
Abstract
Objectives:
To observe the expression of the dopamine transporter (DAT) in six cerebral regions of a methamphetamine (MA)-dependent rat, which were frontal cortex, nucleus accumbens septi, striatum, hippocampus, substantia nigra and ventral tegmental area.
Methods:
The rats were administrated intraperitoneally with 10 mg/kg/day of MA for 10 days consecutively; the behaviour changes were measured via the conditioned place preference (CPP), and the scores of stereotyped behaviour (SB) were used to confirm animal addiction. Then, the animals were further injected with MA respectively for 1, 2, 4 and 8 weeks to establish different periods of MA-dependent models. The expressions of DAT and DAT messenger RNA in six cerebral regions were detected.
Results:
The results of CPP and SB scores were significant different when comparing all four experimental groups with the control group (p < 0.05). Comparing between different experimental groups, the expression of DAT mainly decreased and had dynamic changes in the same regions (p < 0.05). Comparing the different regions with each other in the same experimental group, the expression of DAT also had significant difference in several regions p < 0.05).
Conclusions:
The expression of DAT mainly decreased and had different in the six cerebral regions at the same MA-dependent time period as well as at different time periods in the same cerebral region. It was speculated that DAT might play a crucial role in the mechanism of MA dependence.
Introduction
Methamphetamine (MA) is a monoamine, which is highly lipophilic, and exerts significant excitation on the central nervous system (CNS). It can induce neurochemical, pathological and behaviour changes in CNS during long-term addiction. MA can increase the release of dopamine and upregulates the function of dopaminergic system in the brain. 1 In the animals treated with MA, both dopamine and its metabolite are reduced in the cerebral regions including the striatum, hippocampus and frontal cortex, indicating that the toxicity of MA has close integration with the dopaminergic system in the brain. 2,3
Dopamine enlists the dopamine transporter (DAT) and specific dopamine receptor (DR) for its physiological functions. Literature indicates that DAT and DR have important roles in the mechanism of MA dependence. MA enters the neuron or nerve terminal via the effect of DAT and replaces the dopamine which is in the cells or its vesicles. The monoamine which is replaced then transforms to reactive oxygen species, leading to a series of chemical reaction and the cells will finally die. 4
Past results showed that antioxidants could reduce the injury caused by MA. 5 DAT is an important component of the synapse in the nerve terminal, which regulates the cell function related to dopamine. DAT has crucial effects in hyperthermia and in the toxicity induced by MA. DAT is itself upregulated via activation of the dopamine 2 receptor. 6 Past studies showed that both single and repeated in vivo amphetamine and/or MA administrations reduced DAT uptake, but did not altered DAT localization posttreatment. 7
MA dependence affected many cerebral regions of the CNS, such as the striatum 8,9 and hippocampus. 10 Recently, Yuan et al. 11 found that the Han Chinese people with MA dependence had significant DAT dysfunction in the striatum, and the DAT levels were reversible after 4 weeks of abstinence but still were remained lower than those in the controls. The recovery of DAT might parallel improvements in craving, anxiety and depression.
However, there were few papers that focused on many regions of the dopaminergic nervous system in the CNS and, for example, regions of mesolimbic system play important roles in the mechanism of MA dependence. Thus, this study selected six regions of mesolimbic system, which were the frontal cortex, nucleus accumbens septi, striatum, hippocampus, substantia nigra and ventral tegmental area to investigate the changes of DAT in these regions and the relationship between the expression of DAT and the mechanism of MA dependence.
Material and methods
Purification the abusive drug tablets
The tablets named ‘ice’ were provided by the Institute of Criminal Science and Technology of Public Security Bureau of Lincang City, Yunnan Province, People’s Republic of China. The mean purity of the MA tablets was 8.40% as detected by the gas chromatograph–mass spectrometer (GC-MS). Then, the tablets were purified in the laboratory of the School of Chemistry, Yunnan University. The chemical constitution after purification was identified using the method of nuclear magnetic resonance (NMR). The purity of final products was above 98%.
Establishment of animal models and behavioural tests
Forty healthy SD rats, 20 males and 20 females, weighing 260 ± 20 g were used in accordance with the ethics committee of the Kunming Medical University’s guidelines for the care and use of laboratory animals. The animals were divided into 5 groups (n = 8): group of MA dependence for 1 week, group of MA dependence for 2 weeks, group of MA dependence for 4 weeks, group of MA dependence for 8 weeks, and the control group. The rats were housed respectively in cages at 20°C, with 40% humidity and under 12-h light/12-hdark cycle with free access to water and rat chow.
The protocol of the biased procedure of conditioned place preference (CPP) was used along with the stereotyped behaviour (SB) to confirm the rat model of MA dependence. The apparatus of CPP has two big conditioned chambers and a small connected chamber. Two big chambers had different shapes and colours of floors and were located at the end of each side, with the small connected chamber as a corridor. The animals could go to the two conditioned chambers freely through the sliding door during the accommodation period before the drug treatment. The natural preference chamber for each rat and the time it stayed in before drug administration was recorded, and the rat would be put into the non-preference chamber after each drug injection. After becoming drug addicted, the rat would prefer to stay at the latter chamber rather than to stay in its original natural preference chamber. This phenomenon would confirm that the rat was addicted. 12,13
For this test, the animals were put into the CPP apparatus to accommodate for 15 min once per day for consecutive 3 days before the drug administration. On day 4, the animals of the dependent groups were administered with MA (intraperitoneally, 10 mg/kg, once at 8:00 a.m.), and the animals were put into the non-natural preference chamber for 30 min. Simultaneously, the scores of SB were recorded with reference to the methods of Sams-Dodd. 14 These procedures were maintained for 10 consecutive days. The control group was put on the same protocol, but injected with the equivalent dose of saline only. On day 11, the CPP tests were performed and the duration of each rat that stayed at the conditioned chambers was recorded. The results of CPP and the scores of SB of the experimental groups were compared with the control group to confirm that the MA-dependent rat model was established successfully.
After the animals were confirmed to be drug dependent, the four dependent groups were further injected with doses of 10 mg/kg MA for another consecutive 1, 2, 4 and 8 weeks respectively, to establish the model of different dependent periods. Then, the animals of different groups were examined with the CPP procedure on the first day after the drug injection had finished respectively. The control group had the same protocol but with injection of saline only.
Western blotting
The animals were anaesthetized by 10% chloral hydrate and killed, the whole brain was dissected out on ice and the six cerebral regions were located according to the stereotaxic atlas of rat brain. The samples were put into 1.5 ml Eppendorf pipes with 0.5 ml extraction buffer solution and 5 μl phenylmethyl sulphonylfluoride (PMSF, 10 mg/ml) and the content was homogenized for 2 min. Then, the samples were centrifuged at 12,000 r/min for 40 min at 4°C. The supernatant were transferred, and the protein concentration was determined by the DC protein assay kit (Bio-Rad Laboratories Inc., Berkeley, California, USA).
Then, the samples were subjected to 10% sodium dodecyl sulphate (SDS)–polyacrylamide gel electrophoresis and transferred to a nitrocellulose membrane at 4°C with 18 V for 6.5 h. The membrane was blocked with 10% skim dry milk in Tris-buffered saline for 8 h at 4°C and then incubated overnight at 4°C with primary antibodies such as β-actin (1:400, Boster Biotechnology, Wuhan, China) and DAT (1:400, Boster Biotechnology, Wuhan, China). The membrane was put on the swing bed at room temperature for 1 h and incubated at 37°C for 2 h after washing with phosphate-buffered saline with Tween 20 on the next day. Then, the membrane was further incubated with horseradish peroxidase (HRP)-conjugated secondary antibody (immunoglobulin G, 1:5000) for 1 h at room temperature. Blots were developed using an ECM Western Blotting Plus Kit (Boster Biotechnology, Wuhan, China) and exposed on a Kodak medical X-ray film for 30 s, subsequently developed for 1 min and fixed. The optical density (OD) values were quantified with QuantityOne software (Bio-Rad Laboratories Inc.).
The procedure of RT-PCR
For reverse transcription polymerase chain reaction (PCR) analysis, total RNA was isolated from brain tissues using TRIZOL reagent (Molecular Research Center Inc., Cincinnati, Ohio, USA). The yield of the total RNA was measured at 260 nm and 280 nm using a spectrophotometer ND-1000 (Bio-Rad Laboratories Inc.). Then, Revert Aid™ First-strand complementary DNA (cDNA) synthesis kit (Fermentas, Thermo Fisher Scientific Inc., Waltham, Massachusetts, USA) was used to reverse transcript the first chain of cDNA according the manufacturer’s protocol. Then the PCR was employed to amplify the genes of DAT and β-actin. The primers of DAT and endogenous β-actin were designed using Primer Premier 5.0 software (Premier Biosoft International, Palo Alto, California, USA). The primer of rat DAT was as follows: forward primer: 5′-CAGCCTATGGAAGGGAGTAAA-3′, reverse primer: 5′-CAGGAAAGTAGCCAGGACAAT-3′. The primer of rat β-actin was as follows: forward primer: 5′-GTAAAGACCTCTATGCCAACA-3′, reverse primer: 5′-GGACTCATCGTACTCCTGCT-3′.
The amplification reactions were performed using a 7900 HT Fast real-time PCR system (Applied Biosystems, Foster City, California, USA). The PCR parameters were pre-denatured for 5 min at 94°C, and 35 cycles were programmed as follows: denaturation at 94°C for 1 min, annealing 55°C for 1 min and extension 72°C for 1 min. Total reaction volume was 25 µl which included 12.5 µl 2× PCR Master Mix from PCR Master Mix Kit (Fermentas, Thermo Fisher Scientific Inc.), 1 µl cDNA template, 0.5 µl forward primer, 0.5 µl reverse primer and nuclease-free water 10.5 µl.
Statistical analysis
The results of CPP were compared using the t-test. The data of Western blotting and RT-PCR were analyzed by one-way analysis of variance following Fisher’s protected least optical significant difference test (SPSS software version 11.5). Criteria for significance were set at the 0.05 level.
Results
The Scores of SB
The methods of Sams-Dodd 14 were used to grade the scores of SB. MA-dependent rats were found to exhibit obvious SB that had significant differences when compared with the rats in the control group (see Figure 1).

The groups of MA-dependent rats were found to exhibit obvious SBs and had significant difference when comparing with the control group, respectively (p < 0.05). MA: methamphetamine; SB: stereotyped behaviour.
Besides the SB which was described by Sams-Dodd, 14 we also observed other aberrant behaviours in the MA-dependent animals, including standing tail, accentuation of muscular tension, prostrated and expansion of the limbs, general irritability and occasional shaking of heads. Some rats were found eating their stool, chewing on the sawdust in the cage, biting its own limbs, attacking, and manifesting hydrostomia. In addition, signs of SB induced by MA were observed for 4–8 h after injection, which included irritability, hyperactivity, accentuation of muscular tension and standing tail. Especially in the first stage of drug injection, the signs of accentuation of muscular tension and irritability could be observed even 12 h after drug administration.
The result of CPP
The results of CPP revealed rats injected with MA preferred to stay at the chamber which was of non-natural preference after drug administration, and the duration was longer than staying in the chamber which was of natural preference at the stage of accommodation. There were significant differences when comparing the results of CPP of the four dependent groups with the control group, which implied that the animals produced CPP as a consequence of the drug administration and thus the animals were confirmed to be drug addicted (see Figure 2).

The results of CPP revealed that the rats of MA-dependent groups preferred to stay in the chamber which was of non-natural preference but related to drug administration, and the durations were longer than their stay in the other chamber. There was significant difference when comparing the control group with the MA-dependent 1 week group, MA-dependent 2 weeks group, MA-dependent 4 weeks group, and MA-dependent 8 weeks group, respectively (*p < 0.05). MA: methamphetamine; CPP: conditioned place preference.
The results of Western blotting
The expression of DAT protein at the same cerebral region in the different experimental groups was compared. First of all, the expression of DAT decreased in all four cerebral regions in the present research, which included the frontal cortex, the nucleus accumbens septi, the striatum and the substantia nigra. On the contrary, the expression of DAT increased in some other cerebral regions, for example, the hippocampus and the ventral tegmental area. Second, there were significant differences in cerebral regions of the frontal cortex, the nucleus accumbens septi, the striatum and the ventral tegmental area, when comparing the same cerebral region of the control group with the four MA-dependent groups. There were no significant differences in other regions of the hippocampus and the substantia nigra. The data were shown in Figure 3(a) to (f).
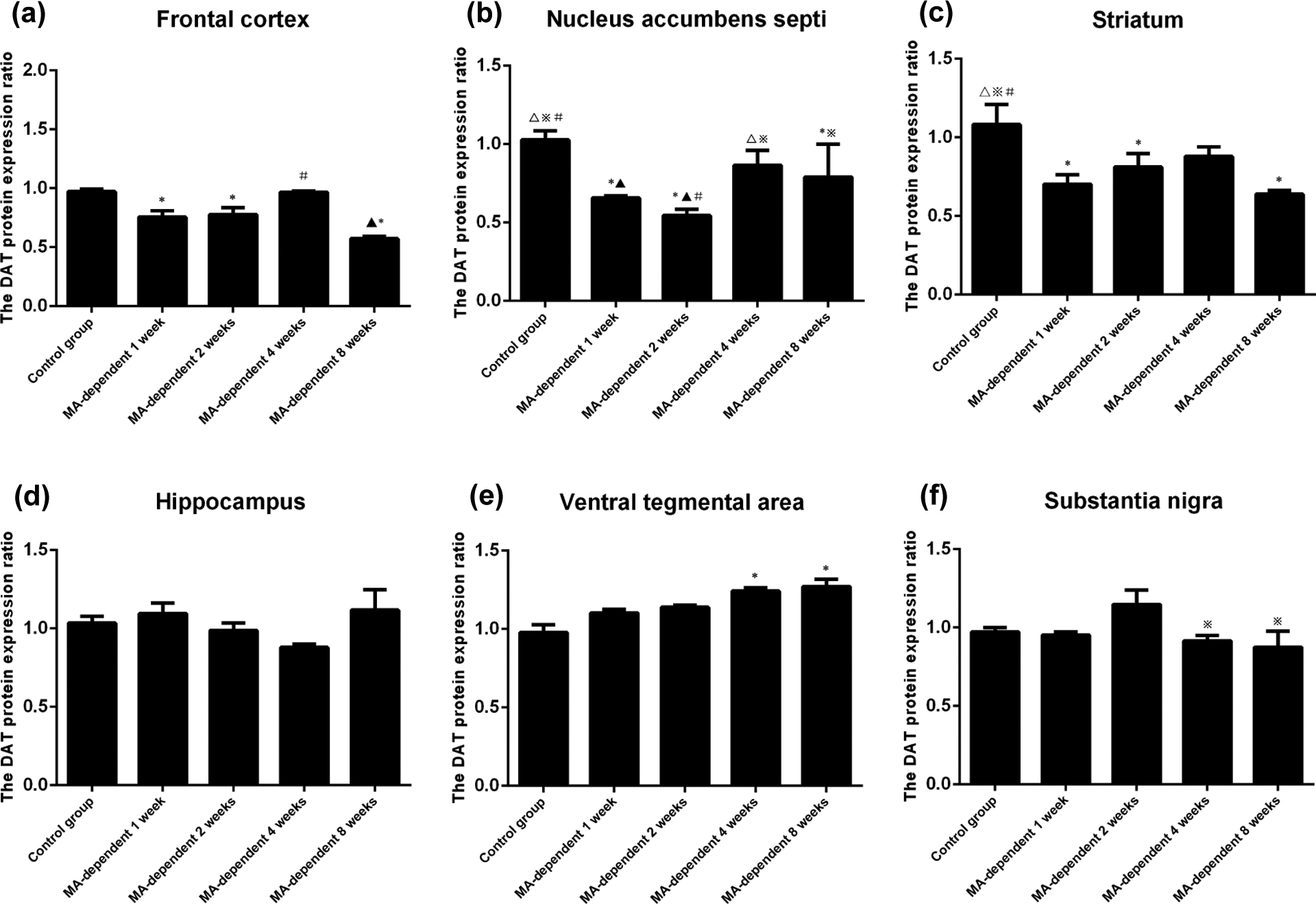
The results of DAT protein expression in the same cerebral region in different experimental groups of MA dependence. (a) The DAT protein expressed in the frontal cortex, and there was significant difference when one compared the control group (0.972 ± 0.022) with 1 week (0.756 ± 0.054), 2 weeks (0.778 ± 0.967), 8 weeks (0.572 ± 0.020) and when comparing the 4 weeks (0.967 ± 0.011) with the 8 weeks groups (p < 0.05). (b) Data in the nucleus accumbens septi show significant differences when comparing the control group (1.027 ± 0.057) with 1 week (0.657 ± 0.012), 2 weeks (0.544 ± 0.041), and 8 weeks (0.791 ± 0.208), respectively. There was significant difference when comparing the 1 week (0.657 ± 0.012) group with the 4 weeks (0.865 ± 0.096) group, and the 2 weeks group with the 4 weeks and the 8 weeks groups, respectively (p < 0.05). (c) The data in the striatum show significant difference when comparing the control group (1.082 ± 0.127) with 1 week (0.703 ± 0.060), 2 weeks (0.812 ± 0.086), and 8 weeks (0.639 ± 0.024) groups, respectively (p < 0.05). (d) The data in the hippocampus show no significant difference when comparing the control group with all 4 weeks MA-dependent groups, respectively (p > 0.05). (e) The data in the ventral tegmental area is shown, and there was significant difference when comparing the control group (0.978 ± 0.050) with 4 weeks (1.242 ± 0.021) and 8 weeks groups (1.271 ± 0.046), respectively (p < 0.05). (f) The data in the substantia nigra show no significant difference when comparing the control group with all four MA-dependent groups, but there were significant differences when comparing the 2 weeks group (1.148 ± 0.092) with the 4 weeks (0.915 ± 0.036) and 8 weeks groups (0.876 ± 0.101), respectively (p < 0.05). MA: methamphetamine; DAT: dopamine transporter.
In the frontal cortex, the DAT protein decreased in the 1-week MA-dependent group, slightly increased in the 2 and 4 weeks groups and decreased again in the 8 weeks group (Figure 3(a)). In the nucleus accumbens septi, the DAT protein decreased in 1 week and more obviously in the 2 weeks groups, and then increased in the 4 weeks group and slightly decreased again in the 8 weeks group (Figure 3(b)). In the striatum, the DAT protein decreased in 1 week and slightly increased in 2 weeks group and the increase persisted in 4 weeks group, then slightly decreased again in 8 weeks group (Figure 3(c)). In the hippocampus, the DAT protein increased in 1 week group, decreased in 2 and 4 weeks groups, then increased again in 8 weeks group (Figure 3(d)). In the ventral tegmental area, the DAT protein increased from 1 week to 8 weeks and reached a high level in the 8 weeks group (Figure 3(e)). In the substantia nigra, the DAT protein decreased in 1 week group and then increased in 2 weeks group. By 4 weeks, it decreased again and such decrease persisted in the 8 weeks group (Figure 3(f)).
Also when the expression of DAT protein at six different cerebral regions of the same experimental group was compared, the significant differences were shown between cerebral regions in the same experimental group. The details were depicted in Figure 4(a) to (e).

The results of DAT protein expression in the different regions in the same experiment group of MA dependence. (a) In the control group, there was significant difference when comparing the ventral tegmental area (*) with the frontal cortex, nucleus accumbens septi, striatum and substantia nigra, respectively (p < 0.05). There were significant differences when comparing hippocampus (※) with striatum and substantia nigra, respectively (p < 0.05), when comparing striatum (Δ) with hippocampus and ventral tegmental area, respectively (p < 0.05) and when comparing substantia nigra (#) with hippocampus and ventral tegmental area, respectively (p < 0.05). (b) In the MA-dependent 1 week group, there was significant difference when comparing the substantia nigra (#) with the frontal cortex, nucleus accumbens septi, striatum, hippocampus and ventral tegmental area, respectively (p < 0.05). (c) In the MA-dependent 2 weeks group, there was significant difference when comparing the ventral tegmental area (*) with striatum (p < 0.05). There were significant differences when comparing hippocampus (※) with striatum and substantia nigra, respectively (p < 0.05) and when comparing striatum (Δ) with nucleus accumbens septi, hippocampus, ventral tegmental area and substantia nigra, respectively (p < 0.05) and also when comparing substantia nigra (#) with frontal cortex, nucleus accumbens septi, striatum, hippocampus and ventral tegmental area, respectively (p < 0.05). (d) In MA-dependent 4 weeks group, there was significant difference when comparing the ventral tegmental area (*) with substantia nigra (p < 0.05). There were significant differences when comparing hippocampus (※) with frontal cortex, nucleus accumbens septi, striatum and substantia nigra, respectively (p < 0.05). (e) In the MA-dependent 8 weeks group, there was significant difference when comparing the ventral tegmental area (*) with frontal cortex and substantia nigra (p < 0.05). There was significant difference when comparing the substantia nigra (#) with frontal cortex, nucleus accumbens septi, striatum, hippocampus and ventral tegmental area, respectively (p < 0.05). MA: methamphetamine; DAT: dopamine transporter.
The results of RT-PCR
Firstly, the expression of DAT mRNA of the same cerebral region in the different experimental groups and control group were compared. DAT mRNA decreased in four cerebral regions including the frontal cortex, the nucleus accumbens septi, the striatum and the substantia nigra. But DAT mRNA increased in the hippocampus and the ventral tegmental area. In addition, the data indicated that there were significant differences in the four regions involving the frontal cortex, the nucleus accumbens septi, the striatum and the ventral tegmental area. Whereas there were no significant differences in the hippocampus and the substantia nigra. The details are shown in Figure 5(a) to (f).

The results of DAT mRNA in the same cerebral region in the different experimental groups. (a) The DAT mRNA expressed in the frontal cortex, and there were significant differences when comparing control group (*) with 1 week group, 2 weeks group, and 8 weeks group, and when comparing the 4 weeks (▲) with 8 weeks group (#), respectively (p < 0.05). (b) In the nucleus accumbens septi, there were significant differences when comparing the control group (*) with 1 week group, 2 weeks group, and 8 weeks group as well as when comparing the 1 week group (Δ) with 4 weeks group (▲); 2 weeks group (※) with 4 weeks and 8 weeks group (#), respectively (p < 0.05). (c) Data in the striatum show significant differences when comparing control group (*) with 1 week group (Δ), 2 weeks group (※) and 8 weeks group (#), respectively (p < 0.05). (d) Data in the hippocampus show no significant difference when comparing control group with all four experimental groups (p > 0.05). (e) Data in the ventral tegmental area show significant difference when comparing control group (*) with 4 weeks and the 8 weeks groups, respectively (p < 0.05). (f) Data in the substantia nigra show no significant difference when comparing control group with all four MA-dependent groups, but there were significant differences when comparing the 2 weeks group (※) with 4 weeks group, and 8 weeks group, respectively (p < 0.05). MA: methamphetamine; DAT: dopamine transporter; mRNA: messenger RNA.
In the frontal cortex, the DAT mRNA in the 1 week MA-dependent group decreased (30.75%), but increased a bit by 2 weeks and further increased nearly to the normal level (91.68%) by 4 weeks. It significantly decreased again by 8 weeks to 45.33% of what was the normal value (Figure 5(a)). In the nucleus accumbens septi, the DAT mRNA in the 1 week MA-dependent and 2 weeks MA-dependent groups decreased (36.18% and 46.36%). Interestingly, by 4 weeks, DAT mRNA increased obviously and then slightly decreased by 8 weeks and the ratio was 22.89% of the normal value (Figure 5(b)). In the striatum, the DAT mRNA in the 1 week MA-dependent group decreased (35.1%), followed by minor increase by 2 weeks and increased obviously by 4 weeks, then slightly decreased again by 8 weeks to 41.81% of the normal value (Figure 5(c)). In hippocampus, the DAT mRNA in the 1 week MA-dependent group increased about 5.66% to that of normal and then slightly decreased from 2 weeks to 4 weeks to 15.14% of normal, followed by slight increase to higher than control group by the 8 weeks (108.98% of normal; Figure 5(d)). In the ventral tegmental area, the DAT mRNA increased from 1 week to 8 weeks, as much as over 29.58% of normal (Figure 5(e)). In substantia nigra, the DAT mRNA in 1 week MA-dependent group decreased slightly (by 1.89% of normal) and increased (over 18.04% of normal) by 2 weeks, then it decreased again by 4 weeks (by 6.7% of normal) and 8 weeks group (by 9.54% of normal; Figure 5(f)).
The DAT mRNA of different cerebral regions in the same experimental group was elaborated. The results indicated that there were significant differences in several regions when comparing these regions with each other in the same experimental group. The details are depicted in Figure 6(a) to (e).

The results of DAT mRNA in different regions in the same experiment group of MA dependence. (a) The data in the control group show significant differences when comparing the ventral tegmental area (*) with frontal cortex, nucleus accumbens septi, striatum and substantia nigra and also when comparing hippocampus (※) with striatum (Δ), and substantia nigra (#), respectively (p < 0.05). (b) The data in the MA-dependent 1 week group show significant differences when comparing the substantia nigra (#) with the frontal cortex, ventral tegmental area, striatum, hippocampus and nucleus accumbens septi, respectively (p < 0.05). (c) The data in the MA-dependent 2 weeks group show significant differences when comparing the striatum (Δ) with nucleus accumbens septi, hippocampus, ventral tegmental area; and comparing the substantia nigra (#) with the frontal cortex, nucleus accumbens septi, striatum, hippocampus and ventral tegmental area, respectively (p < 0.05). (d) The data in the MA-dependent 4 weeks group show significant differences when comparing the hippocampus (※) with the frontl cortex, nucleus accumbens septi, striatum and substantia nigra; and comparing the substantia nigra with ventral tegmental area (*), respectively (p < 0.05). (e) The data in the MA-dependent 8 weeks group show significant differences when comparing the substantia nigra (#) and frontal cortex, nucleus accumbens septi, striatum, hippocampus and ventral tegmental area; and comparing frontal cortex with ventral tegmental area (*), respectively (p < 0.05). MA: methamphetamine; DAT: dopamine transporter.
Discussion
MA and related drugs can upgrade the intercellular dopamine density as the substrate of DAT and via the transport of DAT. MA is a highly addictive mental stimulant. High dose and repeated administration of MA can cause long-term exhaustion of dopamine and degeneration of dopaminergic nerve terminals. Post-mortem functional imaging of the MA users indicated that the DAT density of chronic MA abuser had decreased, and there was reduction of the DAT-binding ligands in several cerebral regions including the caudate nucleus and putamen, 15 striatum, 16 the orbitofrontal cortex, dorsolateral prefrontal cortex and amygdala. 17
The MA can enter the presynaptic vesicle and competitively replaces intra-vesicle monoamine, which then leads to postsynaptic monoamine over release. 18 MA has significant effects on many neurochemicals, including dopamine, 19 γ-aminobutyric acid 20 and glutamic acid. 21 Acute or long-term administration of MA also can cause the postsynaptic dopamine increase in the nigro striatum. 22
Whilst there were other results which indicated that DAT might have no major contribution to the mechanism of MA dependence, Kokoshka et al. 23 showed that saline and MA could influence the co-migration of DAT in the SDS gel at approximately 80 kDa and that acute MA induced decreases of [ 3 H]-dopamine uptake were not due to loss of DAT proteins. This result was the first report on MA having no specific effect on the acute changes of DAT function. Also in the same year, Fumagalli et al. 24 found that DAT had an important role in the dopamine release and could be induced by MA, and the DAT gene deleted mouse was resistant to the neurotoxicity of MA when compared with the normal mouse. Fleckenstein et al. 25 determined the effect of a single high dose of MA on DAT activity in striatal synaptosomes and found that high-dose MA promoted the production of free radicals, and the DAT of striatum was rapidly inactivated by the oxides, so that the dopamine uptake was reduced by 48% 1 h after MA administration. This dramatic decline was reversed 24 h after drug administration. These results indicated that the reversibly decrease of DAT caused by MA had close correlation with oxidization. Recent research also confirmed that MA exposure at the high dose (10 mg/kg) reduced DAT levels in both μ-opioid receptor knockout mice and wild-type mice. 26 These results also demonstrated that the DAT to some extent had contribution to the neurotoxicity of MA.
Our data showed that DAT protein was expressed at the six specific regions of mesolimbic system, including the frontal cortex, nucleus accumbens septi, striatum, hippocampus, ventral tegmental area and substantia nigra. The DAT expression was not only different in time course after MA treatment but was also different in specific regions. The DAT expression had a similar tendency in the frontal cortex, nucleus accumbens septi and striatum. DAT decreased 1 week after MA administration, increased by 2–4 weeks, and then decreased again at 8 weeks of MA administration. In the hippocampus, DAT increased after 1 week of MA administration, decreased from 2 weeks to 4 weeks and then increased by 8 weeks of MA administration. In the ventral tegmental area, DAT continuously ascended and was most significant by 8 weeks of MA administration. In substantia nigra, DAT decreased after 1 week of treatment, increased by 2 weeks, and then decreased again from 4 weeks to 8 weeks of MA administration.
Dopaminergic fibres in mesolimbic cortex system originated from the dopaminergic neurons in the ventral tegmental area. The dopaminergic nerve fibres travelled upward along medial forebrain bundle, deposited in the septal area and other dopaminergic nerve fibres went forward to the anterior olfactory nucleus and bulbus cinereus. Mesolimbic system reward pathway originated from the ventral tegmental area and the medial part of substantia nigra compacta, then projected nerve fibres to nucleus accumbens septi, amygdaloid nucleus, caruncula mammillaris and frontal cortex. In the six regions studied in the present research, the ventral tegmental area, which was the original part of mesolimbic system, had continuous increased expression of DAT. But DAT decreased firstly in the substantia nigra, then increased at 2 weeks and decreased again from 4 weeks to 8 weeks of MA administration. At the other cerebral regions including nucleus accumbens septi, frontal cortex, which were the regions where dopaminergic fibres projected to, the expression of DAT decreased firstly at 1 week, then increased from 2 weeks to 4 weeks and decreased again by 8 weeks of drug administration. These data indicated that in different cerebral regions of the rewards pathway, the changes of DAT were not of uniformity, and the phenomenon was the same in the regions of the ventral tegmental area, nucleus accumbens septi, hippocampus and frontal cortex. We need more evidence to explain the mechanism of these changes.
Another interesting result was in the sensitization of animals to MA, the DAT had different modes of regulation in different cerebral regions, and the MA-induced changes of dopamine transportation was not only totally due to the density of DAT but was also due to the changes of kinetics of DAT. 27 Taken together from the findings in the literature and in this study, there were changed expression of DAT in all six cerebral regions of mesolimbic cortex system, related to the time course of MA administration. The results of CPP were in accordance with the changes of DAT in different groups. These changes could be explained by the mechanism of the neuron’s sensitization and adaption to the MA administration. After drug administration, MA-induced DAT expression decreased. When the duration was prolonged, the neurons became adaptable to the MA and thus the expression of DAT rose again and the behaviour of animals became atypical. But the damage of neuron induced by MA was persistent, so the behavioural changes persisted and could be observed by the scores of SB. The results indicated that the expression of DAT in the six cerebral regions was different during adaptation to MA dependence. Considering the literature and our research data, the first possible reason was MA may cause differential damage in different cerebral regions. Secondly, the density and the dynamics state of DAT may be different in the six different regions. Thirdly, there were interactions between dopaminergic neurons and other neuronal system, such as serotoninergic neurons, which may further regulate the state of DAT.
Footnotes
Authors’ Note
L-H Li and DTW Yew are co-corresponding authors.
Conflict of interest
The authors declared no conflicts of interest.
Funding
This work was supported by the National Natural Science Foundation of China (grant number 81060253), the Collaboration Foundation of Department of Science and Technology of Yunnan Province and Kunming Medical University (grant number 2008CD051) and the Postgraduate Scientific Foundation of Kunming Medical University (grant number KM2007D05).