
Abstract
Circadian rhythms are generated by interlocked transcriptional-translational feedback loops of circadian clock genes and their protein products. Mice homozygous for a functional deletion in the Period-2 gene (Per2m/m mice) exhibit short free-running circadian periods and eventually lose behavioral circadian rhythmicity in constant darkness (DD). We investigated Per2m/m mice in DD for several months and identified a categorical sex difference in the dependence on Per2 for maintenance of circadian rhythms. Nearly all female Per2m/m mice became circadian arrhythmic in DD, whereas free-running rhythms persisted in 37% of males. Remarkably, with extended testing, Per2m/m mice did not remain arrhythmic in DD, but after varying intervals spontaneously recovered robust, free-running circadian rhythms, with periods shorter than those expressed prior to arrhythmia. Spontaneous recovery was strikingly sex-biased, occurring in 95% of females and 33% of males. Castration in adulthood resulted in male Per2m/m mice exhibiting female-like levels of arrhythmia in DD, but did not affect spontaneous recovery. The circadian pacemaker of many gonad-intact males, but not females, can persist in DD for long intervals without a functional PER2 protein; their circadian clocks may be in an unstable equilibrium, incapable of sustaining persistent coherent circadian organization, resulting in transient cycles of circadian organization and arrhythmia.
Circadian rhythms in vertebrate physiology and behavior are generated by a molecular cascade in which transcription of the “clock genes” Period (Per) 1 and 2 and Cryptochrome (Cry) 1 and 2 are driven by the CLOCK:BMAL1 complex. PER and CRY heterodimerize, translocate to the nucleus, and repress the transcription of Clock and Bmal1, thus inhibiting their own production. Accessory circadian feedback loops, including Ror and Rev-erb genes, add stability and redundancy to the circadian genomic network (Mohawk et al., 2012). Together, these clock gene interactions create an autoregulatory transcriptional-translational feedback loop with a circadian period (tau; τ) of ~24 h (Panda et al., 2002; Storch et al., 2002; Ko and Takahashi, 2006; Mohawk et al., 2012).
PER2 is critical for the integrity of the organismal circadian network: mutant mice (mPer2Brdm1) with a functional deletion of the mPER2 protein dimerization PAS domain initially exhibit an extremely short free-running circadian period in constant darkness (DD) and become behaviorally circadian arrhythmic (Zheng et al., 1999); mice with a null mutation in the mPER2 gene (mPer2ldc) exhibit a similar DD arrhythmia phenotype (Bae et al., 2001). In both mPer2Brdm1 and mPer2ldc mutants, the persistence of circadian rhythmicity in constant conditions varies from a few days to several weeks. In humans, hPer2 single nucleotide polymorphisms are associated with changes in circadian period (Chang et al., 2019) and an early chronotype (Jones et al., 2016), and phosphorylation of hPER2 is linked to a short τ and heritable circadian sleep disorders (Toh et al., 2001). Studies of Per2m/m animals have assessed the impact of this gene on reproductive and maternal behavior (Pilorz and Steinlechner, 2008) and documented how gonadal hormones affect Per2 expression (Nakamura et al., 2005; Nakamura et al., 2008). Of particular note, ovarian hormone secretion was disrupted in 9- to 12-month-old Per2m/m mice: 4-day estrous cycles occurred significantly less often than in wild-type (WT) mice (Pilorz and Steinlechner, 2008) and estradiol applied to explanted cultures from ovariectomized PER2::LUC knockin mice shortened the period of rhythmic PER2::LUC expression in uterus but not SCN (Nakamura et al., 2008). In general, the importance of circadian rhythms in modulating hormones of the reproductive system has been reviewed by Kriegsfeld (2013) and Bailey and Silver (2014). However, despite these studies, widespread evidence of sex differences in circadian biology (Yan and Silver, 2016), and a strong female bias in the prevalence of sleep disorders (Krishnan and Collop, 2006), the role of PER2 in the generation of circadian rhythms has not been specifically examined in female mice. We thus examined the role of gonadal sex on the circadian phenotype of mPer2-mutant mice in constant conditions. The outcomes revealed an unexpected and striking sex difference in the dependence on PER2 for the maintenance of circadian rhythmicity.
Materials and Methods
Experiments
Experiment 1 consisted of a pilot study to characterize sex differences in the necessity of mPER2 for the maintenance of circadian rhythmicity. Experiment 2a replicated experiment 1 and included multiple assays of locomotor activity (passive infrared and running wheels [RWs]). Experiment 2b examined the long-term persistence of circadian arrhythmicity in constant darkness. Experiment 3 examined the contribution of gonadal hormones toward the circadian behavioral phenotype in mPer2 mutant mice.
Animals
Adult female and male Per2 mutant mice (Per2m/m: B6.Cg-Per2tm1Brd Tyrc-Brd/J, JAX#: 003819) and control mice (WT: C57BL/6J [JAX#: 000664] or B6(Cg)-Tyrc-2J/J [JAX#: 000058], see below) were obtained from Jackson Laboratory (Bar Harbor, ME, USA). Mice of both sexes were single housed in conventional cages with wirebar lids, and without microisolator filters in a 12L:12D photocycle of approximately 150-200 lux until the housing room was switched to continuous darkness (DD). Each experiment was performed within a single room. Cage location in the room was chosen randomly. At all times mice had ad libitum access to standard rodent diet (Irradiated Teklad Global 18% Rodent Diet 2918, Envigo RMS) and filtered drinking water. Animal husbandry in DD was facilitated via dim handheld red illumination (<1 lux), otherwise DD mice were exposed to complete darkness. Cage changing was performed at 2-week intervals. All mice were acclimated to RW or passive infrared motion detector (PIR) cages for at least 1 week prior to data collection. The integrity of experimental LD cycles and DD treatments was continuously monitored and verified by dataloggers (HOBO, UX90, Onset Comp Bourne, MA, USA). Estrous cycles of females were not monitored. All experimental procedures complied with the ARRIVE guidelines. All procedures related to animal use were approved by the University of Chicago Institutional Animal Care and Use Committee.
For experiment 1, mice were ordered directly from JAX and upon arrival were placed under PIR sensors and exposed to 12L:12D (LD) for 21 days before subsequent transfer to DD for behavioral data collection (Per2m/m: B6.Cg-Per2tm1Brd Tyrc-Brd/J, JAX#: 003819 [albino]; WT: C57BL/6J, JAX#: 000664 [black]); mice in this experiment (Per2m/m: n = 5 females, n = 5 males; WT: n = 7 females, n = 7 males) were 2 months of age when transferred to DD. One male Per2m/m mouse died 2 weeks into experiment 2 and was excluded from all analyses. Mice in experiment 2 were the offspring of homozygous Per2m/m breeding pairs (Per2m/m: B6.Cg-Per2tm1Brd Tyrc-Brd/J, JAX#: 003819 [albino]) or WT breeding pairs (B6(Cg)-Tyrc-2J/J, JAX#: 000058 [albino]); mice in this experiment (Per2m/m: n = 20 females, n = 19 males; WT: n = 20 females, n = 17 males) were 2-5 months of age when transferred to DD. Mice in Experiment 3 were the offspring of heterozygous breeding pairs, heterozygous-homozygous breeding pairs, and homozygous breeding pairs, all of which were derived within 3 generations from heterozygous mice (B6.Cg-Per2tm1Brd Tyrc-Brd/J, JAX#: 003819; heterozygous for the Per2m/m and the TyrC-Brd mutations) supplied directly from JAX out of cryopreserved stock (8 of 39 Per2m/m mice, and 3 of 14 WT mice in experiment 3 exhibited an albino phenotype, the remainder were black). Mice in experiment 3 (Per2m/m: n = 20 females, n = 19 males; WT: n = 9 females, n = 5 males) were 8-12 months of age when transferred to DD and 6-10 months of age when gonadectomized. Offspring were group housed by sex at weaning and singly housed in adulthood shortly before experiments began. Initial sample sizes were designed to be >10 per group to permit sufficiently powered nonparametric tests on the incidence circadian arrhythmia.
At the conclusion of the experiments, homozygous WT and Per2m/m genotypes were confirmed in all mice by polymerase chain reaction (PCR) using the protocol described for this genotype by JAX (see below). Mice in experiment 1, in which experimental animals were delivered directly from JAX, were not genotyped. Heterozygous mice were not included in any analyses.
Activity Monitoring and Telemetry
Mice were housed in polypropylene cages equipped with either 7″ steel RWs or PIR, as described in detail in other reports (Kampf-Lassin et al., 2011; Prendergast et al., 2012). Activity was collected using Clocklab Acquisition software (Actimetrics; Evanston, IL, USA).
Circadian Activity Measures
Activity (PIR and RW) data were visualized and analyzed using Clocklab (Actimetrics). Actograms and fast Fourier transform (FFT) vector graphics were generated in Clocklab v2.5.3; all quantitative analyses were performed using Clocklab 6. Characterization and quantitative evaluation of circadian chronotypes in mice were performed via methodology described previously for this mutant (Zheng et al., 1999; Albrecht et al., 2001): double-plotted activity records were scored by one experimenter (I.Z.), blind to sex, genotype, and surgical manipulations. Circadian arrhythmia for the purposes of this analysis manifested as activity occurring randomly around the clock; distinct intervals of activity and rest were no longer evident, and the effect persisted for multiple days. The beginning and end of intervals of free-running activity and arrhythmicity were similarly determined and were verified via quantification of the amplitude of the circadian peak in the FFT performed on a 10-day epoch of activity either preceding or following the chronotype state change.
Additional quantification of circadian phenotypes included determination of phase angles of entrainment in LD, free-running circadian period in DD, and total activity counts and were performed using Clocklab 6 software. Daily activity onsets in LD were obtained directly from Clocklab’s “onset/offset” feature, with manual corrections performed by an experimenter blind to file identity. Total daily activity counts in LD were derived from Clocklab over a 10-day interval and aggregated in Matlab 2018b. Circadian period was calculated using a Lomb-Scargle periodogram (LSP) analyses on 10 days of DD activity data (Ruf, 1999; Tackenberg and Hughey, 2021). An FFT power spectrum with a Blackman-Harris window was used to determine rhythmic power in Clocklab 6, also using 10 days of data. Among WT and Per2m/m mice that did not exhibit circadian arrhythmia, LSP and FFT were computed on data from experimental dates yoked to mean (±1 day) onsets of arrhythmia (ARR) and recovery (FR2) in arrhythmic mice to control for age and duration of exposure to DD. In experiments 2 and 3, initial chronotypes (FR1) were evaluated beginning 10 days after transfer to DD. In a small number of cases, equipment malfunction caused missing data for the proscribed 10-day chronotype evaluation window, and in these instances the chronotype analysis window was delayed by the minimum number of days required to obtain 10 successive days of data. In one instance, a male’s recovery from circadian arrhythmia occurred during an interval of channel malfunction; in this case, the date of circadian recovery was assigned to the day on which the data collection resumed.
Wavelet Analyses
Wavelet scalograms were generated according to methods previously described (Leise and Harrington, 2011; Leise, 2013) using JLAB software with MATLAB 2018b. Briefly, wavelet matrices were calculated using activity data sampled at 1 min, with outliers (values greater than 4 x the standard deviation of each time series) replaced with an interpolated moving mean and NaN replaced with zeroes. A generalized Morse wavelet of the strictly analytic Airy family (γ = 3; to avoid negative frequency leakage) was utilized; time-frequency resolution was toggled to a β value of 10. To manage edge effects periodic boundary extension was performed by trimming of 1.5 × the longest period measured (as previously recommended). For aid of comparison scalograms were normalized across all individuals in all experiments by dividing each by its average total power. Once normalized the average value of every 3 time points in the wavelet matrix was found. This did not result in any visually detectable difference, but greatly reduced the graphical memory required to generate the scalograms.
Surgical Procedures
Mice were randomly assigned to either a gonadectomy or sham-operation control group in experiment 3. Gonadectomy was performed under 3%-4% isoflurane/O2 gas anesthesia. In males, a ventral midline incision was made, testicular blood vessels were ligated and cauterized, and the testes removed. In females, after a dorsal midline incision, ovarian blood vessels were ligated and cauterized, and the ovaries were removed. Incisions were closed with non-reabsorbable vinyl sutures and cutaneous wound clips. Topical antibiotic ointment was applied to the wound site. Analgesic buprenorphine was administered immediately after surgery and every 12 h for the next 48 h. Animals were allowed 2 months to recover prior to behavioral experiments. Surgical condition was confirmed in all mice at the conclusion of the study via necropsy.
Genotyping
Only homozygous WT or homozygous Per2m/m mice were included in these analyses (a priori criterion). Genotyping of all mice bred in our vivarium was done using primers from the Jackson Lab Website [5′ → 3′: Common Forward (TTC CAC TCT GTG GGT TTT GG), Wild-Type Reverse (AAA GGG CCT CTG TGT GAT TG), and Mutant Reverse (GCC AGA GGC CAC TTG TGT AG)] and using specification for the Platinum Taq Polymerase (Life Technologies, Invitrogen catalog number: 10966-018). For each individual PCR, 16.65 µL of DNAase-free H2O, 2.5 µL of 10× PCR buffer with no MgCl2, 0.75 µL 50 mN MgCl2, 0.5 µL 10 mM dNTP mix, 0.5 µL of each primer, and 0.1 µL of Taq were added to a master mix and thoroughly mixed by pipetting up and down. 22 µL of master mix was aliquoted and added to 3 µL of DNA derived via QIAGEN DNeasy Blood and Tissue Kits (catalog number: 69504 and 69506) in experiment 2 or from HotShot (Truett et al., 2000) in experiment 3, from tail clips collected at the conclusion of the studies. We used the following PCR protocol on a thermocycler: (1) 94 °C for 2 min, (2) 94 °C for 20 s, (3) 65 °C for 15 s, with a −0.5 °C decrease with each cycle, (4) 68 °C for 10 s, (5) repeat steps (2-4) 10 times, (6) 94 °C for 15 s, (7) 60 °C for 15 s, (8) 72 °C for 10 s, (9) repeat steps (6-8) 38 times, (10) 72 °C for 2 min, (11) 10 °C for 2 min, (12) End. 7-8 µL of the resultant PCR products and a 100 bp to 2000 bp ladder (Thermo Fisher catalog number: 15628050) for reference were mixed with 1.4-1.5 µL of loading dye (Thermo Scientific catalog number: R0611), loaded on a 2% agarose gel with 2.5 µL of ethidium bromide (stock solution: 10 mg/mL), and visualized. Resultant bands (amplicon sizes) per Jackson Lab Website were as follows: mutant (m/m) = ~200 bp, heterozygote (m/+) = ~200 bp and 297 bp, and WT (+/+) = 297 bp. Genotyping identified a total of 5 heterozygous mice (experiment 3: n = 5), all of which were excluded from the analyses.
Statistics
Mean values of activity onset, onset variance, circadian period, and total activity counts were evaluated using analysis of variance (ANOVA); the F statistic is robust to violations of sample size inequality or normality (Lindman, 1974). To control for alpha inflation and type I error, pairwise comparisons which were performed using 2-tailed t tests were justified by a significant omnibus F statistic, except in instances of a priori planned comparisons. Paired, 2-tailed t tests were used to evaluate changes in FFT power within groups over 2 or more time points. Survival analyses were performed by generating Kaplan-Meier survival plots of the latency to arrhythmia onset in DD of Per2m/m mice, followed by logrank post hoc tests. All statistical comparisons were performed using StatView software (SAS), except in cases of nonparametric Fisher’s exact tests, which were performed using an online calculator (available at https://www.socscistatistics.com). Differences were considered significant if p < 0.05.
Results and Discussion
Experiment 1: Sex Differences in the Necessity of mPer2 for Expression of Circadian Rhythms
To examine the circadian role of mPer2 in female and male mice, activity rhythms of mice homozygous for the Per2Brdm1 mutation (Per2m/m mice; females, n = 5; males, n = 5) and of WT controls (females, n = 7; males, n = 7) were monitored using passive infrared detectors (Figure 1a-1d). WT mice became active ~30 min before dark onset, whereas mPer2 mutants began activity several hours earlier (ANOVA: F1,20 = 51.8, p < 0.0001; Figure 1e). In addition, entrainment to the LD cycle was considerably less stable in mutant compared with WT mice (F1,20 = 20.9, p < 0.0005; Figure 1f).
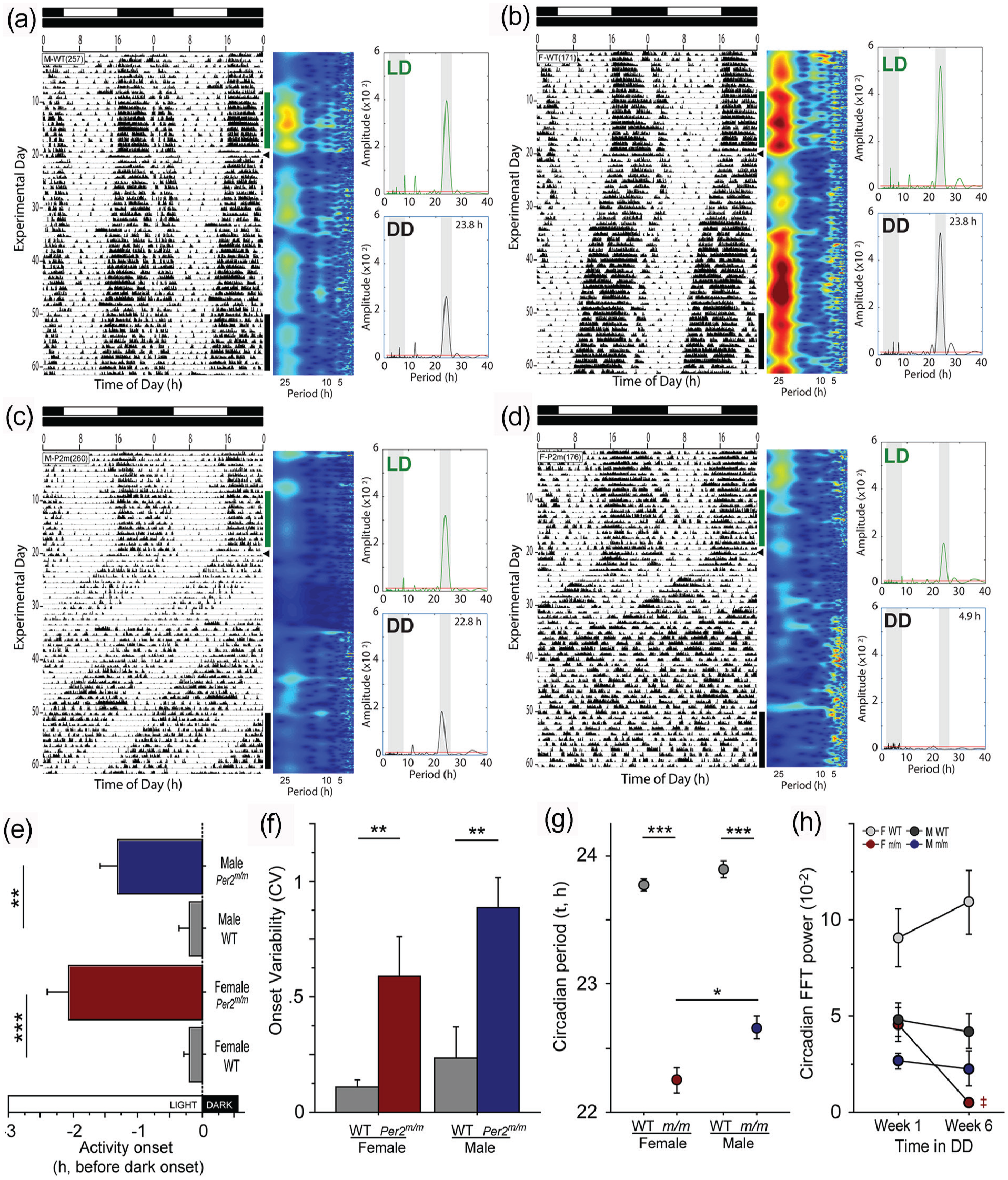
Sex differences in circadian behavior of Per2m/m mice. Representative double-plotted actograms of (a) WT male, (b) WT female, (c) Per2m/m male, and (d) Per2m/m female mice housed in a 12L:12D (LD) photocycle (shown at the top of each raster plot) for 3 weeks, then transferred to continuous darkness (DD; arrowhead indicates time of transfer) for 6 weeks. Home cage locomotor activity was monitored continuously using passive infrared (PIR) motion detectors. Abscissae indicate circadian time, successive days descend from the top, and vertical bars indicate 10-day epochs of LD (green) and DD (black) activity subjected to LSP analyses. To the right of each actogram, scalograms depict rhythmic power across circadian and ultradian period ranges (1-28 h), determined by continuous analytic wavelet transform. (e) Phase analysis: phase angle (timing) of activity onset in LD. (f) Stability analysis: circadian variability (
Following transfer to DD mutant mice exhibited free-running locomotor activity rhythms with very short circadian periods (τDD; Per2m/m: 22.46 ± 0.09 h, n = 10; WT: 23.83 ± 0.04 h, n = 14 [mean ± SEM]; ANOVA: F1,20 = 341.2, p < 0.0001; Figure 1g). Moreover, whereas in WT mice τDD was comparable between males and females (t12 = 1.43, p > 0.15), among Per2m/m mice τDD was significantly shorter in females (t8 = 3.11, p < 0.02; Figure 1g).
Prolonged exposure to DD revealed an additional sex difference in the Per2m/m chronotype. As expected, the short τDD expressed by Per2m/m mice was followed by a loss of the circadian rhythm of locomotor activity (Figure 1d). Unexpectedly, however, whereas 4 of 5 female mutants became arrhythmic during the first 6 weeks in DD, all 5 male mutants maintained robust free-running rhythms in constant darkness (Fisher’s exact test: p < 0.05; Figure 1c). To quantify changes in power of the circadian rhythm, as has been reported previously for this mutant (Zheng et al., 1999, 2001), an FFT was performed on activity during weeks 1-2 and weeks 5-6 in DD (Figure 1h). FFT power remained relatively higher over these intervals in WT mice and Per2m/m males, but decreased by nearly an order of magnitude in arrhythmic female mutants (Figure 1h; t3 = 5.67, p = 0.011), confirming a striking loss of circadian power. Based on visual examination of activity records, the latency to exhibit circadian arrhythmia varied from 14 to 29 days in Per2m/m females (e.g., Figure 1d). Arrhythmic female mice also exhibited a marked increase in FFT power in the ultradian range (Figure 1d; Zheng et al., 2001).
Experiment 2a: The Role of Per2m/m in Circadian Function Does Not Depend on the Method of Measurement
Two aspects of the Per2m/m chronotype support the inference that mPER2 is a core component of the mammalian circadian pacemaker: Per2m/m mice exhibit a markedly shortened period length and a loss of circadian rhythmicity in constant conditions (Albrecht et al., 2001; Bae et al., 2001; Zheng et al., 2001). Each of these Per2m/m chronotypes was sexually differentiated (cf. Figure 1g and 1h). To our knowledge, circadian arrhythmia in mPER2 mutants has only been documented in males (or in mice of unspecified sex; e.g., of the 8 Per2 studies cited in this report, 5 employed males and 3 did not specify sex), and primarily using cages equipped with RWs, which not only provide feedback onto the circadian system (Reebs and Mrosovsky, 1989) but also alter pacemaker period (Edgar et al., 1991) and amplitude (Schroeder et al., 2012). To interrogate whether the persistence of free-running rhythms in DD in male Per2m/m mice was a consequence of the absence of RWs, female and male Per2m/m and WT mice were housed with or without wheels and transferred from 12L:12D to DD (Figure 2a-2e). Again, τDD was shorter in Per2m/m than WT mice (genotype: F1,71 = 175.5, p < 0.0001; Figure 2f), and shorter in female than male mutants (t36 = 2.39, p < 0.025; Figure 2f). RW access did not impact the development of circadian arrhythmia: 9 of 18 Per2m/m mice (50%) housed with wheels and 13 of 21 housed without wheels (62%) became arrhythmic over the first 6 weeks in DD (χ2 = 0.56, p > 0.4; thus, data were pooled across wheel access groups). Sex, however, markedly affected the development of circadian arrhythmicity: consistent with data from experiment 1, 16 of 20 (80%) female mutants exhibited arrhythmia, whereas only 5 of 19 (26%) male mutants became arrhythmic during the first 6 weeks in DD (Fisher’s exact test: p < 0.005). Mice remained in DD for an additional 12 weeks, during which time the remaining 4 Per2m/m females, and an additional 7 Per2m/m males, became arrhythmic. In total, 12 of 19 (63%) male and 20 of 20 (100%) female mutants exhibited circadian arrhythmia over the course of 18 weeks in DD (Fisher’s exact test: p < 0.004). Survival analysis indicated a striking sex difference in the emergence of the arrhythmic chronotype (χ2 = 15.3, p < 0.001, logrank test; Figure 2g), with a mean (±SD) latency to arrhythmia of 36.3 ± 19 days in females versus 52.1 ± 24 days in males (t30 = 2.06, p < 0.05). To further characterize rhythmic and arrhythmic chronotypes, activity during the 2 weeks immediately after the transition from rhythmicity to arrhythmicity was evaluated, again using FFT: in both sexes the loss of behavioral circadian rhythmicity was characterized by a log-scale reduction in FFT power (female Per2m/m: t18 = 3.70, p < 0.005; male Per2m/m: t11 = 4.71, p < 0.001); FFT power did not decrease significantly over a yoked interval in female WT mice (t19 = 1.33, p > 0.15) but did decrease among male WTs (t16 = 2.63; p = 0.018; Figure 2h) and a redistribution of rhythmic power into ultradian frequency bands (Figure 2a-2e); LSP analyses identified comparable decreases in rhythmic power in Per2m/m mice (females: t18 = 4.21, p < 0.001; males: t11 = 4.71, p < 0.001) and no significant decreases in rhythmic power in WT mice (females: t19 = 1.69, p > 0.10; males: t16 = 2.06, p > 0.05). Finally, the onset of circadian arrhythmia did not bear any obvious relation to identifiable external stimuli: all mice were housed in the same room for the entire experiment, and DD treatment was initiated on the same calendar date, yet behavioral arrhythmia among 32 mice (20 F, 12 M) began on 22 unique days over an 87-day interval, and the onset of arrhythmicity did not appear to occur at an obvious phase of the circadian cycle.
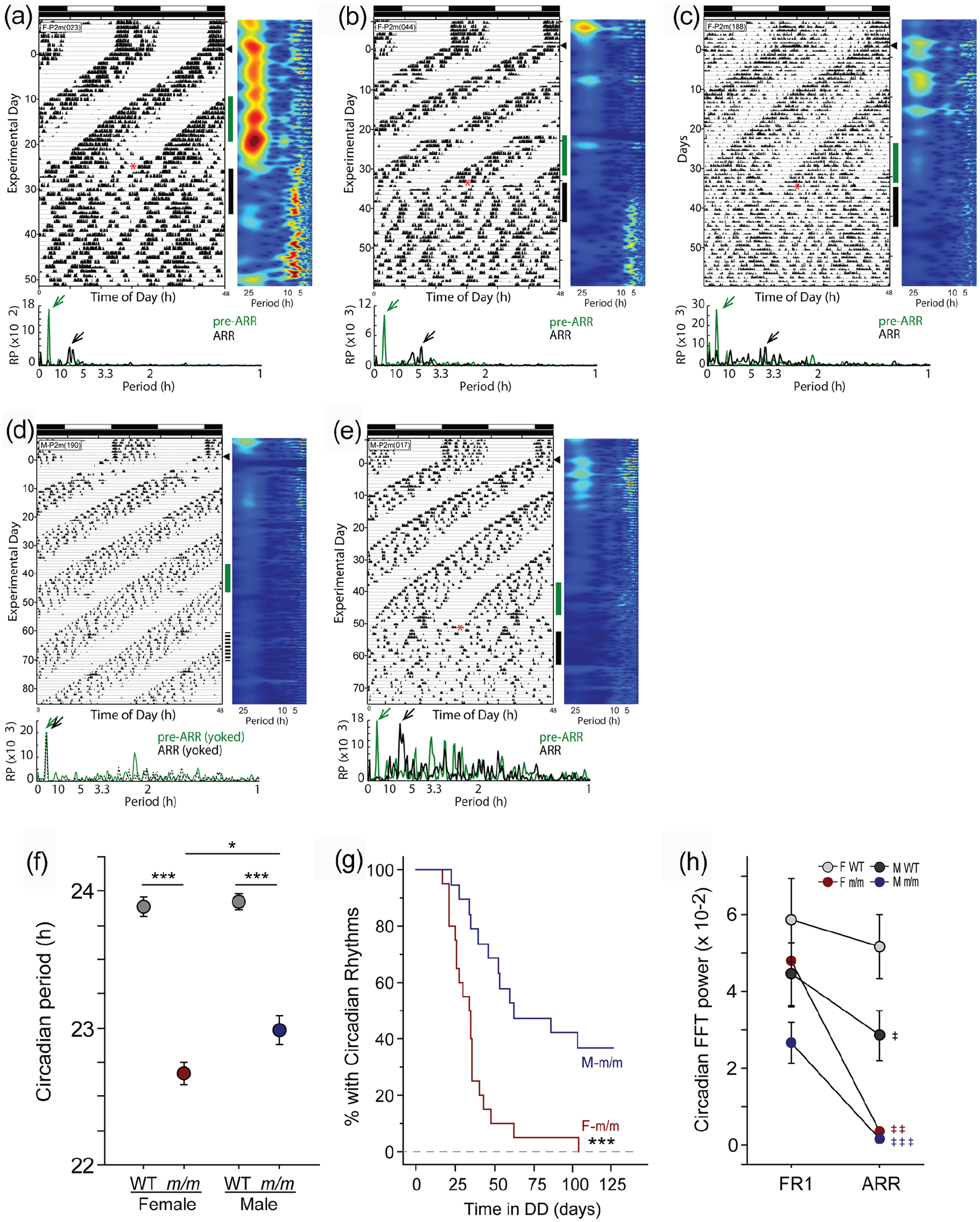
Uniform loss of circadian rhythms in female but not male Per2m/m mice. Double-plotted actograms of representative (a-c) female and (d-e) male Per2m/m mice housed in LD and transferred to DD (arrowhead, conventions as in Figure 1). Home cage locomotor activity was monitored continuously using RWs (a, b, d, e) or PIR motion detectors (c). Vertical green bars alongside activity data indicate 10-day intervals just prior to the onset of circadian arrhythmia (pre-ARR; green) and just after arrhythmia onset (ARR; black) subjected to FFT analyses, which appear beneath each actogram (arrowheads indicate peak FFT value). The mouse in panel d did not exhibit arrhythmicity; instead, activity was analyzed during a yoked interval of FR activity (dashed line). (f) Periodogram analysis: free-running circadian period (τ) among male and female WT and Per2m/m mice during initial exposure to DD as determined by Lomb-Scargle periodogram. (g) Survival analysis: Kaplan-Meier survival plot evaluating the emergence of circadian arrhythmia in DD among male and female Per2m/m mice (12/19 male and 20/20 female Per2m/m mice exhibited arrhythmia in DD). (h) Fourier analysis: relative power in the circadian range (20-26 h) of the FFT prior to (FR1) and after (ARR) the onset of circadian arrhythmia in male and female Per2m/m mice, confirming the arrhythmia classification. Abbreviations: FR = free-running; RW = running wheels; PIR = passive infrared motion detectors; ARR = arrhythmia; FFT = fast Fourier transform; WT = wild-type. *p < 0.05. ***p < 0.001. ‡p < 0.05. ‡‡p < 0.01. ‡‡‡p < 0.001 versus FR1 value.
Despite the widespread use of Per2 mutants in chronobiology research, this sex difference has not been reported previously. In earlier reports (Zheng et al., 1999, 2001), 100% of Per2Brdm1 mice were reported to exhibit arrhythmicity after 2-18 days in DD. Such rapid and global development of arrhythmia in the Per2Bdrm1 mutant, combined with the lack of examination and analysis by sex, likely prevented the recognition of this categorical sex difference in the dependence of circadian rhythmicity in DD on PER2. Per2m/m mice in the present report (bearing the Per2Bdrm1 mutation) were first generated on a B6;129S background (Zheng et al., 1999), but then backcrossed for 5-6 generations with C57 mice, and are therefore 97%-98% C57BL/6 (Wang et al., 2009). (Additional information at http://www.informatics.jax.org.) Genetic background may impact chronotype expression in a trait-specific manner; for example, Per2ldc mutants crossed to a C57BL/6 background exhibited a short τ (~22 h), but not the rapid-onset circadian arrhythmicity typical of the Per2ldc mutation (Bae et al., 2001; Xu et al., 2007); however, behavior in DD was only examined for 14-18 days, which, on a B6 background, may be too short an interval for the arrhythmic chronotype to emerge (cf. Figure 2g). The present data would be consistent with the conjecture that genomic background delays the onset of arrhythmicity in Per2 mutants, unmasking a previously unrecognized categorical sex difference in the dependence on Per2 for circadian rhythmicity in constant darkness.
The Per2m/m mutation interacts with biological sex to generate a sexually differentiated circadian phenotype. Arrhythmia in Per2 mutants manifests synchronously in at least 2 circadian traits (spontaneous locomotor activity and wheel running), suggesting that the entire circadian network is arrhythmic. These data underscore the well-established critical role of PER2 in the generation and maintenance of circadian rhythms (Bae et al., 2001; Zheng et al., 2001), but indicate that compensation for the absence of a functional PER2 protein is considerably more robust in males.
Experiment 2b: Per2m/m Mice Spontaneously Recover Circadian Rhythmicity
Surprisingly, Per2m/m mice did not remain arrhythmic indefinitely in DD. Instead, the majority of circadian-arrhythmic mutants spontaneously recovered coherent, free-running circadian rhythms in DD when behavioral chronotyping was extended longer than in previous studies (Figure 3a-3d). Females were categorically superior to males in re-establishing behavioral circadian rhythms: 19 of 20 (95%) Per2m/m females, but only 4 of 12 (33%) males, recovered free-running rhythms after a bout of arrhythmicity (Fisher’s exact test: p < 0.0005). Spontaneous recovery of circadian rhythmicity also occurred more rapidly in females: the mean latency to recovery (i.e., the duration of circadian arrhythmia) was 38.6 ± 16 days in females and 69.5 ± 15 days in males (t21 = 3.6, p < 0.005; F range: 12-69 days; M range: 51-85 days; Figure 3e). As with the induction of arrhythmia, the recovery of circadian rhythmicity occurred in mice housed with (11 of 18; 61%) and without (12 of 21; 57%) RWs and was not linked to any obvious external stimuli: in 23 mice, circadian rhythms re-appeared on 18 different days over an 83-day interval; rhythm re-emergence did not consistently occur at any specific time of day. Circadian rhythmicity re-emerged as abruptly as it disintegrated: in the modal phenotype, a consolidated inactive phase emerged via a reduction in activity levels over 1-2 circadian cycles (e.g., Figure 3b). In neither sex did the latency to arrhythmia predict arrhythmia duration (p > 0.8, both comparisons). Circadian period or power (FFT or LSP) during the rhythmic interval in DD did not predict the latency to (p > 0.05, all comparisons), or persistence of (p > 0.05, all comparisons), arrhythmia in DD, nor did circadian power during the arrhythmic interval of DD predict the duration of arrhythmia (p > 0.3, all comparisons).
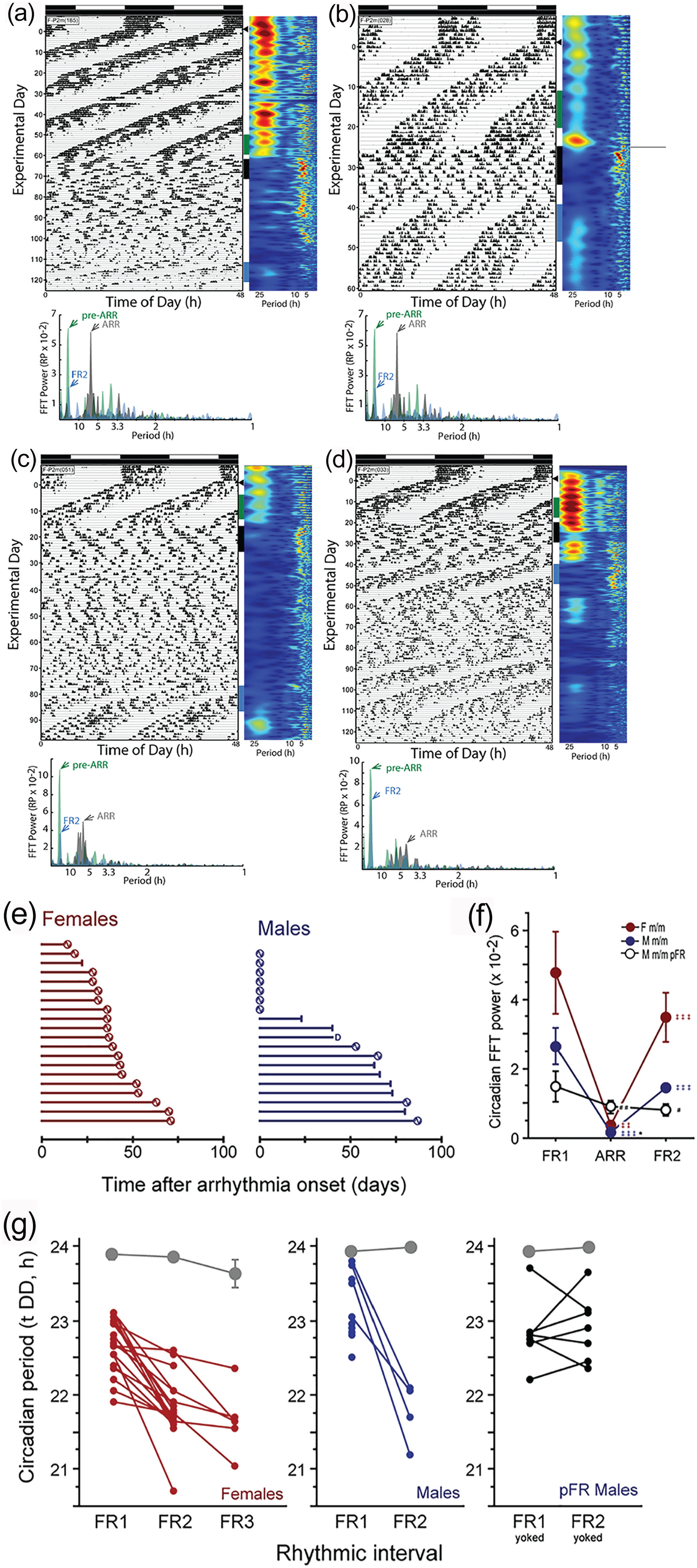
Spontaneous recovery of circadian rhythms in Per2m/m mice. Representative double-plotted RW actograms of female mice that exhibited circadian arrhythmia in DD and subsequently recovered free-running rhythms. Mice in panels a-c exhibited a single bout of arrhythmia, weeks to months in duration; the mouse in panel d exhibited multiple bouts of arrhythmia and recovery of rhythmicity. Vertical bars indicate free-running locomotor activity occurring just prior to ARR (green), during ARR (black), and following recovery of circadian rhythmicity (FR2; blue); these intervals were subjected to FFT analyses, shown beneath each actogram. (e) Idiographic recovery plots: horizontal bars indicate the number of days elapsed between the onset of circadian arrhythmia and either the recovery of free-running circadian rhythms (circles) or the end of the experiment (bars). One mouse died prior to the end of the experiment (indicated as “D”); 7 of 19 male Per2m/m mice did not exhibit arrhythmia. (f) Fourier analysis: relative power in the circadian range (20-26 h) of the FFT prior to (FR1) and after (ARR) the onset of circadian arrhythmia, and after recovery of free-running (FR2) circadian activity rhythms, confirming the FR2 classification. Per2m/m males that did not exhibit arrhythmicity are designated “persistent free-running” (pFR) mice. Due to the small number of males that recovered rhythms, we performed a supplemental LSP analysis to confirm this pattern. LSP confirmed increases, albeit more modest, in power in Per2m/m mice that recovered circadian rhythms (females: t18 = 4.85, p < 0.001; males: t3 = 2.75, p = 0.071); LSP power increased 3.4- to 7.9-fold during the transition from ARR to FR2, restoring in females and males, respectively, 58% and 56% of power that had been present in the LSP of the initial (FR1) free-running rhythm. (g) Periodogram analysis: circadian period (τ) of the initial episode of free-running circadian locomotor activity in DD (FR1) and of the free-running circadian rhythm after recovery from circadian arrhythmia (FR2). Note: 15 of 19 females exhibiting FR2 exhibited a second bout of arrhythmia; and 5 of these 15 females exhibited a subsequent cycle of recovery (indicated as FR3). In panel f: *p < 0.05 versus female value; #p < 0.05, ##p < 0.01 versus all other groups at same time point; ‡‡p < 0.01, ‡‡‡0.001 versus previous time point, within group. Abbreviations: RW = running wheels; ARR = arrhythmia; FFT = fast Fourier transform; pFR = persistent free-running; LSP = Lomb-Scargle periodogram.
Finally, the reconstituted circadian rhythm differed from that exhibited prior to the development of arrhythmia. FFT power increased 8- to 14-fold during the transition from arrhythmicity to free-running circadian recovery (FR2); restoring in females and males, respectively, 74% and 82% of power that had been present in the FFT of the initial (FR1) free-running rhythm (Figure 3f). In addition, a consistent feature of the reconstituted circadian pacemaker was its shorter τ: in every individual that exhibited circadian recovery τFR2 was shorter than τFR1 (females: t17 = 6.86, p < 0.001; males: t3 = 6.11, p < 0.01; Figure 3g). Indeed, 6 female mice with reconstituted circadian rhythms exhibited a second bout of arrhythmicity, followed by yet another recovery of coherent free-running rhythmicity (FR3), and in each of these individuals τFR3 < τFR2 < τFR1 (Figure 3g).
Experiment 3: Gonadectomy Promotes Circadian Arrhythmia in Male Per2m/m Mice
Prior investigations have revealed marked activational influences of circulating gonadal hormones, in particular androgens, on the circadian system, notably on the consolidation and amplitude of circadian locomotor activity (Daan et al., 1975; Iwahana et al., 2008; Yan and Silver, 2016). In light of the prominent sex differences described here, we examined the effects of gonadal hormones on DD-induced circadian arrhythmia in Per2m/m mice. Adult mice in 12L:12D were surgically gonadectomized (females: OVx, males: GDx) or sham-operated (n = 9-10/group), and after recovery from surgery all mice were transferred to DD for >100 days and monitored with RWs. As expected (Iwahana et al., 2008; Bailey and Silver, 2014; Yan and Silver, 2016), gonadectomy markedly reduced locomotor activity (F1,45 = 9.14, p < 0.005; Suppl. Fig. S1A), with significant effects in Per2m/m females (t18 = 2.24, p < 0.05), but not in Per2m/m males (t17 = 0.51, p > 0.6). Upon transfer to DD, Per2m/m mice also exhibited the expected short τ (F1,45 = 8.68, p < 0.01; Figure 4a-4d); although τ during the initial interval of exposure to DD did not differ between male and female Per2m/m mice (t17 = 0.076, p > 0.9; Suppl. Fig. S1B). GDx did not affect τ in WT mice of either sex (F: t7 = 0.86, p > 0.40; M: t3 = 1.67, p > 0.15).
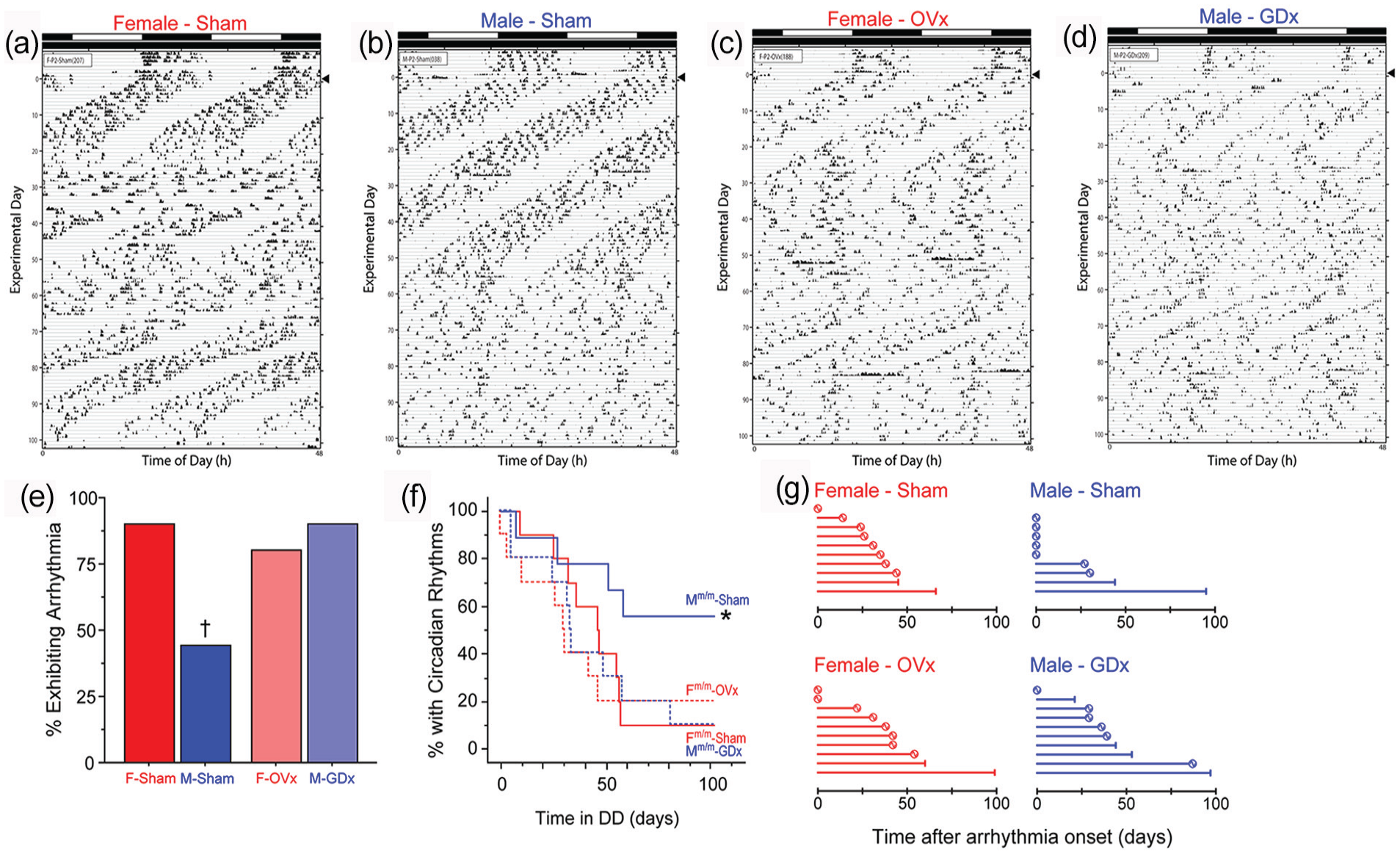
Castration promotes circadian arrhythmicity in Per2m/m males. Representative double-plotted RW actograms of female and male Per2m/m mice that were sham-operated (female: panel a; male: panel b) or gonadectomized (OVx female: panel c; GDx male: panel d), and maintained in DD for >100 days (transfer from LD to DD indicated by arrowhead; actogram conventions as in Figure 1). See Supplementary Figure S1 for representative actograms of WT control mice, which did not exhibit circadian arrhythmia. (e) Percentage of Per2m/m mice of each sex and surgical condition that exhibited circadian arrhythmia in DD. (f) Survival analysis: Kaplan-Meier survival plot evaluating the emergence of circadian arrhythmia in DD among male and female WT and Per2m/m mice. (g) Idiographic recovery plots: horizontal bars indicate the number of days elapsed between the onset of circadian arrhythmia and either the recovery of free-running circadian rhythms (circles) or the end of the experiment (vertical bars). Recovery of free-running circadian rhythms was evident in 7 of 9 sham-females, in 2 of 4 sham-males, in 6 of 8 OVx females, and in 5 of 9 GDx males. Abbreviations: RW = running wheels; OVx = ovariectomized; GDx = orchiectomized; WT = wild-type. *p <0.05 vs. F-Sham and vs. M-GDx. †p < 0.06 vs. F-Sham and vs. M-GDx.
WT mice, regardless of surgical treatment, did not exhibit circadian arrhythmia (Suppl. Fig. S1C-S1F). As before, the vast majority of gonad-intact females exhibited arrhythmia (9 of 10), whereas fewer than half (4 of 9) of the intact males did so (90% vs 44%; Fisher’s exact test: p = .057; Figure 4e). Ovariectomy (OVx) did not meaningfully alter the likelihood of arrhythmia in female Per2m/m mice: 8 of 10 OVx females exhibited at least one bout of circadian arrhythmia in DD (Fisher’s exact test: p > 0.9 vs sham-operated females; Figure 4e). In Per2m/m males, castration (GDx) caused a striking increase in circadian arrhythmia: 9 of 10 GDx males exhibited arrhythmia in DD (Fisher’s exact test: p = 0.057 vs sham-males; Figure 4e)—an incidence comparable with that of sham- and OVx females (Fisher’s exact test: p > 0.9, both comparisons; Figure 4e). Overall, arrhythmia in 30 animals occurred on 25 unique days.
Survival analyses indicated that among gonad-intact mice, sex again significantly impacted the development of circadian arrhythmia (χ2 = 4.07, p < 0.05), and among males, GDx markedly altered the emergence of arrhythmia over time (χ2 = 4.13, p < 0.05; Figure 4f). Gonad-intact males also tended to differ from OVx females (χ2 = 3.44, p = 0.06), but circadian arrhythmia evolved comparably in DD among all other groups (p > 0.60, all comparisons; Figure 4f). Circadian arrhythmia occurred more rapidly in this study compared with the previous experiment, and the mean latency to arrhythmia did not differ between the 4 intact males (35.8 ± 12 d) and 9 intact females (40.3 ± 5 d; t11 = 0.41, p > 0.6) that became arrhythmic. Taken together this pattern of results suggests that the sex difference in circadian arrhythmia in the Per2m/m mouse is mediated in part by concurrent effects of male gonadal hormones and that ovarian hormones are not required for high rates of expression of arrhythmia in females.
As in the prior experiment, many (20 of 38; 53%) arrhythmic Per2m/m mice also exhibited spontaneous recovery from circadian arrhythmia (Figure 4g). Recovery was evident in 78% (7 of 9) and 75% (6 of 8) of intact and OVx females, respectively. Just 50% (2 of 4) of intact males that exhibited arrhythmia recovered, and 56% (5 of 9) arrhythmic GDx males recovered. These 20 recoveries occurred on 15 unique days. Sample sizes were not sufficient to permit statistically powerful categorical analyses of the incidence of recovery, but the patterns suggest (1) again an increased likelihood of spontaneous recovery in females, (2) no obvious augmentation of recovery in GDx males, and (3) no effect of OVx on recovery in females (Figure 4g). Circulating male gonadal hormones may play a role in the induction of arrhythmia, but not circadian recovery, whereas circulating female gonadal hormones are not mandatory for either. Organizational effects of gonadal hormones in both sexes may be implicated in the latter. Whether male and female gonadal hormones act via androgen or estrogen receptors in the mediation of these phenomena requires further investigation (Vida et al., 2008; Yan and Silver, 2016).
General Discussion
Mice lacking a functional copy of the Per2 gene exhibited an atypical pattern of entrainment and largely failed to sustain circadian rhythms in multiple measures of general locomotor activity (RW activity, PIR activity) in constant darkness, confirming and extending the critical role for PER2 in the molecular transcription and translation feedback loop (TTFL) that comprises the circadian pacemaker. The Per2m/m circadian phenotype was exaggerated in female mice: two core features of the Per2m/m chronotype—shortened circadian period and loss of circadian rhythmicity in DD—were markedly enhanced in females compared with males. Females were far more likely to become arrhythmic in DD, and circadian arrhythmia occurred many weeks earlier in females than males. Moreover, once circadian arrhythmia emerged, most (8 of 12) males remained in this state; in contrast, nearly all (19 of 20) females recovered from circadian arrhythmicity, re-establishing coherent free-running circadian rhythms. Indeed, several (6 of 19) female mice repeated this arrhythmia/recovery cycle multiple times. Whereas many males can sustain circadian rhythms in behavior without a functional PER2 protein, few females can. The data identify an unstable state of equilibrium in the absence of a functional PER2 protein, characterized by sequential fluctuations between transient states of coherent circadian rhythmicity and arrhythmicity.
Restoration of rhythmicity in Per2 mutant mice was previously reported (Steinlechner et al., 2002; Abraham et al., 2006). Double mutant Per2/Cry1 mice exhibited loss of circadian rhythms in wheel running activity in DD and the emergence of robust ultradian rhythms. Exposure to high-intensity constant illumination (LL) restored circadian rhythms with a much shorter period (Abraham et al., 2006). We also observed emergence of strong ultradian rhythms in association with DD arrhythmicity, but circadian reinstatement occurred in DD in the absence of light signals. Whether the relay of timing information from ultradian rhythms to the suprachiasmatic nuclei is critical for reestablishment of circadian rhythms, as posited by Abraham et al. (2006) remains to be verified. Steinlechner et al. (2002) reported rapid development of arrhythmicity in Per2m/m mice held in DD; subsequently, under constant light, these mice exhibited robust circadian activity and body temperature rhythms with short period lengths, an outcome consistent with the original description of this mutant model, in which a 6-h light pulse immediately restored circadian rhythmicity in 100% of mice (Zheng et al., 1999). Collectively, these findings suggest that residual potentially unstable clock function in Per2m/m mice supports coherent circadian rhythms in DD as well as in LL, with interspersed circadian arrhythmicity.
PER2 plays a prominent role in the generation of cellular circadian rhythms (Zheng et al., 1999; Bae et al., 2001; Zheng et al., 2001); mice with loss-of-function mutations in the Per2 gene (e.g., Per2Brdm1 and Per2ldc) exhibit 2 essential chronotypes in DD: a short τ (22-23 h) and ubiquitous loss of circadian rhythmicity (Zheng et al., 1999; Bae et al., 2001). The present data identify quantitative and categorical sex differences in each of these chronotypes: females exhibited a much shorter τ, and all females lost circadian rhythmicity, but only approximately half of all males did. PER2 is indispensable for the generation of circadian rhythms in peripheral circadian oscillators, but network dynamics within the SCN introduce compensation or redundancy that allow persistent circadian rhythmic output by the SCN in the absence of a functional mPER2, and thus transient rhythmicity in Per2m/m fibroblasts in DD (Liu et al., 2007). As in other reports, circadian arrhythmicity did not occur immediately upon transfer to DD; instead, Per2m/m mice exhibited free-running rhythms that lasted from 12 days to 18 weeks, supporting previous conclusions that the Per2Brdm1 mutant has a partially functional clock (Albrecht et al., 2001; Zheng et al., 2001; Zhang et al., 2019). mPer1 mRNA rhythms were attenuated but not eliminated in Per2Brdm1 mice (Zheng et al., 1999), suggesting that residual levels of mPER1 provided only partial compensation for the absence of mPER2, which may account for the rapid onset, but not instantaneous, arrhythmicity in DD. As compared with prior reports in B6;129 hybrid Per2 mutants, rhythmicity was sustained by Per2m/m mice for much longer intervals in DD, although arrhythmia onset occurred somewhat earlier in experiment 3 (Figure 4) compared with the previous experiment (Figure 3); regardless, in many males, the circadian pacemaker remained functionally robust for months, and perhaps indefinitely (Figure 2g). Given that the PERIOD ortholog mPer3 does not compensate for mPer2 deficiency (Bae et al., 2001), the present results suggest that in the Per2m/m mutant, severely dampened mPER1 rhythms may afford a more potent compensation for the absence of mPER2 in the substantial population of males that exhibit persistent free-running rhythms in DD. A similar degree of compensation is categorically absent in females.
Circadian arrhythmicity was not permanent; rather, free-running locomotor activity rhythms spontaneously re-emerged in arrhythmic mice (Figure 3a-3d). It was not possible to determine, from activity records alone, whether the loss and subsequent gain of circadian rhythmicity merely reflected a transient uncoupling of the circadian pacemaker from output pathway(s) that mediate overt rhythmicity, or whether the pacemaker was arrested during behavioral arrhythmicity. Activity onset of FR2 (the second, recovered, free-running rhythm) did not resume at the same time of day that activity onset of FR1 had last been expressed, which would be predicted if the circadian pacemaker had become arrested at the end of FR1 and subsequently restarted in FR2. Alternatively, if the circadian pacemaker had continued to keep time during the interval of behavioral arrhythmia, then the phase of the restored rhythm ought to be predictable by extrapolation of the phase of the rhythm prior to the loss of rhythmicity. However, because τFR1 often lengthened in the days leading up to arrhythmicity, it was not possible to clearly extrapolate onset phase between FR1 and FR2. Further complicating such an extrapolation, τ changed markedly in mice that recovered from arrhythmia. Further characterization of this novel model of circadian instability (e.g., via longitudinal evaluation of the molecular TTFL within the SCN) may afford direct insight into whether the circadian pacemaker continues to oscillate during the interval of arrhythmia.
The propensity of most female and many male Per2m/m mice to fluctuate between states of coherent circadian rhythmicity and arrhythmicity has not been reported previously, likely because the 18 weeks of recording employed herein is, to our knowledge, the longest interval over which mice bearing mutations in any of the core circadian clock genes have been continuously monitored. Similar long-term monitoring of behavior has been performed in rodents and primates bearing SCN lesions, with no evidence of functional rhythm recovery (Mosko and Moore, 1978). In contrast, spontaneous recovery of circadian pacemaker expression was pervasive in Per2m/m mice, occurring in 95% of females and 33% of males, indicating that the state of circadian arrhythmia induced by the Per2Brdm1 mutation is not permanent. This categorical difference suggests that circadian disruptions induced by genomic mutation may not be isomorphic with those induced by chemical or physical lesions of the anatomical pacemaker. During transient arrhythmicity, molecular circadian rhythms may be eliminated at the level of individual SCN oscillatory neurons; alternatively, cells within the SCN may continue to oscillate individually during transient arrhythmia, but become transiently uncoupled (desynchronized) from one another at a cellular (Ohta et al., 2005) or network (Evans et al., 2013) level, or uncoupled from SCN output pathways. Irrespective of such formal features, the present data indicate that compensation among molecular elements of the circadian TTFL is more robust than previously described. Moreover, these data suggest that compensation for the absence of a functional PER2 protein is strongly sexually differentiated and mediated, at a minimum, by effects of gonadal hormones. Indeed, the effects of orchidectomy and ovariectomy on arrhythmia in DD are consistent with several reports indicating robust effects of GDx relative to OVx in adult rodents (Daan et al., 1975; Morin et al., 1977; Iwahana et al., 2008; Yan and Silver, 2016). The relative importance of androgen and estrogen receptors in mediating effects of gonadal hormones on arrhythmia in Per2m/m mice remains to be determined, however.
Per2m/m females undergo reproductive aging faster than males (Pilorz and Steinlechner, 2008), and we do not know how long-term DD affects estrous cycling.Given the impact of female reproductive hormones on circadian rhythms in general and Per2 in particular (Nakamura et al., 2005, 2008), processes associated with reproductive aging may contribute to the sex difference in arrhythmia identified here or in part mask the effect of loss of estrous cycling (Pilorz et al., 2020). Pilorz and Steinlechner (2008) identified reproductive senescence in female Per2m/m mice as beginning sometime after 6 months of age, as 9- to 12-month-old Per2m/m females had reduced fertility compared with WT controls. Aging also decreased overall levels of locomotor activity in acyclic female Per2m/m mice, but a similar effect of aging was observed in WT controls; however, the phase angle of entrainment to the LD cycle was comparable in aged and young Per2m/m mice (Pilorz et al., 2009). In experiment 2, some female Per2m/m mice would have entered this window of reproductive senescence during exposure to DD, whereas in experiment 3, female Per2m/m had already aged well into, if not beyond, this window before being subjected to DD. The comparable rates of arrhythmia observed among females in experiments 2 and 3 (100% and 90%, respectively), and the arrhythmia observed in male mice (albeit a lower incidence), together argue against aging of the reproductive system per se playing a categorical role in DD-induced arrhythmia. However, the different latencies to arrhythmia onset documented across the 2 experiments may indeed be related to such aging. The effects of long-term DD on estrous cycles of Per2m/m mice are not known. The lack of an effect of ovariectomy on arrhythmia in experiment 3 may be related to the relatively advanced age of female mice in this study; although effects of OVx on overall activity were still evident. The circadian instability in Per2m/m mice characterized here may offer a useful model for further investigations of the role of aging and hormones on circadian biology and behavior.
Behavioral genomic assays used in recent decades to identify circadian clock genes in mice typically examine free-running locomotor activity for ≤21 days in DD (Siepka and Takahashi, 2005; Siepka et al., 2007). Assays of this length are inherently biased toward genes that markedly alter τ and induce a rapid loss of circadian rhythmicity (Siepka et al., 2007). The identification here of circadian chronotypes which emerge only after much longer intervals of exposure to DD indicates that such assays are less robust in identifying genes that regulate the circadian system, or in a sexually differentiated manner. Moreover, even core molecular components of the circadian TTFL likely have more complex roles than currently recognized.
Supplemental Material
sj-pdf-1-jbr-10.1177_07487304211060896 – Supplemental material for Spontaneous Recovery of Circadian Organization in Mice Lacking a Core Component of the Molecular Clockwork
Supplemental material, sj-pdf-1-jbr-10.1177_07487304211060896 for Spontaneous Recovery of Circadian Organization in Mice Lacking a Core Component of the Molecular Clockwork by Jonathan P. Riggle, Kenneth G. Onishi, Jharnae A. Love, Dana E. Beach, Irving Zucker and Brian J. Prendergast in Journal of Biological Rhythms
Footnotes
Acknowledgements
We thank Lance Kriegsfeld for comments on a draft of the manuscript, two anonymous reviewers for suggestion that improved the manuscript, Andrew Maneval for technical assistance, and Drs Betty Theriault and Darya Malhiot for expert veterinary care. This research was supported by NIH grant AI-67406 and the Social Sciences Division of the University of Chicago.
Conflict of Interest Statement
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
NOTE
Supplementary material is available for this article online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.