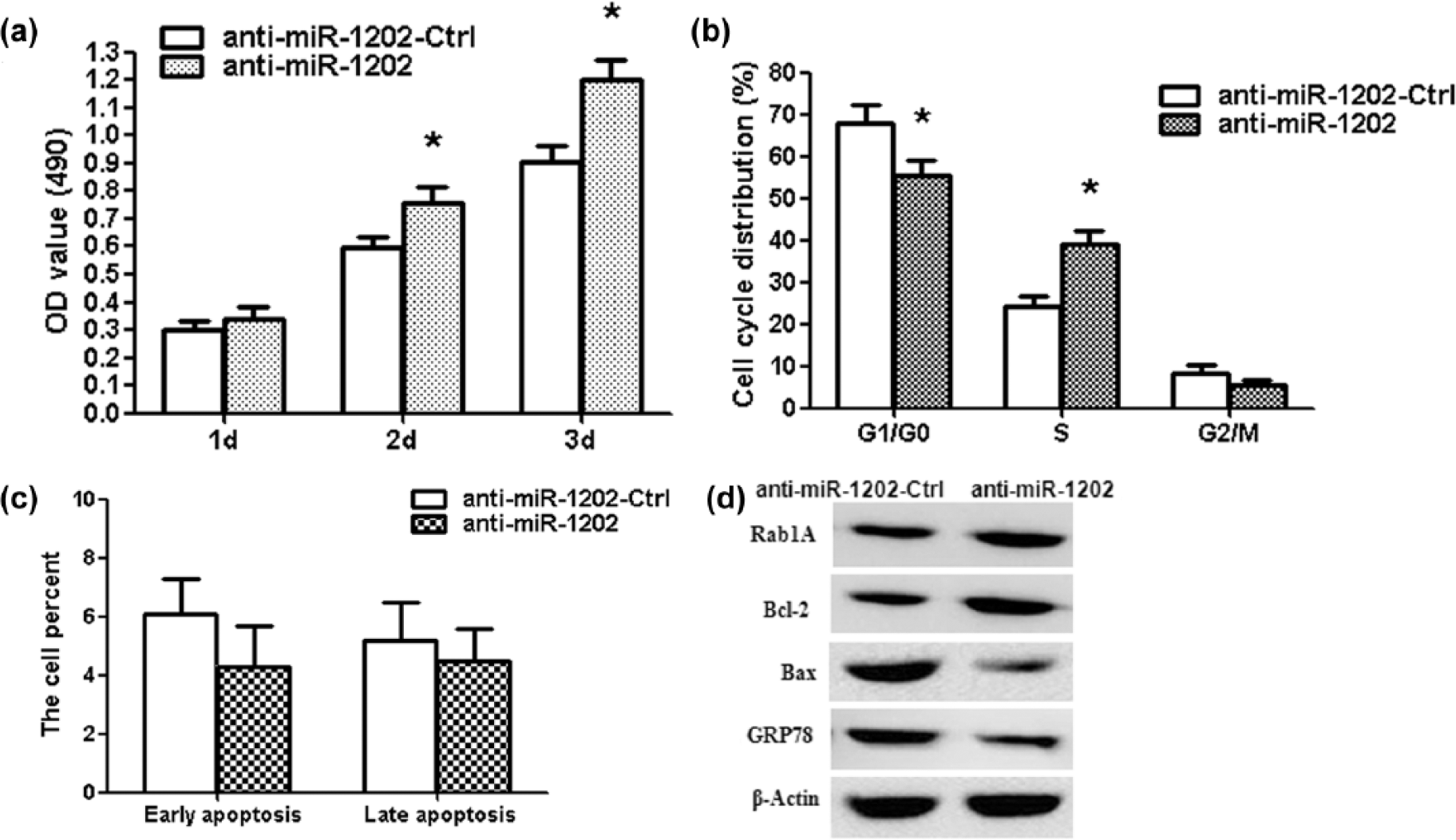
Inhibition of miR-1202 promotes cell proliferation and suppresses apoptosis in glioma U87 cells. (a) MTT assay showed that anti-miR-1202 increased the cell activity at 2 and 3 days. (b) The flow cytometry analysis showed the percentage in the G1/G0, S, and G2/M phases. S-phase cells significantly increased after anti-miR-1202 treatment. (c) The data showed the percentage of early and late apoptosis. (d) Rab1A, GRP78, Bcl-2, and Bax protein expressions were measured after anti-miR-1202 treatment. *p < 0.01, compared with anti-miR-1202-Ctrl group, n = 3.
Knockdown of Rab1A inhibits glioma U87 cell growth and induces ERS and apoptosis
To investigate the functional roles of Rab1A in glioma cells, we knocked down Rab1A in U87 cells by Rab1A-specific small interfering RNAs (Rab1A siRNAs). The results showed that Rab1A siRNA significantly decreased the cell activity compared to NC-siRNA group (Figure 5(a); p < 0.01). The cell cycles were arrested significantly at G1/G0 phases in the siRNA group. The number of S-phase cells significantly decreased in the siRNA group (Figure 5(b); p < 0.01). The proportion of early apoptosis and late apoptosis increased significantly (Figure 5(c); p < 0.01). These were similar to the variations that resulted from miR-1202 overexpression, indicating a similar effect of Rab1A knockdown and miR-1202 overexpression. Furthermore, we analyzed knockdown efficiency of Rab1A siRNA in mRNA and protein levels. The Rab1A mRNA and protein expression decreased significantly in siRNA group compared to NC-siRNA group; Bcl-2 protein expression also decreased, but GRP78 and Bax protein expressions increased (Figure 5(d) and (e); p < 0.01).
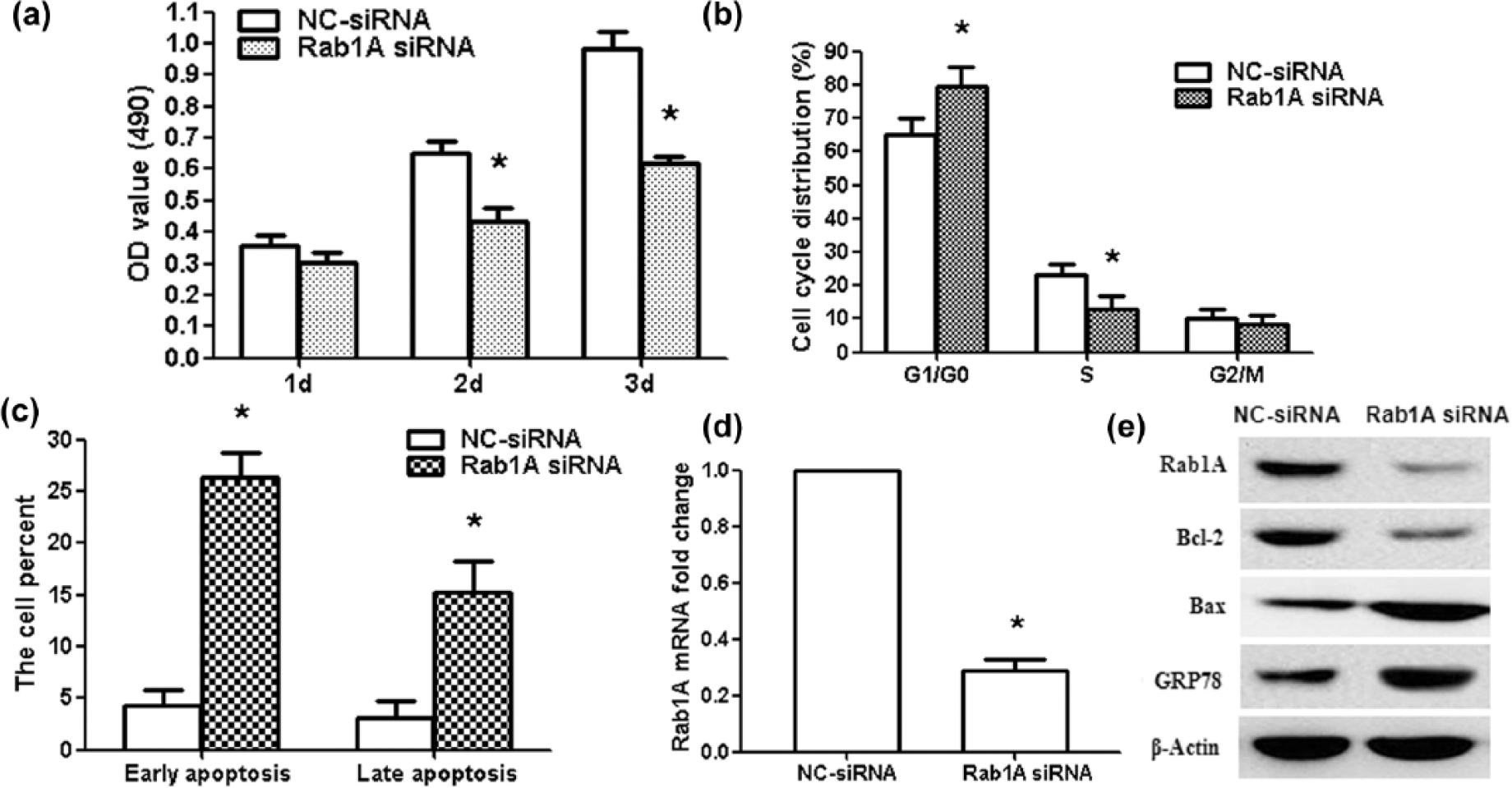
Rab1A siRNA suppresses proliferation and induces apoptosis in glioma U87 cells. (a) MTT assay showed that Rab1A siRNA decreased the activity of U87 cells at 2 and 3 days. (b) The results of flow cytometry analysis showed the percentage of cells in the G1/G0, S, and G2/M phases. G1/G0 phase cells increased significantly after Rab1A siRNA treatment and S-phase cells decreased after Rab1A siRNA treatment. (c) The data showed the percentage of early apoptosis and late apoptosis after Rab1A siRNA treatment. (d) qRT-PCR results showed the knockdown efficiency of Rab1A siRNA in U87 cells. (e) Rab1A, GRP78, Bcl-2, and Bax protein expressions were measured after Rab1A siRNA treatment. *p < 0.01, compared with NC-siRNA group, n = 3.
Discussion
Tumor progression is a highly orchestrated process that requires complex transcription and posttranscriptional regulation of gene expression.23–25 Growing evidence has revealed that miRNAs play an important role in carcinogenesis and cancer progression. Depending on their mRNA targets, miRNAs can act as oncogenes or tumor suppressor genes.26,27 The changes of miRNA are involved in most aspects of cancer biology, including proliferation, apoptosis, migration, and invasion.28–30 Therefore, miRNAs are usually viewed as a potential diagnostic and therapeutic tool.31,32 Now, only a small proportion of miRNAs have been investigated to elucidate their precise biological roles in glioma. Identifying miRNAs specifically involved in glioma carcinogenesis will aid in developing new targets for diagnosis and therapy.
In this study, we provide the first evidence that the expression of miR-1202 is significantly downregulated in glioma tissues. This is similar to that in ovarian cancer and clear cell papillary renal cell carcinoma as reported by researchers.18,19 The aim of this work is to investigate the biological functions of miR-1202 in glioma. Our findings demonstrate that overexpression of miR-1202 significantly inhibited glioma cell proliferation and induced cell apoptosis in vitro. However, suppression of miR-1202 remarkably promoted glioma cell proliferation in vitro. We explored potential targets of miR-1202. Rab1A was postulated to be a target of miR-1202 using three different databases. Our results showed that a significant inverse correlation was identified between Rab1A and miR-1202. As indicated on luciferase reporter assays, miR-1202 suppressed the construct with PmirGLO-Rab1A-3′-UTR. Overexpression of miR-1202 inhibited the expression of Rab1A mRNA and protein. MiR-1202 overexpression also repressed Rab1A-3′-UTR luciferase report activity, and this effect was abolished by mutation of the miR-1202 seed binding site. The results indicate that miR-1202 may play the role of a negative regulator or tumor suppressor for proliferation and apoptosis partly mediated through inhibiting Rab1A expression.
The Rab GTPase protein family, including more than 60 Rab proteins, localizes to specific intracellular membranes and plays a key role in membrane trafficking of the Golgi, ER, lysosomes, phagosomes, endosomes, and autophagosomes. 33 Recent evidences indicate that Rab1A is an oncogene in hepatocellular carcinoma, prostate cancer, colorectal cancer, and tongue cancer.34–36 The last two studies show that Rab1A promotes hepatocellular carcinoma cell proliferation.34,37 Our results show that miR-1202 and Rab1A siRNA inhibited glioma cell proliferation, and this indicates that miR-1202 inhibited glioma cell proliferation by downregulating target Rab1A expression.
Rab1A is located at ER exit sites and pre-Golgi intermediate compartment. Rab1A can regulate ER-Golgi trafficking in the late stages of ER-to-Golgi transport through promoting the fusion of COPII-coated vesicles with the cis face of the Golgi. ER plays an important role in the synthesis, folding, and structural maturation of many proteins made. Genetic and environmental factors influence proper folding and post-translationally modify secretory and transmembrane proteins in the ER. 38 Damaged or misfolded proteins accumulated in this organelle can exceed the capacity of the ER to fold them and lead to ERS. Cells quickly restore protein-folding capacity in order to reduce the accumulation of misfolded proteins and activate UPR under ERS. When ERS persists at high levels, adaptive mechanisms are useless, ERS is irremediable, and UPR transforms into an alternate signaling platform called terminal UPR, promoting cell death. ERS interferes with the suppressive effects of GRP78, which is the ER chaperone. When unfolded proteins and chaperones are imbalanced, transmembrane proteins ATF6, PERK, and IRE1 are activated through dissociation with GRP78. 39 UPR can transit into apoptosis with high expression of CHOP/GADD153 transcription factor. CHOP/GADD153 induces cell death and facilitates the expression of pro-apoptotic factors through inhibiting the expression of anti-apoptotic Bcl-2. Our results show that miR-1202 and Rab1A-siRNA induce ERS and apoptosis in glioma cells.
In conclusion, this study showed that downregulation of miR-1202 and upregulation of Rab1A are observed in human glioma tissues. The inverse correlation between miR-1202 and Rab1A expression in glioma tissues, luciferase assays results, and the downregulation of Rab1A in response to overexpression of miR-1202 in glioma cells identified Rab1A as a target of miR-1202 in glioma. Here, for the first time, we demonstrate that miR-1202 suppresses proliferation and induces ERS and apoptosis through targeting Rab1A in glioma cells. These results provide evidence that miR-1202 could be a useful marker in glioma and a potential therapeutic target for the treatment of glioma patients.
Footnotes
Acknowledgements
The authors would like to thank Professor Xiao Han for polishing the language in this manuscript.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was supported by the Wu Jieping Foundation for Clinical Scientific Research (320675014057).