
Abstract
Isolated primary hepatocytes, which are widely used for pharmacological and clinical purposes, usually undergo certain periods of cold storage in suspension during processing. While adherent hepatocytes were shown previously to suffer iron-dependent cell death during cold (4 °C) storage and early rewarming, we previously found little iron-dependent hepatocyte death in suspension but severely decreased attachment ability unless iron chelators were added. Here, we focus on the role of mitochondrial impairment in this nonattachment of hepatocyte suspensions. Rat hepatocyte suspensions were stored in a chloride-poor, glycine-containing cold storage solution with and without iron chelators at 4 °C. After 1 wk of cold storage in the basic cold storage solution, cell viability in suspension was unchanged, while cell attachment was decreased by >80%. In the stored cells, a loss of mitochondrial membrane potential (MMP), a decrease in adenosine triphosphate (ATP) content (2 ± 2 nmol/106 cells after cold storage, 5 ± 3 nmol/106 cells after rewarming vs. control 29 ± 6 nmol/106 cells), and a decrease in oxygen consumption (101 ± 59 pmol sec−1 per 106 cells after rewarming vs. control 232 ± 83 pmol sec−1 per 106 cells) were observed. Addition of iron chelators to the cold storage solution increased cell attachment to 53% ± 20% and protected against loss of MMP, and cells were able to partially regenerate ATP during rewarming (15 ± 10 nmol/106 cells). Increased attachment could also be achieved by addition of the inhibitor combination of mitochondrial permeability transition, trifluoperazine + fructose. Attached hepatocytes displayed normal MMP and mitochondrial morphology. Additional experiments with freshly isolated hepatocytes confirmed that impaired energy production—as elicited by an inhibitor of the respiratory chain, antimycin A—can decrease cell attachment without decreasing viability. Taken together, these results suggest that mitochondrial impairment with subsequent energy deficiency is a key factor for the lack of attachment of cold-stored hepatocyte suspensions.
Introduction
Isolated primary hepatocytes, which are widely used for pharmacological and toxicological research, usually undergo at least brief periods of cold storage in suspension at 2 to 8 °C during processing and, possibly, during transport.
However, it is known from adherent hepatocytes that low temperature, which is used to protect the cells by slowing down metabolic processes, also induces cell damage by itself. 1 –3 The largest proportion of this injury is caused by an increase in intracellular chelatable iron ions of yet unknown sources, 4 –6 leading to the formation of highly reactive oxygen species. 1,5 This increase in chelatable iron ions already occurs within the first hour of cold incubation. In adherent hepatocytes, the iron-dependent pathway of cold-induced injury is known to target primarily the mitochondria and induce mitochondrial permeability transition, 1,6 finally leading to apoptosis during rewarming. 2,7 Addition of iron chelators effectively inhibits this pathway of cold-induced injury.
Cold storage of hepatocyte suspensions is usually performed in buffered salt solutions, cell culture medium, or in organ preservation solutions. 8 –11 In previous experiments, we showed largely improved rat hepatocyte storage in a chloride-poor modification of the tissue preservation solution TiProtec, 12,13 which contains, besides other components, iron chelators for protection against iron-dependent cold-induced cell injury and the small amino acids glycine and alanine. In contrast to cold storage in cell culture medium and organ preservation solutions, hepatocyte viability was mainly unchanged after 1 wk of cold storage in this modified TiProtec solution, both with and without iron chelators, and did not notably decrease during early rewarming. 14 We showed that the protective effect of this solution on viability could be attributed mainly to the presence of glycine and alanine (in higher concentrations than in cell culture medium) during cold storage. Surprisingly, however, in spite of the mainly unchanged viability in the presence of glycine and alanine, cell attachment was significantly decreased after cold storage in solutions without iron chelators. This lack of attachment makes the cells unavailable for classical cell culture experiments and supposedly might also lead to diminished engraftment in cell transplantation settings.
A pathway that leads to the reduced attachment in the absence of iron chelators has not yet been described. We hypothesized that (iron-dependent) sublethal damage to the mitochondria impairs ATP regeneration during the early rewarming phase, which in turn prevents processes—for example, of cytoskeletal rearrangement—necessary for cell attachment. Therefore, we here focused on the role of mitochondria and energy state in the cold-induced loss of the attachment ability of hepatocytes.
Materials and Methods
Chemicals
The basic cold storage solution used in the experiments is a chloride-poor variant (8.1 mM Cl−, 2 mM α-ketoglutarate, 5 mM aspartate, 95 mM lactobionate, 1 mM
Animals
For hepatocyte isolation, male Wistar rats (250 to 350 g) were obtained from the Central Animal Facility of the Universitätsklinikum Essen. Animals received humane care in compliance with the German Law for the Protection of Animals and following institutional guidelines. Permission for liver cell isolation was obtained from local authorities (Landesamt für Natur, Umwelt und Verbraucherschutz NRW, Az. 84-02.05.20.12.007 and Az. 84-02.04.2015.A485).
Cell Isolation and Culture
Hepatocytes were isolated by in situ collagenase digestion (50 U/L collagenase NB 4G, Serva Electrophoresis, Heidelberg, Germany) and further purification steps as described earlier. 2,14 Viability after isolation was routinely determined by trypan blue exclusion to normalize viable cell number to 106 cells for seeding/cold storage. Adherent control cultures were obtained as described earlier, 14 seeding 106 viable cells per well onto collagen-coated 6-well plates in supplemented Leibovitz L-15 cell culture medium. After 2 h, cell cultures were washed 3 times with warm Hanks balanced salt solution (HBSS) and supplied with 2 mL of fresh medium.
Cold Storage of Cell Suspensions and Culture of Cold-stored Cells
Cell suspensions were stored in basic cold storage solution or complete cold storage solution (with iron chelators, see above) with or without a combination of 20 µM trifluoperazine (TFP) + 10 mM fructose 15 (to inhibit mitochondrial permeability transition) or in organ preservation solutions (University of Wisconsin [UW] solution, histidine–tryptophan–ketoglutarate [HTK] solution, Institute Georges Lopez-1 [IGL-1] solution) or in cell culture medium (Leibowitz L-15). For cold storage, 106 viable cells/mL were added to the respective precooled cold storage solution and stored horizontally in 1.8 mL cryovials at 4 °C. After cold incubation, 1 mL of the respective cell suspension was seeded onto collagen-coated 6-well plates containing 2 mL of prewarmed supplemented cell culture medium. After 2 h, cells were washed as described previously.
Rewarming in Suspension
For rewarming in suspension, 1 mL of the cold-stored cell suspension was added to 2 mL supplemented cell culture medium in 5 mL round-bottom tubes (Greiner Bio-One, Frickenhausen, Germany). Tubes were incubated upright at 37 °C and 5% CO2.
Quantification of Cell Attachment and Metabolic Activity
After 24 h of culture, attached viable cells were quantified by determination of intracellular lactate dehydrogenase (LDH). Cell cultures were washed 3 times with HBSS and lysed with 1% triton X-100 in HBSS. LDH activity in the cell lysate was determined using a standard nicotinamide adenine dinucleotide (NADH)-based assay and represents the number of adherent viable cells (i.e., without LDH leakage). Metabolic activity was assessed by resazurin conversion as described earlier. 16
Determination of Mitochondrial Membrane Potential (MMP) and Viability in Suspension
For assessment of viability, cells were stained with 5 µg/mL propidium iodide (PI) for 10 min at 4 °C or 2 min at 37 °C. To quantify MMP, cells were stained with 500 nM tetramethyl rhodamine methyl ester (TMRM) overnight at 4 °C (controls: 20 min at 37 °C). Red fluorescence was recorded by flow cytometry (accuri C6 flow cytometer; λexcitation = 488 nm, λemission > 670 nm). TMRM-positive (TMRM++) cells were defined using a fix threshold for all measurements deduced from clearly TMRM-positive control cells. Cells with a red fluorescence higher than unstained cells but below this threshold were defined as cells with a reduced MMP (TMRM+).
Fluorescence Microscopy
For visualization of viability and MMP in rat hepatocyte suspensions, cells were stained with TMRM (red), PI (5 μg/mL, red stain of nuclei of dead cells), and Hoechst 33342 (1 µg/mL; blue stain of all nuclei) for 20 min at 37 °C (control cells) or overnight at 4 °C. Part of the cold-stored suspension was then rewarmed at 37 °C for 1 h as described above. Nonstored control suspensions from the same cell isolations were treated identically. Fluorescence micrographs of thin layers of the suspensions spread out on cover slips were taken directly after isolation, after cold storage, and after 1 h at 37 °C with a Zeiss Axio Observer.Z1 at λexc. = 546 ± 6 nm/λem. ≥ 590 nm for red fluorescence and λexc. = 359 ± 24 nm/λem. = 445 ± 25 nm for blue fluorescence (Zeiss Axio Observer.Z1 with ApoTome module).
For determination of MMP in early attached cells, cells were seeded onto collagen-coated glass cover slips at 1.1 × 105 initially living cells/square centimeter after staining with TMRM, and fluorescence micrographs of attached cells were taken after 1 h. Fully adherent rat hepatocyte cultures after 24 h were stained with TMRM at the end of culture time.
ATP Content
At the respective time points, perchloric acid (9 M) was added to samples of the cell suspension to a final concentration of 1 M and the thus treated suspensions were frozen immediately in liquid nitrogen (both treatments to stop adenosine triphosphate (ATP) degradation). Samples were then kept at −80 °C until measurement. After slow thawing and centrifugation at 160× g, the supernatant was taken, pH adjusted to 7.1 to 7.7 with 5 M potassium hydroxide, samples were centrifuged and ATP concentration was determined in the supernatants by a coupled enzymatic assay with hexokinase and glucose-6-phosphate-dehydrogenase after Trauschold et al. 17 using a dual-beam photometer (UV/VIS Spectrometer Lambda 40 with UV WinLab software version 2.7, PerkinElmer, Baesweiler, Germany).
Oxygen Consumption
For controls, maximum stable oxygen consumption at 37 °C was determined in 2 mL of cell suspensions in Krebs–Henseleit buffer (106 viable cells/mL) directly after cell isolation using high-resolution respirometry (Oroboros Oxygraph-2k, Oroboros Instruments, Innsbruck, Austria). After cold incubation, 2 mL of cell suspension (containing 2 × 106 initially viable cells) in the respective cold storage solution were added to a precooled chamber of the oxygraph and rewarmed in the instrument. Maximum stable oxygen consumption at 37 °C was recorded.
Inhibition of the Mitochondrial Respiratory Chain
To confirm the influence of impaired respiratory capacity on cell attachment, we used the respiratory chain inhibitor antimycin A in freshly isolated (i.e., nonstored) rat hepatocytes. Cell suspensions (106 viable cells/mL in Krebs–Henseleit buffer) 14 were incubated without inhibitor, with solvent control or with 2.5 µM antimycin A for 30 min at 37 °C in 5 mL round-bottom tubes. After incubation, part of the cells were seeded onto collagen-coated 6-well plates containing 2 mL of prewarmed supplemented cell culture medium at a cell density of 106 initially viable cells per well. After 1 h at 37 °C and 5% CO2, attached viable cells were quantified by intracellular LDH as described above. Part of the cells were incubated for another 60 min in suspension after addition of supplemented cell culture medium (2:1) to the incubation buffer. Viability and oxygen consumption in suspension were determined after 30 min incubation with antimycin A and again after further 60 min as described above.
Statistics
Experiments were performed 4 to 14 times with cells from different cell isolations as indicated in the respective figure legends. Data are expressed as median with 25/75 percentiles in box plots (whiskers represent 1.5 times the interquartile range, outliers are marked as dots), if not indicated otherwise. Within each experimental set, pairing by cell isolation was used to limit the influence of interexperiment variance. Comparison of the different groups was done by Friedman test followed by post hoc Dunn’s multiple comparison test. P ≤ 0.05 was considered as statistically significant.
Results
Cell Viability after Cold Storage
After 48 h of cold storage, cell viability was unchanged in suspensions stored in cell culture medium or cold storage solutions, except for a significant decline in viability after cold storage in HTK solution (Fig. 1A), although different cell attachment could be observed in the different solutions (Fig. 1B). After 1 wk of cold storage in basic cold storage solution or in cold storage solution with iron chelators, still no major change in cell viability occurred (Fig. 2A), confirming previous results. 14 Similarly, in basic cold storage solution with the inhibitors of mitochondrial permeability transition TFP (20 µM) + 10 mM fructose, viability did not change. Also, after 1 h of rewarming in suspension, only a slight, but significant, decrease in viability occurred in cells stored in basic cold storage solution compared to cells stored in cold storage solution with iron chelators (Fig. 2B).

Cell viability and cell attachment after 48 h of cold storage. Suspensions of isolated rat hepatocytes (106 cells/mL) were stored for 48 h at 4 °C in cell culture medium (L-15), cold storage solution with and without iron chelators (basic solution, + chelators), or the organ preservation solutions University of Wisconsin (UW) solution, histidine–tryptophan–ketoglutarate (HTK) solution, or Institute Georges Lopez-1 (IGL-1) solution. The proportion of living (propidium iodide-negative) cells was determined by flow cytometry after cell isolation (nonstored; control) or directly after cold storage (A). In parallel, cold-stored cells were seeded onto collagen-coated 6-well plates in L-15 cell culture medium without further purification steps. After 24 h of culture, adherent viable cells were quantified by the amount of intracellular lactate dehydrogenase. Intracellular lactate dehydrogenase under experimental conditions is given as percentage of the respective nonstored control cells (B). Friedman test; *P < 0.05, n = 5.

Cell viability after 1 wk of cold storage and rewarming. Suspensions of isolated rat hepatocytes (106 cells/mL) in basic cold storage solution, complete cold storage solution (with iron chelators), or basic solution with 20 µM trifluoperazine (TFP) + 10 mM fructose were stored at 4 °C for 1 wk. Part of the cold-stored suspension was then rewarmed at 37 °C for 1 h after adding 1 part of the original cell suspension in the respective cold storage solution to 2 parts of completed cell culture medium. Nonstored control cells directly after isolation, cold-stored cell suspensions, and rewarmed cell suspensions were stained with propidium iodide (PI; 2 min at 37 °C or 10 min at 4 °C, respectively). The proportion of living (PI-negative) cells was determined by flow cytometry directly after storage (A) and after 1 h rewarming (B). Friedman test; **P < 0.01, n = 10.
Cell Attachment
After 48 h of cold storage in standard cell culture medium, cell attachment was markedly decreased (Fig. 1B), while no loss of attachment ability was observed after 48 h cold storage in the new cold storage solution with or without iron chelators compared to nonstored control cells. In contrast, 48 h of cold storage in the organ preservation solutions UW, HTK, or IGL-1 resulted in significantly lower attachment after cold storage compared to the new solutions (Fig. 1B).
After 1 wk of cold storage in standard cell culture medium, hardly any cells (3% ± 3% compared to attachment of nonstored control cells) attached to the cell culture surface after seeding. After cold storage in basic cold storage solution, attachment increased to 17% ± 16% (Fig. 3A). This attachment could not be improved by prolonging the attachment period to 4 h (data not shown). As we already showed earlier, 14 after 1 wk of cold storage, the presence of iron chelators significantly improved attachment rates, resulting in adherent cell cultures of a viable cell density amounting to 59% ± 23% of viable cell density of nonstored control cultures (Fig. 3A, B). Addition of TFP + fructose, a strong inhibitor combination of mitochondrial permeability transition, 15 to the basic solution also significantly improved attachment (compared to the solvent control; Fig. 3A). No additive effect was observed with chelators and TFP + fructose, but attachment rate slightly decreased to the level of TFP + fructose only (37% ± 15% vs. solvent control with chelators 47% ± 17%; difference not significant).

Cell attachment and metabolic activity after 1 wk of cold storage. Suspensions of isolated rat hepatocytes (106 cells/mL) were stored in basic cold storage solution, complete cold storage solution (with iron chelators), and basic cold storage solution supplemented with the inhibitor combination of mitochondrial permeability transition, 20 µM trifluoperazine (TFP) + 10 mM fructose, at 4 °C for 1 wk. Ethanol (+ EtOH) was added as solvent control for TFP/fructose. Following cold storage, cells were seeded onto collagen-coated 6-well plates in L-15 cell culture medium without further purification steps. After 24 h of culture, adherent viable cells were quantified (A; intracellular lactate dehydrogenase, given as percentage of the respective nonstored control cells), phase contrast micrographs were taken (B; control cells without cold storage, basic solution, and complete solution only; original magnification 100×), and metabolic activity was determined and is given as percentage of activity of the respective nonstored control cells (C; resazurin reduction). Friedman test; *P < 0.05, **P < 0.01, ***P < 0.001, n = 14 for A and C.
Metabolic Activity
Similar to the significant protective effect of the addition of iron chelators on metabolic activity, which we already showed earlier, 14 addition of TFP + fructose also significantly increased metabolic activity (Fig. 3C).
MMP after Cold Storage and after Rewarming
In control suspensions, living cells (PI-negative nuclei; blue) showed largely normal MMP, although a certain heterogeneity, as expected in primary hepatocyte suspensions, could be observed (Fig. 4A). Dead cells (PI-positive nuclei; red) more or less completely lost their MMP. After 1 wk of cold storage, a fraction of cells with greatly reduced MMP could be observed in both solutions (PI-negative nuclei but reduced TMRM fluorescence; Fig. 4B, C, see arrowheads). During 1 h of rewarming, cells stored in cold storage solution without iron chelators tended to further lose their MMP (Fig. 4D), while MMP appeared to be better preserved during rewarming after cold storage in cold storage solution with iron chelators (Fig. 4E).

Mitochondrial membrane potential of cells in suspension. Suspensions of isolated rat hepatocytes (nonstored control, A) were stored at 106 cells/mL in basic cold storage solution (B, D) or complete cold storage solution (with iron chelators; C, E) at 4 °C for 1 wk. Part of the suspension was then rewarmed at 37 °C for 1 h after adding 1 part of the original cell suspension in the respective cold storage solution to 2 parts of completed cell culture medium. Cell suspensions were stained with tetramethylrhodamine methyl ester (TMRM, indicator for mitochondrial membrane potential [MMP], red; control cells: 20 min at 37 °C; cold-stored suspensions: overnight at 4 °C), propidium iodide (PI; nuclei of dead cells, red; 2 min at 37 °C or 10 min at 4 °C), and Hoechst 33342 (nuclei, blue; 20 min at 37 °C, cold-stored suspensions: overnight at 4 °C). Arrowheads indicate living cells (PI negative, blue nuclei) that lost their MMP (absence of red TMRM outside nuclei), asterisks indicate dead cells (PI-stained nuclei).
Quantification of MMP, as performed by flow cytometry, showed that control cells kept in suspension for 30 min and 1 h (at 37 °C) did not lose MMP during this time (Fig. 5A). Cell stored in basic cold storage solution for 1 wk displayed a slightly decreased fraction of cells with MMP (TMRM+/TMRM++), and within this group a decrease in the fraction with full membrane potential (TMRM++) and an increase in the fraction of cells with reduced MMP (TMRM+) compared to control cells. During rewarming, the fraction of cells with full MMP further decreased, while the number of cells with reduced MMP further increased (Fig. 5A). Cells stored in complete cold storage solution (i.e., with iron chelators) largely preserved their MMP (TMRM+/TMRM++), although the fraction of cells with reduced MMP (TMRM+) also increased over time (Fig. 5A). The percentage of cells with full MMP (TMRM++) after cold storage and after 1 h rewarming, as summarized in Table 1 (for a higher number of experiments), differed significantly between cold incubation with and without iron chelators.

Mitochondrial membrane potential (MMP) after cold storage and during rewarming. Suspensions of isolated rat hepatocytes (106 cells/mL) were stored in basic cold storage solution or complete cold storage solution (with iron chelators) at 4 °C for 1 wk. Part of the suspension was then rewarmed at 37 °C for 1 h after adding 1 part of the original cell suspension in the respective cold storage solution to 2 parts of completed cell culture medium. One aliquot was seeded onto collagen-coated 6-well plates. Nonstored control suspensions from the same cell isolation were treated similarly (1 h at 37 °C in suspension with 2 parts of cell culture medium added/seeded onto 6-well plates). Cells were stained with tetramethylrhodamine methyl ester (TMRM, controls: 20 min staining with TMRM at 37 °C, cold-stored cells overnight at 4 °C). MMP in cell suspensions (A) and the supernatant of the plates (i.e., nonattached cells; B) was determined by flow cytometry directly after isolation (control) or after cold storage (basic cold storage solution/complete cold storage solution) and after 30 min and 1 h incubation at 37 °C. Cells were classified as having normal MMP (TMRM++, black bottom portion of bars with white standard deviation [SD] below), reduced MMP (TMRM+, white middle portion with SD above), or no MMP (TMRM−, top gray portion with SD above); n = 5.
Cells with Full Mitochondrial Membrane Potential after Cold Storage and Rewarming in Suspension.

Note: Suspensions of isolated rat hepatocytes (106 cells/mL) were cold stored in cold storage solution with or without iron chelators for 1 wk at 4 °C and stained with tetramethylrhodamine methyl ester (TMRM; indicator for mitochondrial membrane potential) overnight. Part of the suspensions were rewarmed (37 °C) for 1 h after addition of cell culture medium to the cold storage solution (2:1). Cells with normal mitochondrial membrane potential as assessed by flow cytometry are given as percentage of total cells (mean ± standard deviation).
aSignificantly different from control.
bSignificantly different from basic solution at given time point; n = 9.
A general reduction in the fraction of TMRM++ cells could be observed in the supernatant on cell culture plates (i.e., nonattached cells) during 1 h of attachment, despite the only moderate change in the fraction of TMRM++ cells kept in suspension for the entire period (cf. Fig. 5A and B), suggesting attachment of TMRM++ cells. More TMRM++ cells in the supernatant were found after cold storage in basic and complete cold storage solution than in nonstored control cultures after 30 min attachment time (Fig. 5B). However, after an attachment period of 1 h, the difference between nonstored cells and cells after cold storage was no longer pronounced, suggesting slower attachment after cold storage.
Adherent control cells displayed normal mitochondrial morphology and an intact MMP after 1 h of attachment (Fig. 6A) and after 24 h of culture (Fig. 6D). Mitochondrial morphology and MMP were also unchanged in cells that were able to attach after cold storage in either solution (1 h of attachment, Fig. 6B, C and 24 h of culture, Fig. 6E, F).

Mitochondrial membrane potential (MMP) and mitochondrial morphology in adherent cells after cold storage. Suspensions of isolated rat hepatocytes in basic cold storage solution (B, E) or complete cold storage solution (with iron chelators; C, F) were stored at 106 cells/mL and 4 °C for 1 wk. Part of the suspensions were stained with tetramethylrhodamine methyl ester (TMRM; indicator for MMP) overnight at 4 °C and seeded in collagen-coated 6-well plates at 106 initially living cells per well. Nonstored control suspensions from the same cell isolations were treated similarly (20 min staining with TMRM; seeding on collagen-coated 6-well plates at 106 living cells/well; A). Fluorescence micrographs were taken after 1 h (A–C). In parallel, nonstained cells were seeded, cell cultures were stained with TMRM for 20 min at 37 °C after 24 h of culture (control [D], after cold storage [E, F]).
ATP Levels after Cold Storage and Rewarming
Since the results presented above suggest an impairment of mitochondrial function after cold storage, we determined ATP levels in cell suspensions. Control cells contained 29 ± 6 nmol ATP per 106 cells (Fig. 7A). After cold storage, ATP levels were significantly decreased in both cold storage solution with and without iron chelator. After cold storage in the basic solution, ATP levels remained low after 30 min of rewarming, while during rewarming after cold storage in the presence of iron chelators, a considerable ATP regeneration could be observed, although values did not reach the level of nonstored control cells (Fig. 7B).

Cellular ATP content after cold storage/rewarming. Suspensions of isolated rat hepatocytes (106 cells/mL) in basic cold storage solution or complete cold storage solution (with iron chelators) were stored at 4 °C for 1 wk. Part of the cold-stored suspension was then rewarmed at 37 °C for 30 min after adding 1 part of the original cell suspension in the respective cold storage solution to 2 parts of completed cell culture medium. ATP content of nonstored control cells and cold-stored cells directly after cold storage (A) and after 30 min of rewarming (B) was determined. Friedman test; *significantly different from control cells (P < 0.05, n = 7).
Oxygen Consumption
Control cells displayed a maximum stable oxygen consumption of 232 ± 83 pmol/s/106 cells. Oxygen consumption of cells stored in basic cold storage solution was significantly decreased during rewarming compared to nonstored control cells (Fig. 8). Although oxygen consumption was still markedly decreased during rewarming after cold storage in the presence of iron chelators, the difference was not significant compared to the control.

Oxygen consumption after cold storage. Suspensions of isolated rat hepatocytes (106 cells/mL) in basic cold storage solution or complete cold storage solution (with iron chelators) were stored at 4 °C for 1 wk and then rewarmed to 37 °C in the chamber of a high-resolution respirometer. Maximum stable oxygen consumption at 37 °C was determined and compared to oxygen consumption of the respective nonstored control cells at 37 °C. Friedman test; ***P < 0.001, n = 10.
Influence of Interference with Mitochondrial Energy Production on Cell Attachment
To confirm the influence of sublethal mitochondrial impairment on cell attachment, we used the complex III inhibitor antimycin A in freshly isolated rat hepatocytes. The inhibitor concentration was chosen to inflict sublethal damage, while severely inhibiting cell respiration. Viability was slightly decreased after 30 min incubation with 2.5 µM antimycin A compared to both initial viability and solvent control (Fig. 9A), and, as expected, oxygen consumption was decreased to about one-third after incubation with antimycin A (30% ± 9% of that of cells incubated without antimycin A, solvent control: 89% ± 5%). Although cell damage was not lethal (as intended), cell attachment was significantly decreased (Fig. 9B).
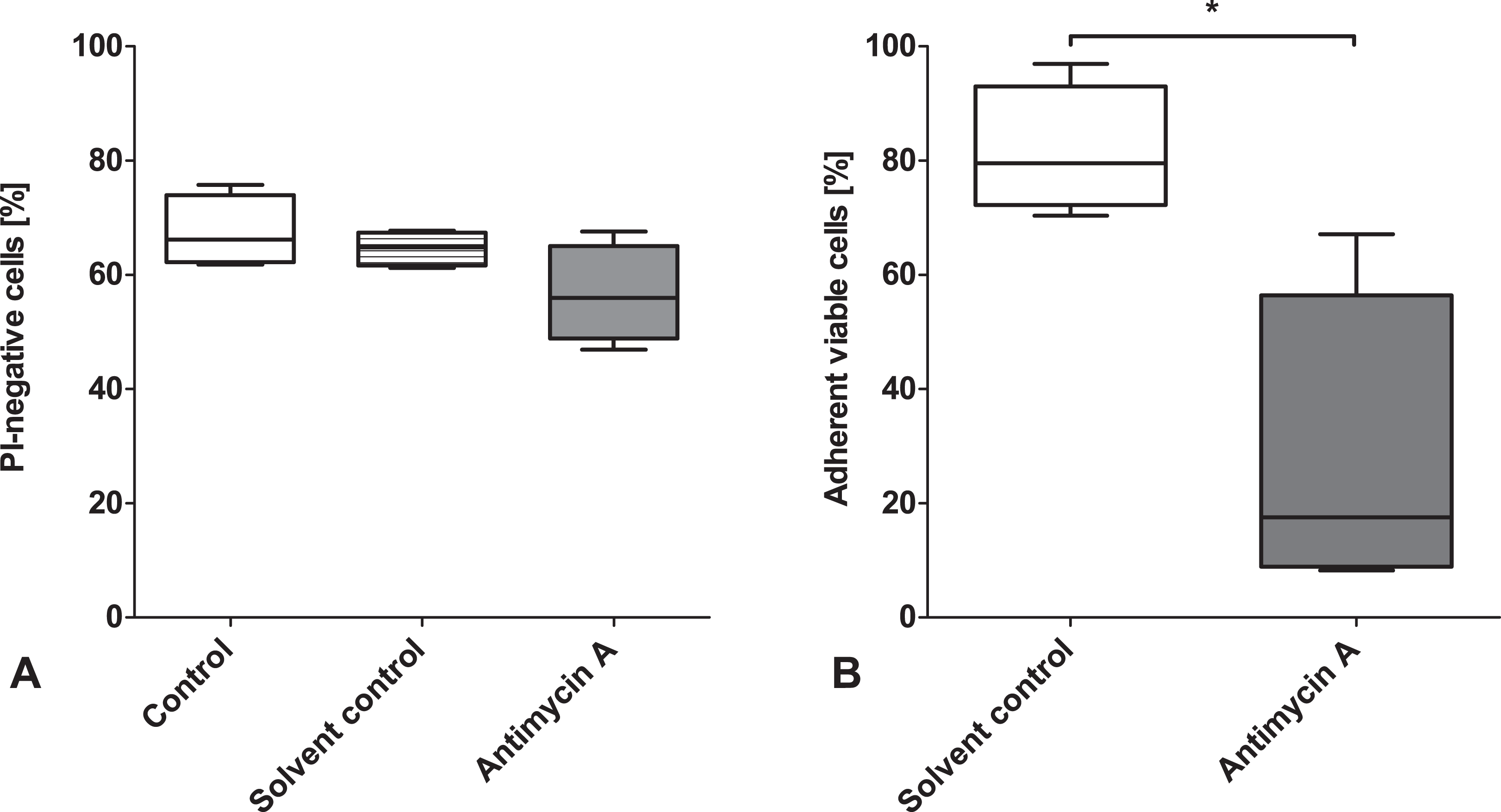
Influence of respiratory chain inhibition on attachment in freshly isolated rat hepatocytes. Nonstored rat hepatocyte suspensions (106 cells/mL) in Krebs–Henseleit buffer with and without 2.5 µM antimycin A (solvent control: 1% ethanol) were incubated for 30 min at 37 °C. Afterwards, cell viability (propidium iodides [PI] exclusion) was determined by flow cytometry (A) and cells were seeded in wells of collagen-coated 6-well plates containing 2 mL supplemented cell culture medium. Cultures were washed after 1 h and cell attachment was quantified (B; determination of intracellular lactate dehydrogenase activity, given as percentage of nontreated control cultures).
Discussion
The results of our study suggest that the main reason for nonattachment of rat hepatocytes after cold storage is energy deficiency caused by cold-induced mitochondrial impairment, which results in reduced MMP (Figs. 4 and 5A), decreased O2 consumption (Fig. 8), and severely impaired ATP regeneration capacity (Fig. 7) during/after rewarming. This mitochondrial impairment is partially inhibited and attachment increased by the addition of iron chelators during cold storage.
Cell Viability after Cold Storage
As we showed earlier, no major changes in cell viability occurred after cold storage in the solutions containing glycine and alanine. 14 These small amino acids are widely known to inhibit not (aerobic) cold-induced but hypoxic (or energy deficiency-induced) cell damage, 18,19 which might, due to increased diffusion distances and higher local cell density (sedimentation), play a greater role in cold storage of cell suspensions than in monolayers. However, our suspension storage conditions (low cell density, horizontal storage of tubes) were chosen to ensure aerobic conditions. That these conditions are indeed largely aerobic could be confirmed in our previous study, 14 in which marked lipid peroxidation (requiring oxygen) was observed during cold storage of the hepatocyte suspensions. Aerobic conditions were confirmed in the current study, as the mean pO2 of cell suspensions after 1 wk of cold storage at 37 °C was >80 mmHg (corresponding to >45 mmHg at 4 °C; data not shown). However, we cannot exclude that pericellular oxygen availability in the suspended cells is somewhat lower than in attached cells.
Although the setting was largely aerobic, ATP levels were low at the end of cold storage (Fig. 7A), and our previous study 14 had shown that hepatocytes died after cold storage in cold storage solution without alanine and glycine . These amino acids have been shown to inhibit the alterations in cellular ion homeostasis subsequent to energy deficiency 18,19 but do not increase cellular ATP levels. 18 –20 Protection of cold-stored hepatocytes by glycine has also been shown by Marsh et al., although their model (sedimented cell suspensions, nitrogen equilibration of the solution) was designed to simulate cold ischemia, 21,22 that is, encompasses hypoxia as a decisive factor, while in the current model, hypothermia itself is likely to contribute decisively to energy deficiency.
Cell Attachment after Cold Storage
Cell attachment was already severely impaired by 48 h cold storage in cell culture medium and especially in organ preservation solutions (Fig. 1B) but not in the new solutions (basic solution and solution with iron chelators). This decreased attachment in the established solutions was observed despite a mainly unchanged viability in all but one of these solutions (i.e., HTK solution). This decline in (cell viability and) cell function after 48 h of cold storage is in line with previous findings 8,11,23 and the superiority of the new solution is also in accordance with earlier results. 14
In contrast to other publications, 9,11,24 where only viable cells are seeded, the nonpurified cell suspension was used for attachment experiments in this study. Therefore, the combined effect of cell loss/reduced viability and reduced attachment ability (plating efficiency) determines the outcome of our attachment assays. Despite the unchanged viability in glycine-/alanine-containing solutions, cell attachment after cold storage was severely decreased in the absence of iron chelators (Fig. 3, basic cold storage solution, and previous results). 14 Chloride-dependent cold-induced injury, which is seen in adherent rat hepatocytes during early rewarming 7 and also prevents hepatocyte attachment after cold storage in suspension, 14 did not play a role in the current setting, since we used chloride-poor variants for cold storage in all experiments. Cells nevertheless experienced functional impairment, which prevented cell attachment, and was, as cold-induced injury in attached hepatocytes, 4,6 iron-dependent (Fig. 3).
Mitochondrial Integrity after Cold Storage
In adherent hepatocytes, the iron-dependent cold-induced cell injury mainly targets the mitochondria. 6 TFP + fructose, a potent inhibitor combination that protects adherent hepatocytes against mitochondrial permeability transition and subsequent apoptosis, 6,15 also increased cell attachment rate (Fig. 3A), although this inhibitor combination was not quite as effective as the iron chelators. This result suggests that mitochondria, especially the mitochondrial permeability transition, also play a major role in the loss of attachment ability of hepatocytes in suspension.
Loss of MMP, which already occurred after cold storage in the absence of iron chelators, became more pronounced during early rewarming and could be partially inhibited by iron chelators (Figs. 4 and 5A). A loss or partial loss of MMP during cold storage was already described for adherent hepatocytes 6 and liver endothelial cells. 25 In most cells, complete loss of MMP is prevented by iron chelators and partial loss is reversible during rewarming when cells are protected against cold-induced injury. 6,16,25 The normal MMP in attached cells (Fig. 6) and the depletion of cells with full MMP in the supernatant during cell attachment (Fig. 5B) imply that those cells which maintained mitochondrial integrity are the ones which were able to attach.
Oxygen Consumption and Energy State
Rat hepatocyte suspensions displayed a dramatically low energy state after 1 wk of cold storage (Fig. 7A) despite oxygen availability. This is similar to—but much more pronounced than—the ATP loss described by Ostrowska et al. 11 after aerobic storage of human hepatocyte suspensions for 72 h. A decrease in ATP content at the end of cold storage has also been described for cold-stored rat livers, 26 isolated rat hepatocytes, 27 and rat hepatocyte spheroids, 28 but all under conditions of combined hypothermia and hypoxia. Although ATP steady state levels, even in the presence of oxygen, might also be influenced by hypothermia (by differing temperature sensitivity of ATP-producing and -consuming processes), this is unlikely a sufficient explanation for the very low ATP levels observed after cold incubation in the current study. Furthermore, the ability of cells in suspension to regenerate ATP during normoxic rewarming was impaired (Fig. 7B) and oxygen consumption was decreased (Fig. 8). This further excluded altered steady state and confirmed mitochondrial injury. Iron chelators not only provided a partial protection of mitochondria (Fig. 5) but also increased oxygen consumption (Fig. 8) and allowed partial regeneration of ATP after return to normothermic conditions (Fig. 7).
Role of Energy State in Cell Attachment
Our results suggest that the low energy status after cold storage and the impaired energy regeneration during the early rewarming phase lead to attachment failure. It is likely that the reconstitution of the hepatocellular cytoskeleton, known to be degraded during cold storage, 27,29,30 and cytoskeletal restructuring during attachment processes requiring ATP cannot take place in the energetically compromised cells, leading to attachment failure. This was confirmed by the accumulation of cells without MMP in the supernatant during the attachment phase (Fig. 5B). Further evidence for the role of the energy state in attachment was the nonattachment of energetically compromised nonstored cells after antimycin A treatment (Fig. 9).
Storage Options for Hepatocyte Suspensions
For longer storage periods exceeding several hours, hepatocytes may be cryopreserved. However, cryopreservation induces severe damage to hepatocytes in terms of reduced post-thaw viability, reduced attachment ability, and reduced metabolic activity 31 –34 along with cell loss during purification steps after thawing, which, in total, may add up to a loss of >50% of the overall number of functional hepatocytes. For cell transplantation, short periods of cold incubation at 0 to 6 °C already occur during the preparation process and short-term storage is required during assessment of cell function. If cell transplantation is performed in several stages, up to 48 h may pass between the first and last transplantation session. 23,35 Furthermore, periods of cold storage may occur during shipping or for delayed experimental use of isolated cells. For rat hepatocytes as used in the current study, standard cryopreservation (in cell culture medium with 10% foetal calf serum (FCS) and 10% dimethyl sulfoxide (DMSO) and a cooling rate of −1 °C/min in a controlled-rate freezer) yielded only about 10% of adherent viable cells (compared to control cultures; G. Pless-Petig, U. Rauen, unpublished results) in contrast to around 60% adherent viable cells after 1 wk of cold storage (Fig. 3A). Thus, for short- to medium-term preservation (up to several days), cold (4 °C) storage of hepatocyte suspensions under suitable protective conditions might be superior to cryopreservation.
Conclusion and Outlook
We here showed that mitochondrial integrity plays a major role for cell attachment after cold storage of rat hepatocyte suspensions. Protection of mitochondrial function during cold storage, as achieved by iron chelators, improved energy state and preserved cell attachment ability.
This important role of the mitochondria in nonattachment of rat hepatocytes stresses the importance of protection of mitochondrial integrity during cold storage of hepatocytes, a finding that might be of relevance for various fields, including storage of cell suspensions for toxicological assays or—provided the findings can be confirmed for human hepatocytes—storage/transport of hepatocytes for hepatocyte transplantation.
Footnotes
Acknowledgments
We thank Ms. J. Weiss and Mr. C. Fehring for their excellent technical support.
Author’s Note
U. Rauen obtained consulting fees from Dr. Franz Köhler Chemie GmbH, Bensheim, Germany. Dr. F. Köhler Chemie holds a patent on a preservation solution covering the modified cold storage solutions used in this study. However, the study design and data interpretation has never been influenced by the company.
Ethical Approval
The use of rats for the cell isolation for the current study was approved by the Landesamt für Natur, Umwelt und Verbraucherschutz NRW.
Statement of Human and Animal Rights
This article does not contain any studies with human subjects. Animals received humane care in compliance with the German Law for the Protection of Animals and following institutional guidelines. Permission for liver cell isolation was obtained from local authorities (Landesamt für Natur, Umwelt und Verbraucherschutz NRW, Az. 84-02.05.20.12.007 and Az. 84-02.04.2015.A485).
Statement of Informed Consent
There are no human subjects in this article and informed consent is not applicable.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.