
Abstract
Numerous studies have proposed the transplantation of mesenchymal stem cells (MSCs) in the treatment of typical type 2 diabetes mellitus (T2DM). We aimed to find a new strategy with MSC therapy at an early stage of T2DM to efficiently prevent the progressive deterioration of organic dysfunction. Using the high-fat-fed hyperinsulinemia rat model, we found that before the onset of typical T2DM, bone marrow-derived MSCs (BM-MSCs) significantly attenuated rising insulin with decline in glucose as well as restored lipometabolic disorder and liver dysfunction. BM-MSCs also favored the histological structure recovery and proliferative capacity of pancreatic islet cells. More importantly, BM-MSC administration successfully reversed the abnormal expression of insulin resistance-related proteins including GLUT4, phosphorylated insulin receptor substrate 1, and protein kinase Akt and proinflammatory cytokines IL-6 and TNFα in liver. These findings suggested that MSCs transplantation during hyperinsulinemia could prevent most potential risks of T2DM for patients.
Keywords
Introduction
Diabetes mellitus (DM) is a major risk factor for many diseases such as ischemic heart disease and stroke, chronic kidney disease, and blindness among adults1–3. Long-term high-fat food is one of the causes leading to insulin resistance (IR) followed by a compensatory hyperinsulinemia4–6. Because of the high secretary activity, β cells are constantly exposed to various kinds of stresses, such as glucolipotoxicity and oxidative stress7,8. Eventually, this results in β-cell death, which is characterized as typical type 2 diabetes mellitus (T2DM) characterized by hyperinsulinemia and hyperglycemia9,10.
IR might last for 10 years before the onset of β-cell dysfunction and diabetes 11 ; therefore, alleviation of IR at the early stage could be the most efficient approach to prevent progressive and inexorable β-cell dysfunction. Clinical treatment of T2DM including oral antidiabetic drugs and exogenous supply of insulin could reverse neither IR nor β-cell dysfunction12,13.
Mesenchymal stem cells (MSCs) can differentiate into different types of connective tissue cells, which have the capability to produce bone, adipose, and cartilage, modulate the local environment, activate endogenous progenitor cells, and secrete various factors14,15. Through clinical trials and mouse models, they have successfully restored insulin and stimulated glucose uptake in typical T2DM16–20. However, most effects are based on regenerating injured pancreatic tissues and anti-inflammatory or paracrine effects16–20.
New therapeutic strategies to ameliorate IR have been tried with modulating intestinal microbiota 21 , autophagy 22 , and cell therapy with MSCs23–25. These reports present remarkable alleviation of IR by MSCs in a typical T2DM rats which has proceeded to a late state of disease. However, whether MSCs could prevent the deterioration of hyperinsulinemia at early stage of T2DM is not clear.
Therefore, we aimed to investigate the effect of bone marrow-derived MSCs (BM-MSCs) on hyperinsulinemia at the early stage of T2DM. The results suggested that early transplantation of MSCs holds a promising role in controlling the progress of T2DM at early stage.
Materials and Methods
Animals and Sample Collection
Male Sprague-Dawley rats (approximately 200 g, HFK bioscience, Beijing, China) were used for all studies. All experimental procedures were approved by Tongji Medical College, Huazhong University of Science and Technology Institutional Animal Care and Use Committee (2016IACUC number, 644). All efforts were made to reduce the number of animals tested and their suffering. Animals were fed either a normal chow diet or a high-fat diet (Animal Center, Huazhong University of Science and Technology)
26
for 4 wk. On experimental days, food was removed at 8
Oral glucose tolerance test (OGTT) was assessed when animals were fasted overnight to determine their glucose response to the oral administration (by gavage) of a solution of 20% glucose (2 g/kg) before (time 0) and 30, 60, and 120 min after administration of glucose 26 .
BM-MSC Preparation and Administration
BM-MSCs were isolated following a previously described method 27 . Briefly, rats were sacrificed and their hind limbs were harvested, bone marrow was flushed out and collected in Dulbecco's Modified Eagle's Medium (DMEM) (Gibco, Grand Island, NY, USA) supplied with 10% fetal bovine serum (Gibco). Thereafter cells were cultured in a 25-cm2 flask in 5% CO2 incubator at 37 °C. Nonadherent cells were removed after 24 h and adherent cells were passaged every week using 0.05% trypsin. About 5 × 105 (for high concentration) or 105 (for low concentration) cells between passages 3 to 6 were intravenously injected via tail vein.
Histological and Immunohistochemical Staining
Pancreases were freshly removed from the rats, 6-µm sections were cut immediately, and fixed with acetone for hematoxylin-eosin (H&E) staining using standard techniques. Some sections were subjected to immunohistochemical staining (animals were previously intravenously injected with BrdU for 3 days). After the process consisting of 10 min of fixation with acetone, 2 h of permeabilization with 0.3% Triton X-100 (Sigma, St. Louis, MO, USA), and 1 h of blocking with 3% albumin from bovine serum (BSA) (Gibco) at room temperature, the sections were incubated with the primary antibody rabbit anti-rat BrdU (1:100, Proteintech Group, Chicago, IL, USA) overnight at 4 °C, followed by a further incubation with the secondary antibody goat anti-rabbit IgG-TRITC (1:50, Proteintech Group) for 60 min at room temperature to detect the cycling cells in the pancreas. 6-Diamidino-2-phenylindole (DAPI, Beyotime, Shanghai, China) was used to stain the nuclei for 10 min. The pictures were captured using Immuno Floure (Olympus, Tokyo, Japan).
Western Blot
Protein extracts of liver were prepared in radio immunoprecipitation assay (RIPA) buffer (50 mM Tris (pH 7.4), 150 mM NaCl, 1% Triton X-100, 1% sodium deoxycholate, 0.1% SDS, and 1 mM phenylmethylsulfonyl fluoride) according to the standard methods (Beyotime, Shanghai, China). Protein concentration was determined using the bicinchoninic acid (BCA) protein assay kit (Pierce Biotechnology, Rockland, ME, USA). About 30 μg of total protein per lane was resolved by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene fluoride (PVDF) membrane. Primary antibody rabbit anti-β-actin antibody (1:10,000, TDY Biotech Co., Ltd, Beijing, China), rabbit anti-GLUT4 antibody (1:1000, Abcam, Cambridge, MA, USA), rabbit anti-p-AKT (1:1000, Cell Signaling Technology, Inc., Danvers, MA, USA), rabbit anti-p-AKT (1:1000, Cell Signaling Technology, Inc.), rabbit anti-p-insulin receptor substrate (IRS)-1 (1:500, Abcam), rabbit anti-IRS-1 (1:1000, Cell Signaling Technology, Inc.), rabbit anti-IL-6 (1:1000, Affbiotech, Changzhou, China), and rabbit anti-TNFα (1:1000, Abcam) were diluted and detected using HRP-goat anti-rabbit IgG (ASPEN, Wuhan, China) and the enhanced chemiluminescent reagent (ECL; Pierce Biotechnology). Immunoreactive bands were detected using Kodak BioMax ML film (Kodak, Rochester, New York, USA). The results were characteristic of at least three independent experiments.
Statistical Analysis
All results are presented as mean ± SEM. Treatments were compared using Student’s unpaired t test or one-way ANOVA with least significant difference post hoc test. A P < 0.05 was considered to be statistically significant.
Results
Characteristics of the High-Fat-Fed Induced Hyperglycemia and Hyperinsulinemia Rat Models
Success of high-fat-fed hyperinsulinemia rat model was confirmed by checking blood glucose, insulin, and proinsulin, respectively (Fig. 1). Four weeks after high fat food, animals were subjected to OGTT. We measured basal blood glucose (normal diet: 4.80 ± 0.58 mM vs high fat diet: 13.23 ± 0.24 mM) as well as blood glucose levels 1 h after oral administration of glucose (normal diet: 7.40 ± 0.12 mM vs high-fat diet: 20.66 ± 0.20 mM) and found that in high-fat diet rats levels were increased by 150% more than normal diet rats (Fig. 1A). Significant insulin (normal diet: 9.57 ± 0.62 ng/ml vs high fat diet: 54.84 ± 0.99 ng/ml) and proinsulin (normal diet: 1.04 ± 0.07 ng/ml vs high fat diet: 2.74 ± 0.09 ng/ml) elevations were also detected (Fig. 1B, C). These data validated that the high-fat diet rats were clinically in early stage of T2DM. Moreover, in these high-fat diet rats, significant deterioration in lipometabolic disorders and abnormal ALT/AST concentration were observed (Fig. 1D, E).

Characteristics of the high-fat-fed hyperinsulinemia rat model. Glucose (A), insulin (B), and proinsulin (C) concentration increased in 4-week high-fat-fed rats with obvious deterioration in lipometabolism (D) and abnormal ALT/AST concentration (E). **P < 0.01 vs control. ALT, alanine aminotransferase; AST, aspartate aminotransferase; LDL, low density lipoprotein; TG, triglyceride; TC, total cholesterol.
BM-MSCs Attenuated the Increased Insulin and Glucose as Well as Restored Lipometabolic Disorder in High-Fat-Fed Rats
Notably tail vein injection of MSCs induced a significant decrease in both insulin and glucose levels in high-fat-fed rats (Fig. 2A, B). Both 105 MSCs and 5 × 105 MSCs significantly attenuated fat-fed induced hyperinsulinemia and hyperglycemia (Fig. 2B, C); 5 × 105 MSCs exerted a greater effect, while incompletely reversed the abnormal OGTT and insulin concentration. Lipometabolic disorders and concentration of AST/ALT were successfully restored to the normal level by 5 × 105 MSCs.

BM-MSCs attenuated changes in glucose/insulin/proinsulin concentration and restored lipometabolism disorder in high-fat-fed rats. MSCs induced a significant decrease in both glucose (A) and insulin/proinsulin concentration (B). Lipometabolic disorders (C) and concentration of AST/ALT (D) were successfully restored and almost to the normal level by 5 × 105 MSCs. **P < 0.01 vs control (I), # P < 0.05 and ## P < 0.01 vs fat-fed (II). ALT, XXX; AST, XXX; BM-MSCs, bone marrow-derived mesenchymal stem cells; LDL, XXX; MSCs, mesenchymal stem cells; TG, XXX; TC, XXX.
BM-MSCs Favored the Histological Structure Recovery in High-Fat-Fed Hyperinsulinemia Rat Model
Fat-fed diet rats had bigger islets (Fig. 3B) than normal diet rats (Fig. 3A) but with irregular morphology and disruption of basement membrane. Lipid accumulation was observed in some islet cells (Fig. 3B). Treatments with 105 MSCs remarkably favored the recovery of the islets and the islet cells. Although abnormal morphology of islets was only partially rescued, lipid in the cells decreased significantly (Fig. 3C). Treatment with 5 × 105 MSCs exhibited better therapeutic effects, where the islets had normal morphology and lipid was almost undetectable in the islet cells (Fig. 3D). Further immunohistochemical staining of BrdU+ cells in the pancreas revealed that BM-MSCs countervailed high-fat-fed induced damage to proliferation ability (Fig. 4).

BM-MSCs favored the histological structure recovery in high-fat-fed rat model. With comparison to control (A), normal diet rats with hyperinsulinemia had irregular islets with lipid accumulation in some islet cells (B). Treatment with 105 MSCs remarkably favored the recovery of the islet and the cells. Although islets remained partially irregular, lipid in the cells decreased significantly (C). Treatment with 5 × 105 MSCs exhibited better therapeutic effects. The islets had normal regular morphology and lipid was almost undetectable in the islet cells (D). BM-MSCs, bone marrow-derived mesenchymal stem cells; MSCs, mesenchymal stem cells.
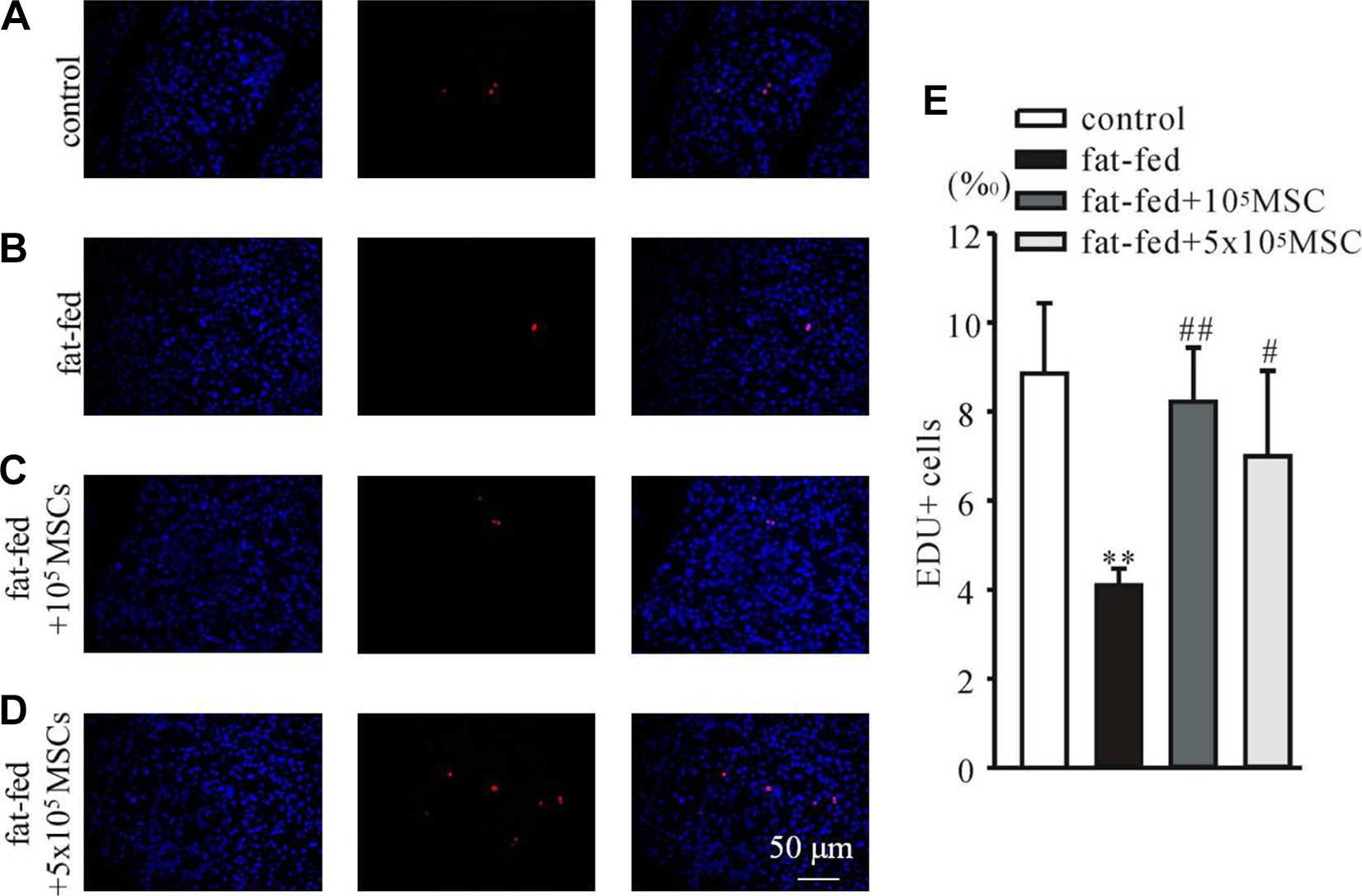
BM-MSCs countervailed high-fat-fed-induced damage to proliferation ability in the pancreas. Immunohistochemical staining of pancreatic sections of control (A), fat-fed (B), fat-fed rat with treatments of 105 MSCs (C) and 5 × 105 MSCs (D) was performed to evaluate the cellular proliferation by detecting BrdU+ (red) cells; nuclei were labeled by DAPI (blue). Statistical data (E) showed that treatments with MSCs significantly countervailed the high-fat-fed induced damage. **P < 0.01 vs control, # P < 0.05 and ## P < 0.01 vs fat-fed. BM-MSCs, bone marrow-derived mesenchymal stem cells; DAPI, 4,6-diamino-2-phenyl indole; EDU, 5-ethynyl-2-deoxyuridine; MSCs, mesenchymal stem cells.
BM-MSCs Reversed the Abnormal Insulin Signaling Transduction and Inflammation in High-Fat-Fed Rat Liver
Protein extracts of liver were used to investigate the influence of BM-MSCs on insulin signaling transduction. High fat-fed rats had less GLUT4 and downregulation of phosphorylated insulin receptor substrate 1 (p-IRS-1) and protein kinase Akt (p-AKT). Administration of BM-MSCs resulted in an increase of GLUT4 expression and enhanced p-IRS-1 and Akt (Fig. 5A–D). Additionally, a high fat-fed induced upregulation of proinflammatory cytokines TNFα and IL-6, which was reversed by BM-MSCs (Fig. 5E–G). These data suggest that BM-MSCs can effectively potentiate the transduction of insulin signaling and inhibit the inflammation in insulin target tissues.
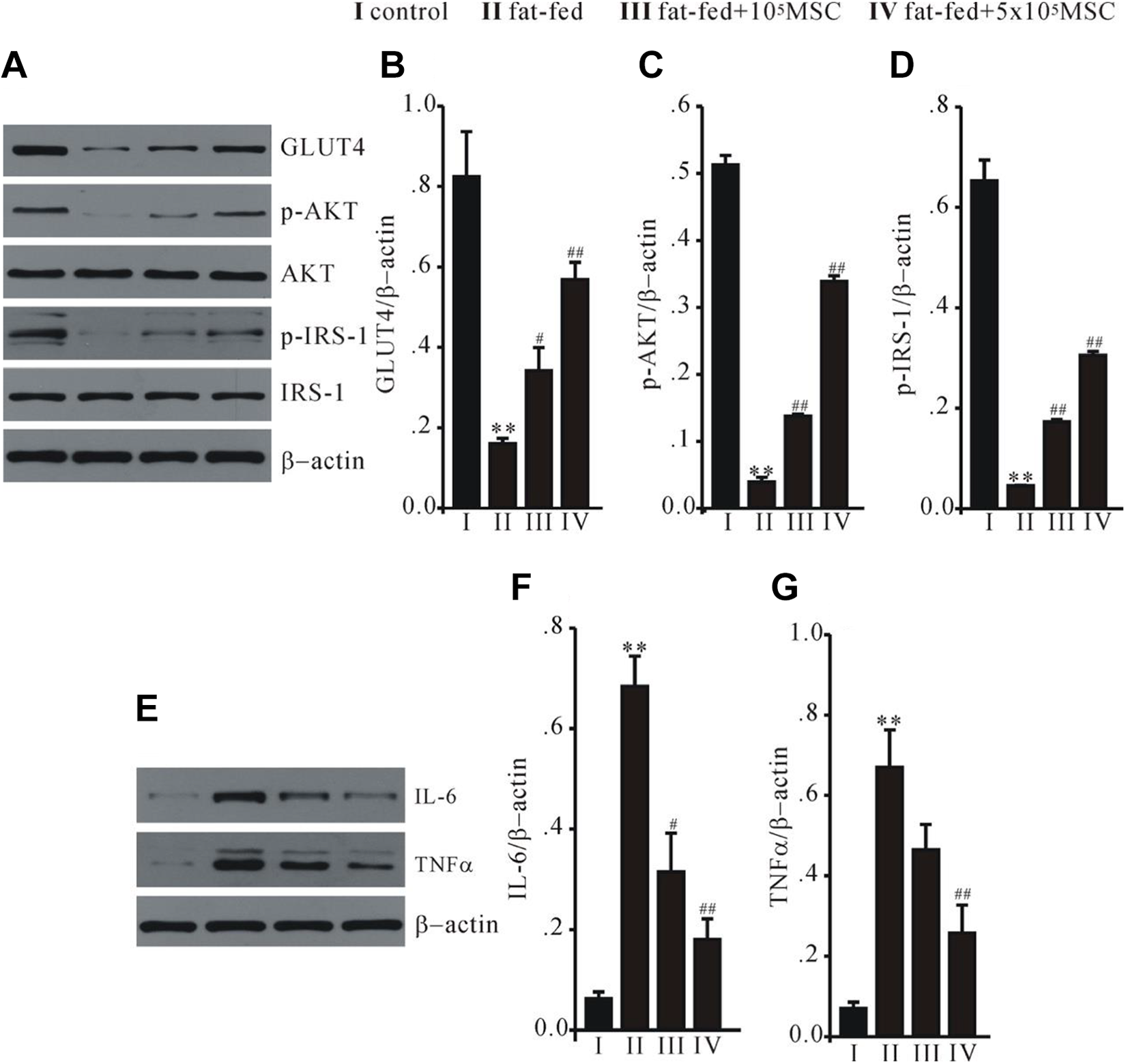
BM-MSCs reversed insulin signaling transduction and inflammation in a high fat-fed rat liver. Western blot analysis on GLUT4, p-IRS-1, and p-AKT (A), and proinflammatory cytokines IL-6 and TNFα (E). Statistical data (B–D, F–G) revealed that administration of MSCs successfully reversed the abnormal expression of these proteins in fat-fed rat model. **P < 0.01 vs control, # P < 0.05 and ## P < 0.01 vs fat-fed. BM-MSCs, bone marrow-derived mesenchymal stem cells; MSCs, mesenchymal stem cells.
Discussion
A high-caloric diet has been broadly characterized as the trigger of T2DM28–30, and T2DM accounts for 90%–95% of all DM cases, IR being the typical symptom and mechanism at the early stage of dietary-induced T2DM4–6,31. High-fat-induced IR could generally associate with alterations in lipid cellular intake and accumulation, followed by disorders of the metabolism of β-cells, stroke, and other diseases32,33. Here in our model, the high fat diet for 4 wk successfully induced hyperinsulinemia and elevation of blood glucose concentration, which was associated with lipometabolic disorders and rising ALT/AST. With this model, we showed that the intervention of MSCs at early stage of T2DM could significantly lead to decline of insulin/glucose as well as rescue lipometabolic disorders and liver dysfunction. A much lower cell dose (5 × 105; vs 2 × 106 cells in typical T2DM model)22,24 induced remarkable effects, which suggested an economic time window for application of MSCs in T2DM.
MSCs harbor great potential to become a routine therapeutic measure for T2DM, partially due to reversing IR. Previous studies have shown that infusion of MSCs ameliorates hyperglycemia by alleviating IR in T2DM rats23–25. Consistently, we found that in vivo transplantation of BM-MSCs attenuated increase in insulin and glucose resulting from a high-fat diet. It was notable that such therapeutic potential was not observed in a typical T2DM model induced by both high-fat diet and injection of streptozotocin 26 , but in a high-fat diet triggered model. This strongly suggests early transplantation of MSCs could serve as a better strategy than have been proposed by previous studies to restore pancreatic or multiple organ dysfunction at later stage of T2DM33,34.
IR could produce hyperinsulinemia, this in turn induces multiple organic dysfunction due to an abnormal intake of lipid and lipid accumulation in cells28,32. We found that MSCs restored lipometabolic disorders and liver dysfunction, as evidenced by the concentration of LDL/TG/TC and AST/ALT. More importantly, high-fat diet caused lipid accumulation in islet cells and disruption of the islet basement membrane; BM-MSCs favored the histological structure recovery, and obviously improved the proliferation potential of islet cells. These findings indicated that the potential risk of T2DM for other organs as well as the pancreas could be prevented if MSC transplantation is exerted at early stage of T2DM.
GLUT4 and phosphorylation of IRS-1 (p-IRS-1) and AKT (p-AK) are crucial for conferring insulin-signaling transduction, and glucose uptake therefore related intensively to IR35,36. Here in our high fat-fed induced IR model, BM-MSCs successfully enhanced GLUT4, p-IRS-1, and p-AKT, which is similar to the findings in typical T2DM model with MSC treatment 24 . A lot of evidence has shown that chronic activation of proinflammatory pathways within insulin target cells could lead to IR 37 . We found that BM-MSC remarkably reversed the upregulation of IL-6 and TNFα in the liver. These observations suggested that both the insulin-signaling pathway and proinflammatory pathways are involved in the favorable function of BM-MSCs in high fat-fed induced IR.
Due to the increase in associated risk factors, such as being overweight or obese, the global prevalence of (age-standardized) DM has been rising dramatically and it might become reality that 1 adult in every 10 will have diabetes in 204029,30. Our study proposed a new strategy with MSC-based cell therapy for T2DM, i.e., MSC transplantation during hyperinsulinemia before onset of diabetes could prevent most possible risks of T2DM for patients.
It is the limitation of our study that we did not go further to investigate the detailed mechanisms for MSCs to reverse IR in the early stage of T2DM. The widely accepted idea is that MSC infusion with host cells is one of the possibilities 24 . This could also be the underlying phenomenon in our study. Future work is necessary to validate it and to explore other possible mechanisms.
Footnotes
Data Availability
All the data used to support the findings of this study are included within the article files.
Ethical Approval
Ethical approval to report this case was obtained from Tongji Medical College, Huazhong University of Science and Technology Institutional Animal Care and Use Committee (Approval: 2016IACUC number, 644).
Statement of Human and Animal Rights
All procedures in this study were conducted in accordance with the Tongji Medical College, Huazhong University of Science and Technology Institutional Animal Care and Use Committee (Approval: 2016IACUC number, 644) approved protocols.
Statement of Informed Consent
There are no human subjects in this article and informed consent is not applicable.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: We gratefully acknowledge funding from the National Natural Science Foundation of China (NSFC, 81801922) and Chronic Wound & Diabetes Medical Clinical Research Center Foundation of Hubei provincial China (2018BCC340).