
Abstract
Nowadays, although chemotherapy is an established therapy for breast cancer, the molecular mechanisms of chemotherapy resistance in breast cancer remain poorly understood. This study aims to explore the effects of endoplasmic reticulum stress on autophagy, apoptosis, and chemotherapy resistance in human breast cancer cells by regulating PI3K/AKT/mTOR signaling pathway. 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay was performed to detect the cell viability of six human breast cancer cell lines (MCF-7, ZR-75-30, T47D, MDA-MB-435s, MDA-MB-453, and MDA-MB-231) treated with tunicamycin (5 µM), after which MCF-7 cells were selected for further experiment. Then, MCF-7 cells were divided into the control (without any treatment), tunicamycin (8 µ), BEZ235 (5 µ), and tunicamycin + BEZ235 groups. Cell viability of each group was testified by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay. Western blotting was applied to determine the expressions of endoplasmic reticulum stress and PI3K/AKT/mTOR pathway–related proteins and autophagy- and apoptosis-related proteins. Monodansylcadaverine and Annexin V–fluorescein isothiocyanate/propidium iodide staining were used for determination of cell autophagy and apoptosis. Furthermore, MCF-7 cells were divided into the control (without any treatment), tunicamycin (5 µM), cisplatin (16 µM), cisplatin (16 µM) + BEZ235 (5 µM), tunicamycin (5 µM) + cisplatin (16 µM), and tunicamycin (5 µM) + cisplatin (16 µM) + BEZ235 groups. Cell viability and apoptosis were also evaluated by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay and Annexin V–fluorescein isothiocyanate/propidium iodide staining. In MCF-7 cells treated with tunicamycin, cell viability decreased significantly, but PEAK, eIF2, and CHOP were upregulated markedly and p-PI3K, p-AKT, and p-MTOR were downregulated in dose- and time-dependent manners. In the tunicamycin + BEZ235 group, the cell viability was lower and the apoptosis rate was higher than those of the control and monotherapy groups. Compared with the cisplatin group, the tunicamycin + cisplatin group showed a relatively higher growth inhibition rate; the growth inhibition rate substantially increased in the tunicamycin + cisplatin + BEZ235 group than the tunicamycin + cisplatin group. The apoptosis rate was highest in tunicamycin + cisplatin + BEZ235 group, followed by tunicamycin + cisplatin group and then cisplatin group. Our study provide evidence that endoplasmic reticulum stress activated by tunicamycin could promote breast cancer cell autophagy and apoptosis and enhance chemosensitivity of MCF-7 cells by inhibiting the PI3K/AKT/mTOR signaling pathway.
Keywords
Introduction
Breast cancer (BC) is the most frequent cancer which occurs in women with 1.7 million newly diagnosed cancer cases worldwide in 2012 and is one of the most common causes of cancer death among women. 1 As a devastating disease in both developed and developing countries, BC is the most prevalent cancer in Chinese women as well and accounts for approximately 12.2% of newly diagnosed BC cases and 9.6% of BC deaths worldwide. 2 Although the etiology of BC has not been clearly explained yet, several factors may be responsible for the development of BC, including external factors like radiation exposure, hormone replacement therapy, and lifestyle as well as internal factors such as age, reproduction, family history of breast disease, and genetic predisposition.3,4 Currently, BC therapy traditionally consists of surgery, radiotherapy, systemic chemotherapy, hormone therapy, and molecular targeted therapy, which may result in a series of complications and chemotherapy resistance in BC patients. 5 Thus, it is necessary to explore the molecular mechanisms underlying chemotherapy resistance for developing new cancer therapies.
Autophagy refers to a stress-activated catabolic process that can promote the degradation of damaged organelles and mis-folded/aggregated proteins in cancer cells during clinical therapies, and thus, autophagy has emerged as a novel strategy to overcome drug resistance during cancer therapies. 6 It was reported that enhanced autophagy induced cell deaths in doxorubicin-resistant osteosarcoma cells 7 and increased radiosensitization in apoptosis-deficient lung cancer cells, 8 supporting autophagy modulation in overcoming cancer drug resistance. According to the previous study, cancer cells are usually under endoplasmic reticulum stress (ERS) and they are in a need of a highly specific ER quality-control system which is also named as the unfolded protein response (UPR) to maintain ER homeostasis. 9 In recent years, it has been found that ERS could directly induce autophagy for protein restoration by synthesizing unfolded, misfolded, and damaged proteins and subsequently inducing UPR-mediated upregulation of binding immunoglobulin protein (BiP), which is the dispensable autophagosome formation. 10 Interestingly, the cells’ ability of responding to ERS is essential for their survivals, and chronic or unresolved ERS can result in cell apoptosis. 11 Thus, both ERS and autophagy induction might contribute to the cytotoxicity of anti-cancer drugs on various cancers. However, overactivation of phosphatidylinositol 3-kinase (PI3K)/AKT/mammalian target of rapamycin (mTOR) signaling pathway was also linked with carcinogenesis and negative regulation of the PI3K/AKT/mTOR signaling pathway induced apoptosis in BC cells. 12 Moreover, it was reported that ERS inducers such as tunicamycin increased cell death via inhibition of the PI3K/AKT/mTOR signaling pathway in melanoma cells. 13 Therefore, the hypotheses were proposed that tunicamycin-induced ERS may regulate autophagy, apoptosis, and chemotherapy resistance of human BC MCF-7 cells through the PI3K/AKT/mTOR signaling pathway, so as to provide evidence for the development of BC-targeted therapy.
Material and methods
Cell culture
The MCF-7, ZR-75-30, T47D, MDA-MB-435s, MDA-MB-453, and MDA-MB-231 cells (obtained from the Chinese Academy of Sciences, Shanghai, China) were maintained at 37°C in a humidified atmosphere containing 5% CO2 in Dulbecco’s modified Eagle’s medium (DMEM; pH = 7.2; Gibco Company, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (FBS; Gibco Company). When cells grew to 80%–90% confluence, the cells were digested with 0.25% trypsin containing ethylenediaminetetraacetic acid (EDTA; Sigma-Aldrich Chemical Company, St. Louis, MO, USA), seeded in a 6-well plate and a 12-well plate at the ratio of 1:3. The DMEM containing 1% FBS was adopted in the digestion and seeding of cells. The drugs were not added until all cells adhered to wall.
Cell grouping and dosage regimens
The MCF-7 cells were treated with tunicamycin at the concentrations of 0, 0.5, 1, 2, 4, and 8 µM for 12, 24, and 48 h and subsequently used for determination of cell viability as well as protein expressions of ERS pathway and the PI3K/AKT/mTOR signaling pathway.
To detect the survival rate, autophagy, and apoptosis of MCF-7 cells when the PI3K/AKT/mTOR signaling pathway was inhibited, the MCF-7 cells were divided into the control (without any treatment), tunicamycin (8 µM), BEZ235 (5 µM), and tunicamycin + BEZ235 groups.
For the detection of cell proliferation and apoptosis, the MCF-7 cells were divided into the control (without any treatment), tunicamycin, cisplatin (16 µM, treated with cisplatin for 24 h), cisplatin + BEZ235, tunicamycin + cisplatin, and tunicamycin + cisplatin + BEZ235 groups. In the tunicamycin + cisplatin group, the MCF-7 cells were pre-treated with tunicamycin (8 µM) for 8 h before incubated with cisplatin (16 µM) for an extra 24 h.
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay for MCF-7 cell viability
The 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) cell proliferation and cytotoxicity assay kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) was adopted to detect the proliferation of MCF-7, ZR-75-30, T47D, MDA-MB-435s, MDA-MB-453, and MDA-MB-231 cells after 24 h culture in 100 µL culture solution, containing 5 µM tunicamycin (Sigma-Aldrich Chemical Company). The MCF-7 cell proliferation in different groups was also testified. The MCF-7 cells at the logarithmic growth phase were digested with trypsin and seeded into a 96-well plate at a density of 5 × 103 cells/well. Meanwhile, the blank control and solvent control groups were set up. Then, the samples were incubated in a CO2 incubator (5% CO2) at 37°C for 24 h before the addition of corresponding drugs. Three duplicate wells were set up in each group. After 24 h, MTT solution (10 µL) was dispensed into each well. Incubated in a CO2 incubator (5% CO2) at 37°C for 3 h, the supernatants were aspirated and dimethyl sulfoxide (DMSO; 100 µL) was added into each well for dissolution. Then, samples were shaken for 10 min, after which the absorbance of each well was detected at 570 nm on a microplate reader and the cell viability was calculated.
Monodansylcadaverine staining for MCF-7 cell autophagy
A volume of 10 mg monodansylcadaverine (MDC) powders (Nanjing Jiancheng Bioengineering Institute) was dissolved in 60 µL of DMSO for preparation of 50 mmol/L concentrated solution (diluted with DMEM to 50 µmol/L working solution before using). The MCF-7 cells at the logarithmic growth phase were digested, resuspended, and seeded into a 6-well plate at a density of 1 × 106 cells/well. When the cells grew to approximately 70% confluence, the cells were treated with corresponding drugs for 24 h, after which DMEM was aspirated and the 6-well plate was washed with phosphate-buffered saline (PBS) twice, refreshed with DMEM containing MDC (final concentration of 50 µM), and incubated at 37°C for 30 min in the dark. Autophagy was analyzed by fluorescence-activated cell sorting (FACS; Becton, Dickinson and Company, Franklin Lakes, NJ, USA).
Flow cytometry for MCF-7 cell apoptosis
Annexin V–fluorescein isothiocyanate (FITC) and propidium iodide (PI) were used for identification of early apoptotic cells and late apoptotic cells. The MCF-7 cells at the logarithmic growth phase were seeded into a 6-welll plate. Then, suspended cells were collected and cells adhering to the wall were digested. The collected cells were stained by the Annexin V-FITC/PI kit (Becton, Dickinson and Company) for 15 min in the dark. After incubation, the samples were immediately analyzed by FACS for avoidance of fluorescence quenching.
Western blotting
The supernatant culture media were aspirated and the cells were washed by D-Hank’s solution for three times before the addition of 500 µL D-Hank’s solution in each well. The cells were scraped and transferred into Eppendorf (EP) tubes which were then centrifuged at 3000 r/min for 5 min with the supernatants aspirated and the cells precipitating in the bottom of EP tubes. Then, the cells were treated with radioimmunoprecipitation assay (RIPA) lysis buffer in a proportion of 1:5 (v/v), placed on a shaker and a piece of ice for 15 s and 30 min, respectively, and sonicated for 10 s until the cell pellets disappeared. The cells were centrifuged at 12,000 r/min (4°C) for 15 min and the supernatants (containing total protein) were collected. A protein standard curve for protein concentration was drawn, which was detected and calculated using bicinchoninic acid (BCA) kit. Extracted proteins (40 µg) were mixed with the loading buffer (5×) and then heated at 100°C boiling water for 5 min. After protein denaturation, the mixture was loaded onto a 10% polyacrylamide gel and underwent electrophoresis under the voltage from 70 V (30 min) to 115 V (1 h), followed by semi-dry transfer to a polyvinylidene fluoride (PVDF) membrane (15 V for 30 min) that was sealed in 5% skim milk for 2 h and incubated with the primary antibodies of relevant proteins in ERS pathway (PEAK, eIF2α, and CHOP), autophagy (Beclin1, LC3II/I, p62, and DAPK1), apoptosis (Bcl-2, Bax, caspase3, caspase8, and caspase9), and the PI3K/AKT/mTOR signaling pathway.
Statistical analysis
SPSS 20.0 software (SPSS Inc., Chicago, IL, USA) was used for statistical analysis, and all data were expressed as mean ± standard error of measurement (SEM). The multiple groups were compared using one-way analysis of variance (ANOVA), and the two groups were compared using the least significant difference (LSD) test. The p < 0.05 showed statistical difference.
Results
ERS inhibits the proliferation of BC cells
The survival rates of different cell lines treated with 5 µM tunicamycin for 24 h were shown in Figure 1(a). The survival rate of MCF-7 cells was significantly higher than that of the ZR-75-30, T47D, MDA-MB-435s, MDA-MB-453, and MDA-MB-231 5 cells (all p < 0.05). The MCF-7 cells were selected for further experiment. The cytotoxicity of MCF-7 cells treated with different concentrations of tunicamycin for 12, 24, and 48 h was detected by MTT assay. According to Figure 1(b), it was shown that cell proliferation was inhibited at different degrees with the increased concentrations of tunicamycin and prolonged incubation time. More specifically, the MCF-7 cells treated with 1 µM tunicamycin for 48 h showed a cell viability of 72.9% ± 1.7%, significantly inhibited cell viability when compared with the control group (p < 0.05). As for the MCF-7 cells treated with tunicamycin at the maximal concentration (8 µM) for 12, 24, and 48 h, the corresponding cell viabilities were 64.5% ± 1.1%, 50.0% ± 1.1%, and 38.5% ± 0.7%, which implied that tunicamycin inhibited the proliferation of MCF-7 cells at both time- and dose-dependent manners (p < 0.05).

Effects of tunicamycin on cell viability of MCF-7, ZR-75-30, T47D, MDA-MB-435s, MDA-MB-453, and MDA-MB-231 cells: (a) cell viability of different cell lines treated with 5 µM tunicamycin for 24 h (*p < 0.05, compared with the other groups) and (b) MCF-7 cell viability treated with different concentrations of tunicamycin (*p < 0.05, compared with the control group; #p < 0.05, compared with the cells treated with 1 µM tunicamycin; ^p < 0.05, compared with the cells treated with 2 µM tunicamycin; &p < 0.05, compared with the cells treated with 4 µM tunicamycin).
ERS inhibits the PI3K/AKT/mTOR signaling pathway in MCF-7 cells
In Figure 2(a), the results of western blotting showed that the protein expressions of PEAK, eIF2, and CHOP in the ERS pathway were upregulated in a dose-dependent manner as the concentrations of tunicamycin increased (p < 0.05), leading to activation of ERS. But the protein expressions of p-PI3K, p-AKT, and p-MTOR in the PI3K/AKT/mTOR signaling pathway were dose-dependently downregulated with increased tunicamycin concentrations (p< 0.05). Thus, expression of the PI3K/AKT/mTOR signaling pathway was inhibited (Figure 2(b)).

Effects of different concentrations of tunicamycin on the relative protein expressions of ERS pathway and the PI3K/AKT/mTOR signaling pathway in MCF-7 cells. (a) Western blotting for the determination of ERS pathway-related protein expression. (b) Western blotting for the determination of PI3K/AKT/mTOR signaling pathway–related protein expression (ERS: endoplasmic reticulum stress; *p < 0.05, compared with the control group; #p < 0.05, compared with the cells treated with 1 µM tunicamycin; ^p < 0.05, compared with the cells treated with 2 µM tunicamycin; &p < 0.05, compared with the cells treated with 4 µM tunicamycin).
ERS inhibits MCF-7 cell proliferation and promotes MCF-7 cell apoptosis by inhibiting the PI3K/AKT/mTOR signaling pathway
As can be seen from the results of MTT assay (Figure 3(a)), the cell viabilities in 12, 24, and 48 h after corresponding treatment were substantially lower in the tunicamycin + BEZ235 group than those in the control group, the tunicamycin group, and the BEZ235 group (p < 0.05), showing a time-dependent manner.
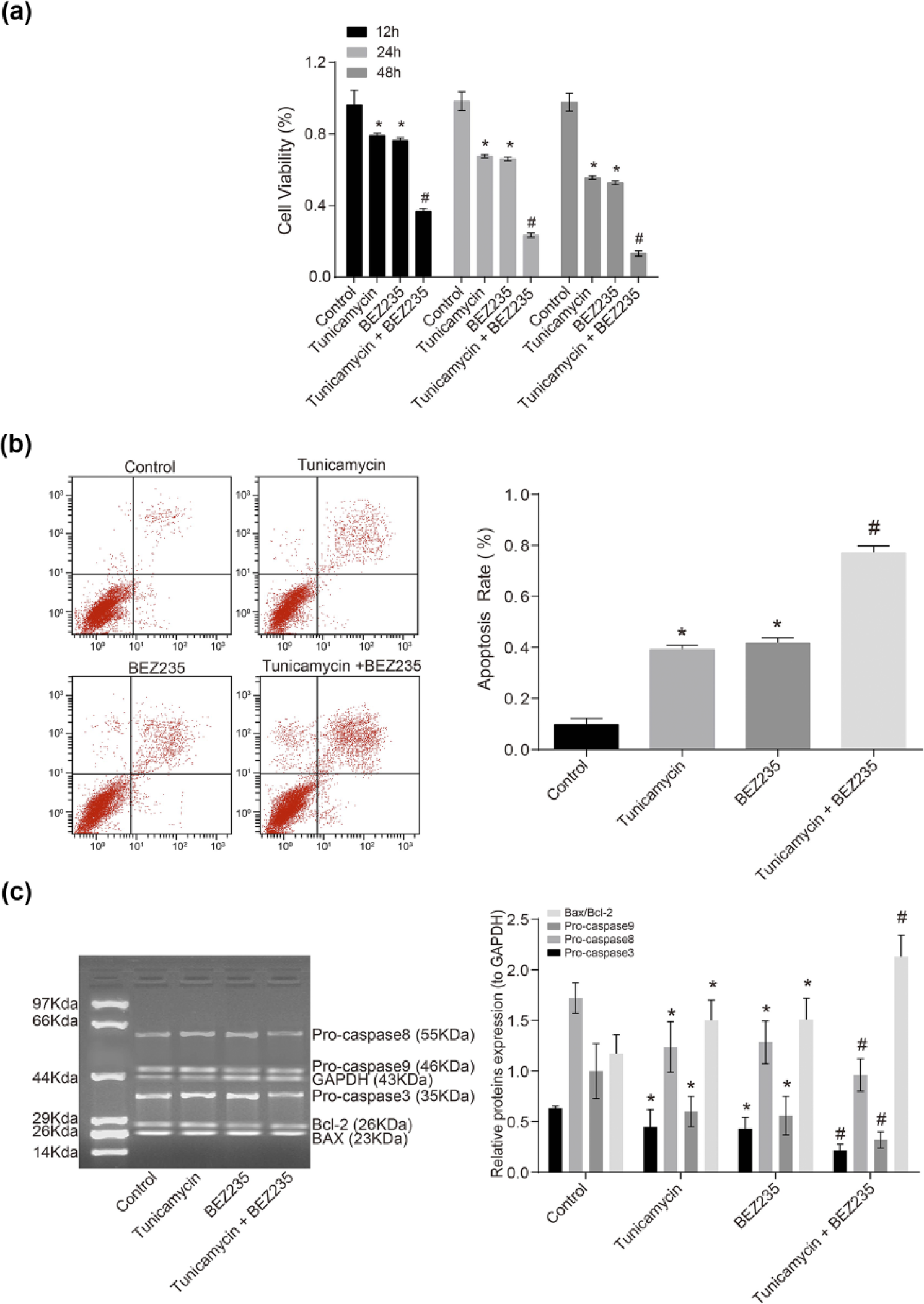
Effects of tunicamycin and BEZ235 on MCF-7 cell apoptosis and expressions of apoptosis-related proteins in MCF-7 cells: (a) MTT assay for the determination of cell viability after treatment, (b) flow cytometry for the determination of cell apoptosis rate after treatment, and (c) western blotting for the determination of relative protein expression after treatment (*p < 0.05, compared with the control group; #p < 0.05, compared with other groups).
From Figure 3(b) presenting the apoptosis rates of MCF-7 cells determined by flow cytometry, it can be seen that the tunicamycin and BEZ235 groups had significantly higher apoptosis rates than the control group (p < 0.05). The tunicamycin + BEZ235 group showed a substantially higher apoptosis rate when compared with the control, tunicamycin, and BEZ235 groups (p < 0.05). The results of western blotting (Figure 3(c)) showed that the protein expressions of pro-caspase3, pro-caspase8, and pro-caspase9 decreased markedly and the ratio of BAX/Bcl-2 increased significantly in the tunicamycin and BEZ235 groups than that of the control group (p < 0.05). Compared with the tunicamycin and BEZ235 groups, significantly downexpressed pro-caspase3, pro-caspase8, and pro-caspase9 proteins and increased ratio of BAX/Bcl-2 were observed in the tunicamycin + BEZ235 group (p < 0.05), indicating the synergistic effects of BEZ235 and tunicamycin on inhibiting the PI3K/AKT/mTOR signaling pathway.
ERS promotes MCF-7 cell autophagy by inhibiting the PI3K/AKT/mTOR signaling pathway
The MCF-7 cells were treated with BEZ235 and/or tunicamycin for 24 h, after which MDC staining was performed, and then, MCF-7 cell autophagy was detected using flow cytometry. It was shown in Figure 4(a) that autophagy occurred more frequently in the tunicamycin, BEZ235, and tunicamycin + BEZ235 groups than that in the control group (p < 0.05). And that autophagy was more common in the tunicamycin + BEZ235 group than the tunicamycin and BEZ235 groups (p < 0.05). According to the results of western blotting (Figure 4(b)), compared with the control group, LC3II/I, Beclin1, and DAPK1 were significantly upregulated in the tunicamycin, BEZ235, and tunicamycin + BEZ235 groups (all p < 0.05), but the P62 was downregulated (p < 0.05). The tunicamycin + BEZ235 group showed significantly increased LC3II/I, Beclin1, and DAPK1 and remarkably decreased P62 in comparison to that of the tunicamycin and BEZ235 groups (all p < 0.05).

Effects of tunicamycin and BEZ235 on MCF-7 cell autophagy and the expressions of autophagy-related proteins in MCF-7 cells: (a) flow cytometry for the determination of autophagy rate after treatment and (b) western blotting for the determination of relative protein expressions after treatment (*p < 0.05, compared with the control group; #p < 0.05, compared with other groups).
ERS increases chemosensitivity of MCF-7 cells by inhibiting the PI3K/AKT/mTOR signaling pathway
Based on the results of MTT assay (Figure 5(a)), it was indicated that the growth inhibition rates were significantly higher in the treatment groups than that of the control group (all p < 0.05). Compared with the cisplatin, tunicamycin + cisplatin and tunicamycin + cisplatin + BEZ235 groups exhibited a relatively higher growth inhibition rate (p < 0.05). The growth inhibition rate substantially increased in the tunicamycin + cisplatin + BEZ235 group than that in the tunicamycin + cisplatin and cisplatin groups (both p < 0.05). According to the results of flow cytometry (Figure 5(b)), it was shown that the apoptosis rates of the control, tunicamycin, cisplatin, cisplatin + BEZ235, tunicamycin + cisplatin, and tunicamycin + cisplatin + BEZ235 groups were 4.5% ± 0.5%, 19.1% ± 2.7%, 30.6% ± 3.3%, 52.0% ± 5.6%, 53.2% ± 6.0%, and 83.9% ± 10.5%, respectively. Compared with the control group, the treatment groups showed significant increases in apoptosis rates (all p < 0.05). The tunicamycin + cisplatin and tunicamycin + cisplatin + BEZ235 groups showed significantly increased apoptosis rate than the cisplatin group (p < 0.05). In the tunicamycin + cisplatin + BEZ235 group, the apoptosis rate was much higher than those in the tunicamycin + cisplatin group (p < 0.05). Thus, it was speculated that both tunicamycin and BEZ235 (a PI3K/mTOR inhibitor) could increase chemosensitivity of MCF-7 cells to cisplatin.

Comparisons of MCF-7 cell proliferation and apoptosis among groups treated with different chemotherapies: (a) MTT assay for determination of growth inhibition rates after treatment and (b) Annexin V-FITC/PI staining for determination of apoptosis rates after treatment (*p < 0.05, compared with the control group; #p < 0.05, compared with the tunicamycin group or the cisplatin group; and &p < 0.05, compared with other groups).
Discussion
BC is the most frequently diagnosed life-threatening cancer in women with increasing trends in its morbidity and mortality. 1 Although surgery, systemic therapy, and radiotherapy have been applied for the treatment of BC, these approaches are limited to some extent, due to potential complications and chemotherapy resistance.5,14 In order to explain the molecular mechanisms underlying autophagy, apoptosis, and chemotherapy resistance, this study examined the ability of ERS induction to circumvent chemotherapy resistance of MCF-7 cells and delineated the intracellular signaling pathway involved in MCF-7 cell autophagy and apoptosis so as to provide evidence for the development of BC-targeted therapies.
According to the study, it has been found that the expressions of PEAK, eIF2, and CHOP in the ERS pathway were upregulated with increased tunicamycin concentrations, in dose- and time-dependent manners. Tunicamycin is an ERS inducer which activates CHOP and induces UPR including phosphorylation of PERK and eIF2α, consequently leading to autophagy and apoptosis due to excessive synthesis or accumulation of unfolded proteins within the ER lumen.9,15,16 According to the research of Zheng et al., 9 CHOP has pronounced cytotoxicity and it plays a key role in ERS-induced cell death. It was reported that tunicamycin treatment significantly triggered autophagy and apoptosis probably by elevating the levels of relevant proteins in the ERS pathway,17,18 which was consistent with the results of this study.
However, tunicamycin exerted similar inhibitory effects as BEZ235 on the expressions of p-PI3K, p-AKT, and p-MTOR in the PI3K/AKT/mTOR signaling pathway and both compounds significantly decreased the levels of pro-caspase3, pro-caspase8, and pro-caspase9 but markedly increased the ratio of BAX/Bcl-2. Besides, the combination of tunicamycin and BEZ235 exhibited significant increases in the expressions of LC3II/I, Beclin1, and DAPK1 but a marked decrease in the level of P62. It is well recognized that the PI3K/AKT/mTOR signaling pathway plays a central role in regulating cell proliferation and apoptosis, and overactivation of the PI3K/AKT/mTOR signaling pathway has been indicated in carcinogenesis of BC. 12 Another study showed that p-PI3K, p-AKT, and p-MTOR are highly expressed in BC, and thus, they could serve as potential molecular therapeutic targets in the treatment of BC. 19 Targeting at the PI3K/AKT/mTOR signaling pathway, a dual PI3K/mTOR inhibitor, BEZ235, was developed to inhibit tumor growth, 20 the mechanism of which might be partially attributed to phosphorylation of relevant proteins in the PI3K/AKT/mTOR signaling pathway. 19 Yang et al. 21 have reported that PI3K/AKT signaling pathway, one of the survival signaling pathways, plays an important role in resisting the apoptosis-inducing ability of ERS. Moreover, it has been proved that BEZ235 inhibited MCF-7 cell proliferation via the PI3K/AKT/mTOR signaling pathway. 22 In addition, increased levels of Bax and decreased levels of Bcl-2 have been indicated in glioma cell lines treated with BEZ235. 23 Based on the fact that tunicamycin exhibited similar effects as BEZ235 on inhibition of the PI3K/AKT/mTOR signaling pathway and stimulation of apoptosis, it was supposed that both chemicals might share similar mechanisms. With respect to autophagy, a recent study showed that tunicamycin enhanced conversion from LC3I to LC3II (autophagosome-associated form) and increased the Beclin1 levels, 24 which had also been described in this study.
Further studies had illustrated that ERS increased autophagy, apoptosis, and chemotherapy resistance of MCF-7 cells via inhibition of the PI3K/AKT/mTOR signaling pathway. Besides, both tunicamycin and BEZ235 (a PI3K/mTOR inhibitor) were shown to increase chemosensitivity of MCF-7 cells to cisplatin. ERS is a highly dynamic organelle which synthesizes and folds a series of structural and functional proteins, and disruption of ER homeostasis disturbs protein folding, leading to accumulation of unfolded and misfolded proteins in the ER lumen as well as apoptosis. It was reported that cisplatin could induce ERS-associated apoptosis in ovarian cancer cells and that ERS tolerance might be involved in cisplatin resistance. 25 In another recent study, negative regulation of the PI3K/AKT/mTOR signaling pathway was shown to mediate apoptosis in BC cells. 12 Therefore, the conclusions from previous studies provide extra evidence further supporting our results. In order to overcome chemotherapy resistance, BEZ235 had been developed and showed effective inhibition on cell growth of various cancers in the presence of cisplatin resistance.26,27
In conclusion, this study demonstrated that tunicamycin could stimulate ERS and downregulate relevant proteins of the PI3K/AKT/mTOR signaling pathway in time- and dose-dependent manners and that ERS increased autophagy and apoptosis of MCF-7 cells as well as chemosensitivity of MCF-7 cells to cisplatin through inhibition of the PI3K/AKT/mTOR signaling pathway. Nevertheless, the relationship between ERS and MCF-7 cells has not been clearly explained. Further studies would be required for validation of our results.
Footnotes
Acknowledgements
The authors would like to acknowledge the helpful comments on this paper received from the reviewers.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The research was supported by Taihang Young Scholar Foundation of Xinxiang Medical University, Doctoral Scientific Research Foundation of Xinxiang Medical University (no. 505079), the National Natural Science Foundation of China (no. U1404816), Scientific Research Foundation of the Higher Education Institutions of Henan Province, China (no. 17A310023), and the National Natural Science Foundation of China (no. 81602132).