
Abstract
Lung cancer, of which non-small cell lung cancer accounts for 80%, remains a leading cause of cancer-related mortality and morbidity worldwide. Our study revealed that the expression of WD repeat containing antisense to P53 (WRAP53) is higher in lung-adenocarcinoma specimens than in specimens from adjacent non-tumor tissues. The prevalence of WRAP53 overexpression was significantly higher in patients with tumor larger than 3.0 cm than in patients with tumor smaller than 3.0 cm. The depletion of WRAP53 inhibits the proliferation of lung-adenocarcinoma A549 and SPC-A-1 cells via G1/S cell-cycle arrest. Several proteins interacting with WRAP53 were identified through co-immunoprecipitation and liquid chromatography/mass spectrometry. These key proteins indicated previously undiscovered functions of WRAP53. These observations strongly suggested that WRAP53 should be considered a promising target in the prevention or treatment of lung adenocarcinoma.
Keywords
Introduction
Lung cancer remains a leading cause of cancer-related mortality and morbidity worldwide, with an estimated 1.7 million new cases and more than 1.4 million deaths per year. 1 Non-small cell lung cancer (NSCLC), such as adenocarcinoma, squamous cell carcinoma, adenosquamous cell carcinoma, and large-cell carcinoma, accounts for 80% of all cases of lung cancer, and the combined 5-year survival rate for all stages and subtypes of NSCLC is low (15%). It has also been estimated that more than 50% of lung-adenocarcinoma patients show local invasion or distant metastasis at the time of diagnosis and that the 5-year survival rate of patients with stage-IV lung cancer is less than 1%. 2 Despite advances in various treatments, such as surgical resection, radiation therapy, chemotherapy, target therapy, and combinations of the above, tumor invasion and metastasis continue to considerably limit treatment options, and the prognosis for patients with early lung adenocarcinoma remains poor. The molecular mechanisms underlying the malignant behavior of lung adenocarcinoma are still unclear. Therefore, it is urgently necessary to characterize and identify novel prognostic markers and therapeutic targets.
The WD repeat containing antisense to P53 (WRAP53, also known as TCAB1 or WDR79) is located on chromosome 17p13 and partly overlaps the P53 tumor suppressor gene in the opposite direction. 3 The WRAP53 gene is alternatively spliced and has three alternative start exons: 1α, 1β, and 1γ, giving WRAP53α, WRAP53β, and WRAP53γ, respectively. Only WRAP53α directly overlaps with the first exon of P53 in an antisense fashion and encodes WRAP53α transcripts, which regulate p53 protein levels by targeting the 5′-untranslated region of P53 messenger ribonucleic acid (mRNA). Despite this connection with p53, neither the transcripts nor the proteins of WRAP53 are involved in the regulation of the P53 gene.3,4
The involvement of WRAP53 in telomere maintenance was discovered only recently in 2009. The WRAP53 protein seemed to be a new subunit of the telomerase enzyme, which is essential to the recruitment of telomerase to Cajal bodies and telomere elongation in human cancer cells. In the nucleus, the WRAP53 protein also binds certain RNA species, known as small Cajal body–specific RNAs (such as telomerase RNA, which is part of the telomerase holoenzyme), and recruits them to Cajal bodies. 5 The WRAP53 protein is an essential component of Cajal-body maintenance; in its absence, Cajal bodies collapse. The WRAP53 protein also interacts with the survival motor neuron (SMN) protein, which is a key regulator of splicing, and recruits the SMN complex from the cytoplasm to Cajal bodies in the nucleus. 6
WRAP53 has been implicated in some human diseases. Mutations in WRAP53 can result in dyskeratosis congenital. 7 The overexpression of WRAP53 and single-nucleotide polymorphisms of WRAP53 have been reported to be related to tumorigenesis in primary nasopharyngeal carcinoma, esophageal squamous cell carcinoma, estrogen-receptor (ER)-negative breast cancer, and ovarian cancer.8–11 The latest research shows that the overexpression of WRAP53 in NSCLC may result in changes to mitochondria and is linked with tumor progression. 12
Our initial research indicated that a knockdown of WRAP53 inhibits the proliferation of WRAP53-expressed lung-adenocarcinoma A549 cells by inducing G1-/S-phase cell-cycle arrest without cell apoptosis as WRAP53 depletion inhibits telomerase trafficking to telomeres in A549 cells, leading to G1 cell-cycle arrest. In this study, we attempted to determine the role of WRAP53 in the biogenesis of lung adenocarcinoma. We obtained the noval findings that WRAP53 is overexpressed in lung-adenocarcinoma surgical specimens relative to specimens from adjacent non-tumor tissues and that the overexpression of WRAP53 is related to tumor size. Intriguingly, the results of co-immunoprecipitation (Co-IP) and mass spectrum (MS) also suggested that WRAP53 triggers the cell-cycle arrest and promotes tumorigenesis by directly interacting with key proteins in earlier researches to contribute to the progressions of miscellaneous tumors.
Materials and methods
Patients and tissue samples
A total of 75 liquid nitrogen–stored human lung-adenocarcinoma specimens, individually matched with those of their adjacent non-tumor tissues (each within 1–3 cm of the corresponding tumor tissue), were obtained from the Department of Thoracic Surgery, First Affiliated Hospital of Zhejiang University, Hangzhou, China between 2013 and 2014. Sufficient high-quality tumor samples were taken by well-trained physicians. Each sample was placed in liquid nitrogen immediately after resection and stored in a refrigerator at −80°C. All specimens were collected after the patients had given their informed consent to participate, and all of the experiments were approved by the institution’s Internal Review and Ethics Board. All procedures involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards.
Reagents and antibodies
The cell culture medium F-12k and RPMI 1640, antibiotics, trypsin, fetal bovine serum (FBS) and phosphate-buffered saline (PBS) were purchased from Gibco Life Technologies (Carlsbad, CA, USA). Other generic chemicals were purchased from Sigma-Aldrich (St. Louis, MO, USA), Thermo Fisher Scientific (Rockford, IL, USA), Merck (Darmstadt, Germany), Amresco (Solon, OH, USA), or the Beyotime Institute of Biotechnology (Shanghai, China).
Quantitative real-time polymerase chain reaction analysis
Total RNA was extracted from the tissues using TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA, USA) and used to synthesize complementary DNA (Bio-Rad, Hercules, CA, USA) according to the standard instructions. Real-time polymerase chain reaction (RT-PCR) was performed according to the manufacturer’s instructions. The primer sequences are shown in Supplemental Data 1. All reactions were performed in triplicate.
Cell culture
Lung-adenocarcinoma A549 and SPC-A-1 cells obtained from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China) were cultured at 37°C in a humidified atmosphere (Thermo Fisher Scientific) in the presence of 5% CO2 and 95% air. The cells were grown in appropriate media (A549 in F-12k and SPC-A-1 in RPMI 1640) containing antibiotics (penicillin at 100 mU/mL and streptomycin at 100 µg/mL) and supplemented with 10% FBS.
Transfection of small interfering RNA (siRNA)
The cells were plated in six-well plates and allowed to attach overnight. Lipofectamine RNAiMAX Transfection Reagent (Invitrogen Life Technologies) was used to perform a transient transfection of control siRNA and WRAP53 siRNA molecules for 48 h, following the instructions of the manufacturer. The sequences of the control siRNA and WRAP53 siRNA molecules are shown in Supplemental Data 2.
Colony-formation assay and 3-(4,5-cimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide cytotoxicity assay
To perform the colony-formation assay, cells were transfected as needed in six-well plates. After 48 h, the cells were harvested by trypsinization and then seeded at 2000 cells per well in triplicate. The cells were then incubated at 37°C in 5% CO2 for 14 days without disturbance. The colonies formed were fixed in 4% paraformaldehyde and stained with 0.1% crystal violet for 30 min, washed with distilled water, dried overnight, and counted the next day. For the 3-(4,5-cimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay, the cells were transfected with siRNA and cultured in 96-well flat-bottomed microtiter plates supplemented with F-12k containing 10% heat-inactivated FBS in a humidified atmosphere (95% air and 5% CO2) at 37°C. After 24, 48, and 72 h of incubation, the in vitro cytotoxic effects of these treatments were determined by an MTT assay (at 490 nm). All reactions were performed in triplicate.
Cell-cycle assay
To perform the cell-cycle assay, the cells were first collected and washed with PBS. Cell pellets were obtained by centrifugation and the supernatant was discarded. Next, 1 mL of DNA staining solution (Multi Sciences Biotech Co., Ltd., Hangzhou, China) was added and blended by vortexing for 10 s. In preparation for flow cytometry, the cells were incubated for 30 min at room temperature in the dark. For the apoptosis assay, the cells were resuspended in 0.5 mL of binding buffer containing Annexin V (1:50) and 40 ng/sample of propidium iodide (1:50; all from BD Biosciences, Franklin Lakes, NJ, USA) and incubated for 30 min at 37°C in the dark prior to flow cytometry. The samples were analyzed using an LSR II Flow Cytometer (BD Biosciences) and ModFit LT software (Verity Software House, Topsham, ME, USA). At least 10,000 cells from each sample were analyzed, and the experiments were repeated three times.
Cell lysis and western blotting
The cells were washed with PBS and lysed in ice-cold radioimmunoprecipitation assay lysis buffer (Beyotime Institute of Biotechnology). The lysates were centrifuged at 16,000×g at 4°C for 10 min, and the supernatants were transferred to new tubes. The samples were then boiled in electrophoresis sample buffer and run on sodium dodecyl sulfate polyacrylamide gel electrophoresis gels at varying acrylamide concentrations, depending on the molecular weight of the proteins to be analyzed. After electrophoresis, the proteins in the gel were transferred to polyvinylidene difluoride membranes (Millipore, Bedford, MA, USA). The membranes were blocked in Tris-buffered saline with Tween 20 (TBST; 100 mM Tris, pH 7.5; 150 mM of NaCl; and 0.05% Tween 20) containing 2% of bovine serum albumin for 1–3 h and then incubated overnight with the corresponding antibody: WRAP53 (Cat. No. NBP1-92592; Novus Biologicals, Littleton, CO, USA), CDK4 (Cat. No. 12790; CST, Danvers, MA, USA), p-Rb (S795, Cat. No. 9301; CST), or p-Rb (S807/811, Cat. No. 8519; CST; 1:1000). After being washed three times with TBST for 10 min, the membranes were incubated with horseradish peroxidase–conjugated anti-mouse or anti-rabbit secondary antibodies for 1–2 h (1:1000). After incubation with the secondary antibodies, the membranes were washed three times with TBST and the bands were visualized on a Kodak XBT-1 Film (Kodak, Rochester, NY, USA). All reactions were performed in triplicate.
Cell synchronization
A double-thymidine block was used to synchronize the A549 cells in the early S phase: the cells were treated with 2.5 mM of thymidine (Sigma Aldrich) for 20 h (first block) before switching to 10% FBS medium for 12 h (first release). To release the cells from synchronization, they were washed with PBS and cultured in a pre-warmed, drug-free, fresh medium for 12 h. Next, the normal medium was replaced with 2.5 mM of thymidine for another 20 h (second block). Finally, the cells were released by adding 10% FBS medium every 0.5 h; cells were collected at each interval. Each group of cells was washed twice with PBS. The efficacy of cell synchronization was confirmed by flow cytometry before further analysis.
Co-IP and MS
Cell lysates were obtained from the synchronized A549 cells using a specific IP lysis buffer (Thermo Fisher Scientific). Next, the cell lysates were incubated with control or indicated WRAP53 antibodies (2–5 µg/mg) overnight at 4°C. The immunocomplexes were each pelleted by incubation with 40 µL of protein A/G agarose (Thermo Fisher Scientific) for 3 h at 4°C with slow agitation and centrifugation for 15 s. The pellets were washed three times with the IP lysis buffer mentioned above, containing a protease inhibitor cocktail (Roche, Mannheim, Germany) buffer, and boiled in an electrophoresis sample buffer. Finally, the samples were centrifuged at 16,000×g at 4°C for 1 min, and the supernatants were transferred to new tubes for liquid chromatography/mass spectrometry (LC/MS) analysis.
The LC/MS analysis was performed on an electrospray ionization quadrupole time-of-flight (ESI-Q-q-TOF) mass spectrometer (micrOTOF-Q II; Bruker, Fremont, CA, USA) at the Shanghai Institute for Biological Sciences, Chinese Academy of Sciences, Shanghai, China. The LC/MS raw data were analyzed using the proteomics software package Mascot with the IPI_human database (IPI_ human_v3.87). 13
Statistical analysis
Continuous variables were compared between the human-adenocarcinoma specimens and the adjacent non-tumor tissue specimens using the denary logarithms of albumin/creatinine ratios from low to high, as shown in Figure 1. The χ2 test, a method of univariate analysis, was used to examine the clinical characteristics of WRAP53. The results of at least two independent experiments were analyzed. The statistical data were presented in the form of mean ± standard deviation. Differences were considered to be statistically significant when p values were smaller than 0.05. The data were analyzed using the software packages IBM SPSS 22.0 (IBM Co., Chicago, IL, USA) and the GraphPad Prism 6.01 (GraphPad Software, Inc., La Jolla, CA, USA).

The expression level comparison of WRAP53 in lung-adenocarcinoma specimens. The WRAP53 expression is higher in lung-adenocarcinoma specimens than those of adjacent non-tumor tissues. The denary logarithms of A/C ratios (C means the expression of WRAP53 in cancer tissue and A means the expression of WRAP53 in adjacent non-tumor tissue, all with ΔCT value) in the order of low to high were shown. Negatives showed a higher expression in lung-adenocarcinoma specimens than those of adjacent non-tumor tissues.
Results
WRAP53 expression is higher in lung-adenocarcinoma specimens than in specimens taken from adjacent non-tumor tissues
We first used quantitative RT-PCR (qRT-PCR) analysis to measure the expression of WRAP53 in 75 human specimens. Each bar represented a single patient. The qRT-PCR results showed that the level of RNA in WRAP53 was higher in lung-adenocarcinoma specimens than in specimens from adjacent non-tumor tissues derived from the same patient (Figure 1). This finding was consistent with the immunohistochemical results of the studies mentioned above. 12
Clinical characteristics of WRAP53-overexpressed lung adenocarcinomas
Lung-adenocarcinoma samples were derived from 46 men and 29 women aged between 38 and 81 (mean = 59.59) years. This population comprised 40 non-smokers (53.3%) and 35 former or current smokers (46.7%). Tumor size ranged from 0.8 to 10 cm (mean = 3.26 cm), and 37 samples (49.3%) were classified as pathologic stage I, 16 (21.3%) as stage II, 21 (28.0%) as stage III, and 1 (1.4%) as stage IV. WRAP53 overexpression was found in 49 of the 75 specimens (65.3%). The patient characteristics are summarized in Table 1.
Clinical characteristics and univariate analysis of the prevalence of WRAP53 overexpression for patients with NSCLC.

NSCLC: non-small cell lung cancer.
According to the median age of 60 years, all patients could be divided into two groups.
Smoking history was defined as smoking at least 100 cigarettes prior to the interview date. 14
Two cases of left pneumonectomy were excluded.
One case (No. 84) which was difficult to measure its tumor size was excluded. We described tumor of NSCLC no larger than 3.0 cm as T1 stage. So, we divided all patients into two groups. 15
A p value <0.05 is considered statistically significant.
Analysis revealed that WRAP53 overexpression was prevalent among the 75 patients. The prevalence of WRAP53 overexpression was significantly higher in patients with tumors larger than 3.0 cm than in patients with tumors smaller than 3.0 cm (p < 0.05). WRAP53 overexpression was not found to be significantly correlated with age, gender, smoking history, tumor location, lymph-node metastasis, or tumor-node-metastasis stage.
WRAP53 knockdown reduces proliferation of A549 cells
To characterize the tumor-promoting function of WRAP53, we first knocked down WRAP53 expression by performing a 48-h transient transfection of three WRAP53 siRNAs into lung cancer A549 cells. The most effective siRNA was chosen for use in further experiments (Figure 2(a)). An MTT assay revealed that WRAP53 downregulation induced a significant time-dependent inhibition of cell proliferation in the A549 cells (Figure 2(b)). Similarly, a colony-formation assay revealed that cell proliferation in A549 cells compared with vector-control cells was significantly suppressed by the knockdown of WRAP53 (colony numbers reduced to <50% of controls; Figure 2(c)).

WRAP53 knockdown reduced the proliferation of A549 cells. (a) The effects of WRAP53 siRNA on the expression of WRAP53 were shown. Western blot analyses demonstrated the effect of WRAP53 siRNA transfection on WRAP53 expression in A549 cells. (b) Knockdown of WRAP53-inhibited proliferation of A549 cells were measured by MTT assay. The values obtained from transfected and control cells represent mean ± SD of three independent experiments (*p < 0.05). (c) Representative colony-formation assays of WRAP53 siRNA transfected A549 cells and control cells. All reactions were performed in triplicate.
WRAP53 downregulation causes G1-/S-phase cell-cycle arrest in A549 cells
To clarify the role of WRAP53 in promoting tumor-cell growth, we used flow cytometry to examine the cell cycle and apoptosis of A549 cells transfected by WRAP53 siRNA. Compared with control cells, A549 cells with a lower expression of WRAP53 showed cell-cycle arrest at the G1/S phase, with an accumulation of cells in the G1 and S phases and a decrease in G2-phase cells (Figure 3(a)). We used an Annexin V/propidium iodide apoptosis detection kit to examine the apoptosis of the A549 cells and found that the percentage of apoptotic cells did not differ significantly between the WRAP53 transfectants and the control siRNA transfected cells (Figure 3(b)).
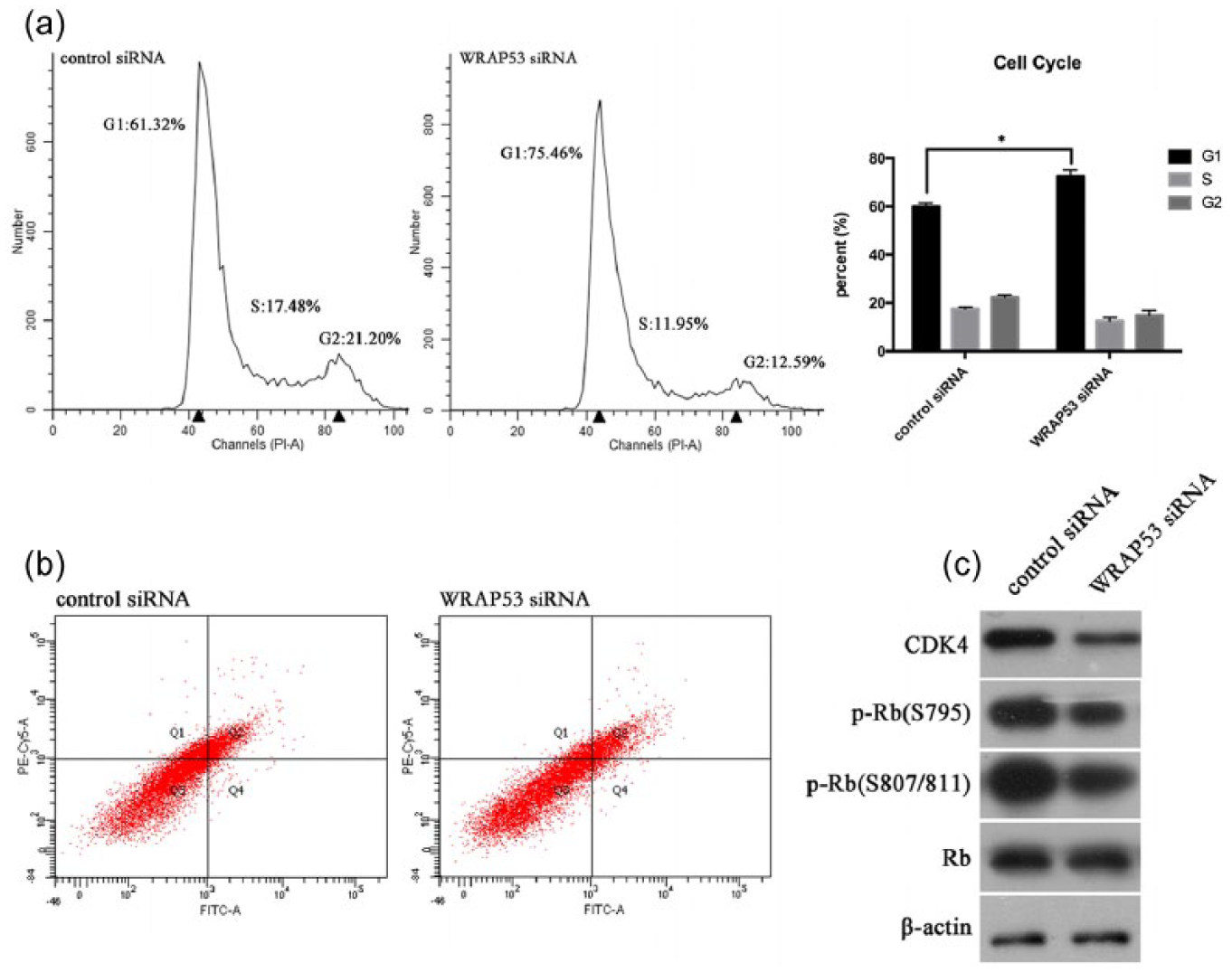
WRAP53 knockdown induced G1-/S-phase cell-cycle arrest but not apoptosis of A549 cells. (a) Cell-cycle profiles of A549 cells transfected with WRAP53 siRNA and control cells were determined by flow cytometry and error bars. All reactions were performed in triplicate, and representative cell-cycle analysis and flow cytometry data were shown. The cell count versus PI staining is shown (n = 10,000 per treatment) (*p < 0.05). (b) Apoptosis assays with A549 cells transfected with WRAP53 siRNA and control cells were shown by flow cytometry analysis. All reactions were performed in triplicate, and representative dot plots of A549 cells transfected with control siRNA or WRAP53 siRNA were shown by flow cytometry. The cell count versus PI staining is shown (n = 10,000 per treatment). (c) Western blot analyses of G1/S cell-cycle regulation and checkpoint proteins with A549 cells transfected with WRAP53 siRNA and control cells. All reactions were performed in triplicate and representative figures were shown.
To confirm that G1-/S-phase cell-cycle arrest had occurred, we verified the levels of G1/S cell-cycle regulation and checkpoint proteins using western blot analysis. Unsurprisingly, the expression of cell-cycle regulation proteins correlated with the G1/S phase, such as CDK4, along with the expression of the G1/S checkpoint proteins p-Rb S795 and S807/811, decreased to varying degrees in the experimental group of A549 cells transfected with WRAP53 siRNA. Overall, this decrease in expression was greater than that in the control group (Figure 3(c)).
Bioinformatic analysis of proteins interacting with WRAP53 indicates a close connection between WRAP53 and the tumorigenesis of lung cancer
WRAP53 has been reported to be an important component of telomerase, responsible for controlling telomerase trafficking and essential to Cajal-body localization and telomere synthesis in the early S phase. 5 The finding that WRAP53 downregulation caused G1-/S-phase cell-cycle arrest in A549 cells led us to hypothesize that WRAP53 has a function in the early S phase other than that previously reported by our research group (namely that WRAP53 depletion inhibits telomerase trafficking to telomeres in the A549 cells, leading to G1 cell-cycle arrest). 16 To test this hypothesis, we accessed early S-phase A549 cells using a double-thymidine block (detected by flow cytometry). The results showed that the early S phase occurs 0–2 h after the G1 phase in A549 cells (Figure 4).

A double-thymidine block accessed the synchronized A549 cells. The flow cytometry data shown that the cell cycle of A549 cells was synchronized in early S phase (the cells were released from synchronization between 0 and 2 h), middle and late S phase (the cells were released from synchronization between 2 and 6 h), and G2 phase (the cells were released from synchronization after 6 h). All reactions were performed in triplicate.
Co-IP experiments were carried out in the A549 cells using an anti-WRAP53 antibody. The Co-IP products were analyzed by LC/MS. In total, 534 proteins involved in cellular progress, single and multicellular organism progress, metabolic progress, biological regulation, and responses to stimulus were identified as WRAP53-interacting partners (Figure 5). These proteins were classified and analyzed according to their biological function, and those listed in Table 2 were selected as promising targets for further analysis.

Bioinformatic analysis of proteins interacting with WRAP53 indicates a close connection between WRAP53 and the tumorigenesis of lung cancer. (a) The cellular component analysis of WRAP53-interacted proteins in the early S-phase A549 cells. (b) The molecular function analysis of WRAP53-interacted proteins in the early S-phase A549 cells. (c) The biological process analysis of WRAP53-interacted proteins in the early S-phase A549 cells.
Candidate proteins interacting with WRAP53 protein were listed below.

Further validation of our research through another lung-adenocarcinoma cell line SPC-A-1
We have demonstrated above that the knockdown of WRAP53 could effectively inhibit the proliferation of A549 cells through the induction of G1-/S-phase cell-cycle arrest. As our research was primarily performed in a single cell line, other lung-adenocarcinoma cell lines should be examined to demonstrate broader observations. So, we took out the experiments with another cell line SPC-A-1. The biological phenomena of SPC-A-1 cells were consistent with the results of A549 cells above as shown in Figure 6. The knockdown of WRAP53 reduced the proliferation of SPC-A-1 cells (Figure 6(a) and (b)). And the WRAP53 downregulation could cause G1-/S-phase cell-cycle arrest in SPC-A-1 cells (Figure 6(c) and (d)). All these findings proved from one side to another that the downregulation of WRAP53 was associated with the tumorigenesis of lung-adenocarcinoma cells.
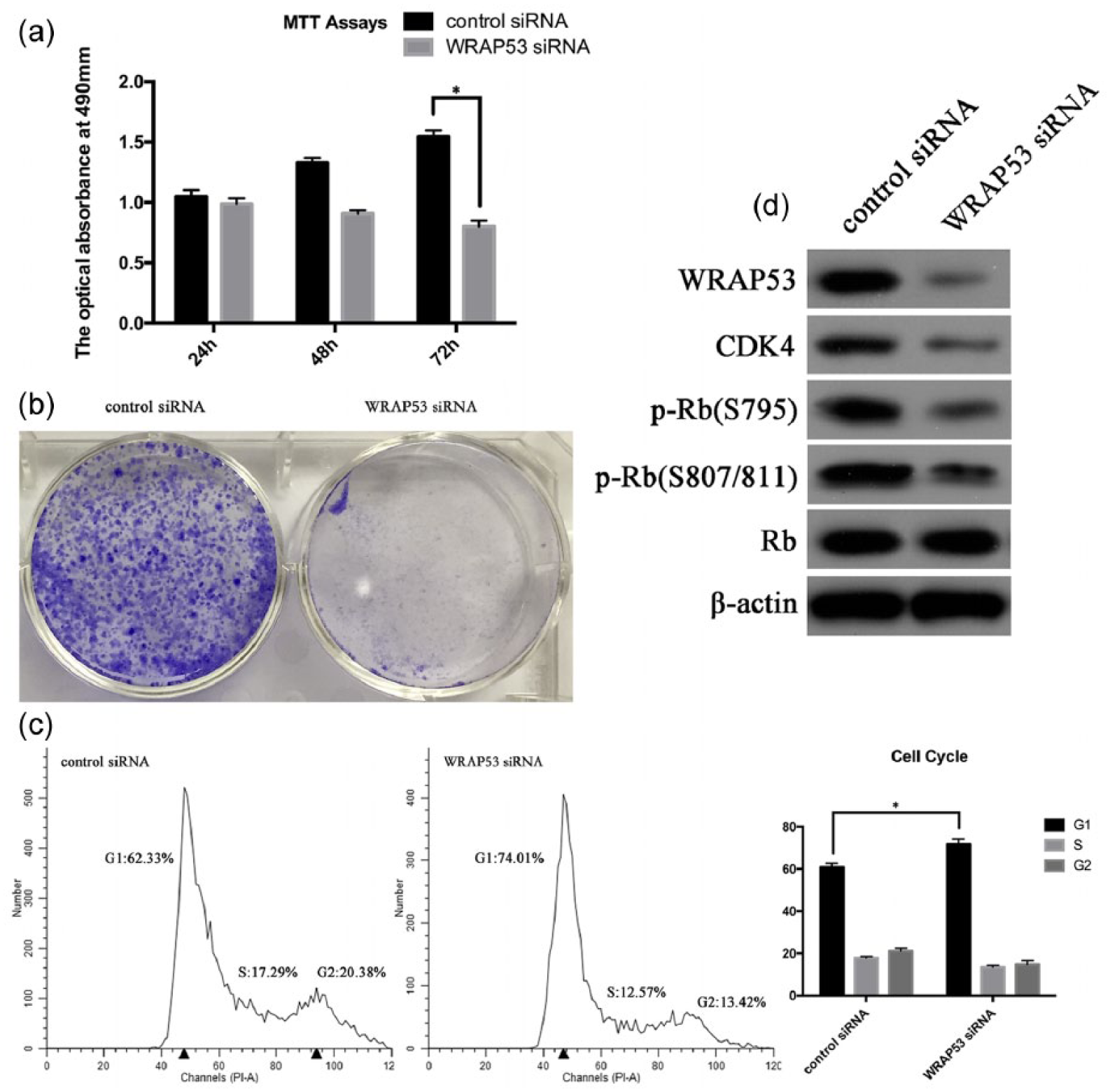
Further validation of our research through another lung-adenocarcinoma cell line SPC-A-1. (a) Knockdown of WRAP53-inhibited proliferation of SPC-A-1 cells were measured by MTT assay. The values obtained from transfected and control cells represent mean ± SD of three independent experiments (*p < 0.05). (b) Representative colony-formation assays of WRAP53 siRNA transfected SPC-A-1 cells and control cells. (c) Cell-cycle profiles of SPC-A-1 cells transfected with WRAP53 siRNA and control cells were determined by flow cytometry and error bars. All reactions were performed in triplicate and representative cell-cycle analysis and flow cytometry data were shown. The cell count versus PI staining is shown (n = 10,000 per treatment) (*p < 0.05). (d) Western blot analyses demonstrated the effect of WRAP53 siRNA transfection on WRAP53 expression in SPC-A-1 cells, as well as the G1/S cell-cycle regulation and checkpoint proteins of SPC-A-1 cells transfected with WRAP53 siRNA and control cells. All reactions were performed in triplicate and representative figures were shown.
Discussion
The role of abnormal molecular activity in triggering uncontrolled cell proliferation is one of the hotspots of cancer research. In 1993, the WRAP53 gene was first reported to cause dyskeratosis congenita, a telomere biological disorder characterized by the triad of dysplastic nails, lacy reticular pigmentation of the upper chest or neck, and oral leukoplakia, and to be responsible for shortening telomeres (telomerase had not yet been discovered in 1993). 17 The participation of WRAP53 in cell activities has since been reported mainly in terms of its roles as an antisense of the P53 gene or a holoenzyme subunit of telomerase.3,5 As little research has been conducted on the additional functions of WRAP53 in lung cancer, especially lung adenocarcinoma, its role in this disease remains unclear. The overexpression of WRAP53 is now considered a biomarker for a variety of cancers, such as colorectal cancer, hepatocellular carcinoma, head and neck carcinomas, breast cancer, esophageal squamous cell carcinoma, and rectal cancer,8,9,18–21 which is consistent with the evidence obtained in this study of high WRAP53 expression in lung-adenocarcinoma tumor tissues and cell lines. The prevalence of WRAP53 overexpression was found to be significantly higher in patients with tumors larger than 3.0 cm, which led us to infer that the expression of WRAP53 is associated with the tumor-growth ability of lung adenocarcinoma. Of course, more clinical samples should be tested to verify this finding in future research.
Our previous results indicated that WRAP53 knockdown inhibits the proliferation of A549 cells. 16 This finding was confirmed experimentally in this study. We performed flow cytometry to analyze the cell-cycle distribution after transfection with WRAP53 siRNA for 48 h to determine whether the inhibition of cell proliferation was associated with cell-cycle arrest or programmed cell death. The results provided strong evidence that WRAP53 downregulation causes G1-/S-phase cell-cycle arrest but not apoptosis in A549 cells. This led us to believe that the WRAP53 protein has a function in cell-cycle regulation in addition to its role as an antisense of the P53 gene or a holoenzyme subunit of telomerase. First, therefore, we accessed early S-phase A549 cells using the double-thymidine block method. Next, we performed Co-IP with these synchronized A549 cells to pull down proteins that might interact with the WRAP53 protein in the early S phase and conducted LC/MS analysis to identify proteins associated with WRAP53 that affected tumorigenesis through the regulation of cell-cycle progression. The LC/MS raw data were analyzed using the IPI_human database. The potential WRAP53-interacting proteins were analyzed as described below.
DKC1 and GAR1 proteins are box H/ACA ribonucleoproteins (RNPs), protein–RNA complexes responsible for pseudouridylation, the most common type of post-transcriptional modification of cellular RNA. H/ACA RNPs play essential roles in ribosome biogenesis, the post-transcriptional processing of nascent ribosomal RNA and spliceosomal RNA, telomerase assembly and stabilization, and subsequently cellular processes such as protein synthesis, gene expression, and chromosome stabilization. 22 Mutations in the protein components of H/ACA RNPs are associated with the human bone marrow failure syndrome dyskeratosis congenita.23,24 The depletion of DKC1 can induce cell-cycle arrest in tumor cells, 25 as can the depletion of GAR1. 26 However, irrespective of the physiological responses of these proteins to genotoxic stress, H/ACA RNPs may participate individually in the regulation of DNA damage repair (DDR) biomarkers through mechanisms that are dependent on the specific type of damage incurred. For example, exposure to doxorubicin and ultraviolet-C alters the cellular localization of GAR1, which moves from the nucleolus to the nucleoplasm, but not that of DKC1 protein. 26 The relationship between WRAP53, as a subunit of telomerase, and these two brother proteins may offer a new explanation for these divergent findings.
RUVBL1 is a highly conserved eukaryotic ATPase that belongs to the superfamily of ATPases associated with diverse cellular activities. In 1998, RUVBL1 was first found to be a growth-promoting gene in eukaryotic cells. 27 Evidence has since been provided in numerous publications that RUVBL1 may have additional functions that are highly relevant to cancer. RUVBL1, like RUVBL2, is an essential component of multiple protein complexes such as the large RNA polymerase II holoenzyme, 27 the TATA-binding protein, 28 the TIP60 histone acetyltransferase complex, 29 the INO80 and SWR1 chromatin-remodeling complexes, 30 an R2TP complex involved in the biogenesis of small nucleolar RNPs, 31 and complexes that regulate the activity of phosphatidylinositol 3-kinase (PI3K)-like kinases, 32 although the functions and relationships of RUVBL1 in such a diverse set of complexes are still unclear. RUVBL1 has been widely reported to be a component of telomerase and to play important roles in telomerase assembly. 33 Levels of RUVBL1 can also regulate H/ACA RNP biogenesis 34 and trigger DNA damage, 35 which is associated with the main functions of the DKC1 and GAR1 proteins mentioned above. Furthermore, several studies have indicated that the RUVBL1 protein plays a chaperone-like role in the assembly of complexes. 36 RUVBL1 has also been reported to be involved in the activities of some oncogenes and transcription factors, such as β-catenin, 37 c-Myc, 38 COS-2, 39 YY1, 40 p53, 41 and NF-Κb. 31 Considering the function of WRAP53, further research is necessary to explore the role of RUVBL1 in lung adenocarcinoma and the interaction of these two proteins, and our study helps to meet this need.
The EEF1A2 gene encodes an isoform of the alpha subunit of the elongation factor-1 complex, which is responsible for the enzymatic delivery of aminoacyl transfer RNAs (tRNAs) to the ribosome. 42 EEF1A2 has been shown to be an important oncogene in almost all kinds of tumor, such as ovarian cancer, 43 breast cancer, 44 lung adenocarcinoma, 45 hepatocellular carcinoma, 46 pancreatic carcinoma, 47 prostate cancer, 48 and gastric cancer. 49 The overexpression of EEF1A2 induces the formation of filopodia in breast cancer cells, which is dependent on the activity of PI3K and Akt kinases. 50 A recent study of hepatocellular carcinoma demonstrated that the EEF1A2/PI3K/AKT/mTOR axis promotes the protumorigenic stabilization of the MDM4 proto-oncogene via a post-transcriptional mechanism. The activation of the EEF1A2/PI3K/AKT/mTOR/MDM4 axis significantly influences the survival probability of hepatocellular-carcinoma patients in vivo. 51 The downregulation of EEF1A2 by direct interaction with the tumor suppressor protein p16 can effectively inhibit the growth of ovarian cancer cells. 52 The focus of most recent research on the cellular progression of EEF1A2 in lung cancer has been its clinical prognostic significance; the biological contribution of EEF1A2 to lung cancer is still unclear. Negative immunostaining for EEF1A2 has been shown to be an independent prognostic factor that is significantly correlated with lymph-node metastasis. 53 Considering the role of the transcription factor as well as the cellular structural function of EEF1A2, the question of whether the telomere-maintenance role of WRAP53 depends on the interaction of these two proteins needs further research.
HSPA2, first discovered in renal cancer cells as a mutation in the heat shock protein 70 family, 54 is a protein essential to spermatogenesis. HSPA2 is required for the growth and survival of cancer cells in nasopharyngeal carcinoma, 55 esophageal squamous cell carcinoma, 56 NSCLC, 57 and pancreatic cancer. 58 The expression of HSPA2 in hepatocellular-carcinoma cells is upregulated in response to hypoxia via the hypoxia inducible factor 1 binding site in the HSPA2 promoter. 59 High HSPA2 expression is significantly related to shorter overall survival (OS) in stages I–II lung cancer patients. 57 Elevated HSPA2 RNA transcription may promote cancer-cell migration and invasion through enforced JAG1 transcription. 60 However, compared with studies of other heat shock protein 70 family members, too little research has been conducted on the mechanism by which HSPA2 regulates lung tumorigenesis. The role of HSPA2 is not simply that of a biomarker. More studies have recently been undertaken to explore the contribution of HSPA2 to the cellular functions of lung adenocarcinoma.
The Warburg effect describes the preferential use by tumor cells of glycolysis rather than oxidative phosphorylation for energy production. Tumor cells exclusively express the embryonic M2 isoform of pyruvate kinase (PKM2). PKM2 is a key enzyme in aerobic glycolysis, known to be associated with the metabolic rewiring of cancer cells and considered an important cancer therapeutic target. The role of PKM2 in lung cancer progression has been widely reported, and multiple mechanisms have been identified.61–63 PKM2 is also associated with clinical strategies for treating lung cancer, such as methods of overcoming docetaxel 64 and gefitinib 65 resistance, platinum-based chemotherapy, 66 and radiosensitivity. 67 However, no researchers have reported an association between PKM2 and telomere or telomerase. Intriguingly, a recent study showed that P53 (N340Q/L344R) can accelerate the growth of liver cancer cells Hep3B by activating telomerase and prolonging telomere through the cascade mediated by PKM2. 68 Considering the roles of WRAP53 as an antisense of the P53 gene and a holoenzyme subunit of telomerase, together with the LC/MS finding that PKM2 interacts with WRAP53, we hypothesize that WRAP53 may take part in this cascade. Previous studies have demonstrated only that WRAP53α is the only WRAP53 variant to directly overlap the first exon of P53 in an antisense fashion and encode WRAP53 transcripts; it remains unclear whether WRAP53β/γ and P53 are related. The interaction of WRAP53 and PKM2 offers new insights into this area.
In sum, we clarified the clinical overexpression of WRAP53 and its prognostic value in lung adenocarcinoma, as well as the inhibitory effect of WRAP53 on the growth of A549 and SPC-A-1 cells via G1/S cell-cycle arrest. We also identified several key proteins that interact with the WRAP53 protein through Co-IP, LC/MS and bioinformatic analysis. All of these proteins should be considered promising targets of future research. The function of WRAP53 in lung cancer is not well understood, due to a lack of attention in the literature; however, our findings offer new insights into the functional role of WRAP53 in lung carcinogenesis. We will conduct additional studies to verify and further explore the mechanisms of the interaction between WRAP53 and the above key proteins. We believe that studying these proteins will reveal hitherto undiscovered functions of WRAP53. Overall, these observations strongly suggest that WRAP53 should be considered a promising target in the prevention or treatment of lung adenocarcinoma.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by National Natural Science Foundation of China with the grant identification number 31170720.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.