
Abstract
Reverse genetic studies have revealed that common clock genes, such as period (per), timeless (tim), cycle (cyc), and Clock (Clk), are involved in the circadian clock mechanism among a wide variety of insects. However, to what degree the molecular oscillatory mechanism is conserved is still to be elucidated. In this study, cDNA of the clock gene Clk was cloned in the cricket Gryllus bimaculatus, and its function was analyzed using RNA interference (RNAi). In adult optic lobes, the Clk mRNA level showed no significant rhythmic changes both under light-dark cycle (LD) and constant darkness (DD). A single injection of Clk double-stranded RNA (dsRNA) resulted in a knockdown of the mRNA level to about 25% of the peak level of control animals. The injected crickets lost their locomotor rhythms in DD. The arrhythmicity in locomotor activity persisted for up to 50 days after the Clk dsRNA injection. Control animals injected with DsRed2 dsRNA showed a clear locomotor rhythm like intact animals. Injection of Clk dsRNA not only suppressed the mRNA levels of both per and tim but also abolished their rhythmic expression. per RNAi down-regulates the Clk mRNA levels, suggesting that per is required for sufficient expression of Clk. These results suggest that Clk is an essential component and plays an important role in the cricket’s circadian clock machinery like in Drosophila, but regulation of its expression is probably different from regulation in Drosophila.
The circadian oscillation is produced by cyclical expressions of so-called clock genes through feedback mechanisms (Dunlap, 1999; Stanewsky, 2002; Tomioka and Matsumoto, 2010). In insects, the molecular oscillatory mechanism has been studied in a variety of species, although most extensively studied is the fruit fly Drosophila melanogaster. The common components in the mechanism are period (per), timeless (tim), cycle (cyc), and Clock (Clk). In Drosophila, the Clk gene works as a key element in the clock machinery together with cyc (McDonald and Rosbash, 2001; Zhao et al., 2003; Kilman and Allada, 2009): CLK forms a heterodimer with CYC and activates the transcription of per and tim during late day to early night (Allada et al., 1998; Rutila et al., 1998). The resultant PER and TIM proteins accumulate in the cytoplasm during the night and form a PER/TIM heterodimer, entering the nucleus to repress their own transcription through inactivation of CLK/CYC (Darlington et al., 1998; Dunlap, 1999). This reduces the PER and TIM levels and therefore releases CLK/CYC from repression to reactivate the transcription of per and tim, proceeding the loop to the next round. CLK/CYC also activates the transcription of other clock genes, vrille (vri) and Par domain protein 1ε (Pdp1ε) (Cyran et al., 2003; Glossop et al., 2003). vri mRNA peaks prior to the peak of Pdp1ε mRNA, and the VRI protein, thus accumulating earlier, represses Clk transcription, while PDP1ε later stimulates the transcription of Clk by competitive binding to the VRI box with VRI. Through this mechanism, Clk is expressed rhythmically with a peak at late night to early day.
The Clk gene has been cloned or found in the genome in a variety of insects, including honey bees, mosquitoes, moths, monarch butterflies, bean bugs, desert locusts, and firebrats (Chang et al., 2003; Rubin et al., 2006; Ikeno et al., 2008; Shao et al., 2008; Zhu et al., 2008; Gentile et al., 2009; Kamae et al., 2010; Rund et al., 2011; Tobback et al., 2011). However, there are only a few instances for its functional analysis. One of the reasons for the lack of analysis may be that molecular biological techniques are usually difficult to apply in nonmodel insect species. We recently found that RNA interference (RNAi) is particularly effective for functional analysis of clock genes in some insects including crickets and firebrats (Tomioka et al., 2009; Kamae et al., 2010). In the cricket Gryllus bimaculatus, the period (Gb’per) gene shows a rhythmic profile in its mRNA expression with a peak in the early part of the night before midnight, and knockdown of its mRNA by RNAi completely abolishes the circadian rhythm at the molecular, behavioral, and electrophysiological levels (Moriyama et al., 2008; Moriyama et al., 2009). Thus, it is likely that Gb’per plays an essential role in the clock machinery in G. bimaculatus. However, timeless (Gb’tim) seems nonessential, although it is rhythmically expressed like Gb’per. Knockdown of Gb’tim slightly changed the free-running period but never stopped the clock (Danbara et al., 2010), suggesting that the clock mechanism of the cricket differs from that of Drosophila.
In the present study, we have analyzed the role of the clock gene Clock (Gb’Clk) in the cricket (G. bimaculatus). We first obtained its cDNA by molecular cloning, examined the daily expression pattern of its mRNA using quantitative real-time RT-PCR (qPCR), and investigated its role in the rhythm generation with RNAi. The results showed that its knockdown by RNAi disrupted circadian locomotor rhythms and suppressed the expression levels of Gb’per and Gb’tim, suggesting that Gb’Clk plays an essential role as a transcriptional activator in the circadian clock in the cricket. Interestingly, Gb’Clk mRNA showed no daily expression, suggesting that Gb’Clk receives transcriptional regulation differently from that in the Drosophila clock.
Materials and Methods
Experimental Animals
The third and final (eighth) instar nymphal and adult male crickets (G. bimaculatus) were used. They were purchased or obtained from a laboratory colony maintained under standard environmental conditions with a lighting regimen of light-dark (LD) 12:12 (light: 0600-1800 h in Japanese standard time [JST]) and a constant temperature of 25 °C. They were fed laboratory chow and water.
Cloning and Structural Analysis of the Clock Gene
Total RNA was extracted with ISOGEN (Nippon Gene, Tokyo, Japan) from 20 heads of third instar nymphs collected at zeitgeber time (ZT) 10 (ZT 0 corresponds to lights-on and ZT 12 to lights-off). There was 5 µg of total RNA used for reverse transcription to obtain cDNA using SuperScript II (Invitrogen, Carlsbad, CA). Using the single-stranded cDNA as a template, we performed PCR with degenerate primers deduced from the conserved amino acid sequences among insect Clock homologs. The primers used were 5′-agaagaagcggcgggaYcaRttYaa-3′ for forward and 5′-gcgtaggacaccactgtgtgNgtRcaNac-3′ for reverse. The PCR conditions employed were 30 seconds for denaturation at 95 °C, 30 seconds for annealing at 57 °C, and 1 minute 30 seconds for extension at 72 °C for 35 cycles with ExTaq DNA polymerase (Takara, Otsu, Japan). The purified fragment was cloned into TOPO-pCR II vector (Invitrogen) and sequenced with BigDye Terminator v3.1 Cycle Sequencing kit (Applied Biosystems, Foster City, CA). 5′ and 3′ rapid amplification of cDNA ends (RACE) were performed with GeneRacer kit (Invitrogen) and SMARTer RACE cDNA Amplification kit (Takara) with gene-specific primers 5′-TAATACGACTCACTATAGGGAAGGGGTGTCTGTAATCTT-3′ and 5′-TCTTGCCACTACCGCTTTCT-3′, respectively. RACE fragments were purified, cloned, and sequenced as mentioned above. Sequences were analyzed by Genetyx version 6 (Genetic Information Processing Software, Tokyo, Japan) and BioEdit version 7.0.5 (Biological Sequence Alignment Editor, Ibis Biosciences, Carlsbad, CA).
CLOCK’s amino acid sequences were analyzed with ClustalW (http://clustalw.ddbj.nig.ac.jp/top-j.html). Sequences of known insect Clks were obtained from GenBank.
Measurement of mRNA Levels
qPCR was used to measure mRNA levels. Total RNA was extracted and purified from 6 or 20 adult male optic lobes with TRIzol Reagent (Invitrogen). To remove contaminating genomic DNA, the total RNA was treated with DNase I. About 500 ng of total RNA of each sample was reverse transcribed with random 6-mers using PrimeScript RT Reagent kit (Takara). Real-time PCR was performed by Mx3000P Real-Time PCR System (Stratagene, La Jolla, CA) using FastStart Universal SYBR Green Master (Roche, Tokyo, Japan) including SYBR Green with primers 5′-AATGACCGTAGTCGAGAAAGTGAAG-3′ and 5′-TTGCGATGATTGAGGTTGTTG-3′ for Gb’Clk, 5′-AAGCAAGCAAGCATCCTCAT-3′ and 5′-CTGAGAAAGGAGGCCACAAG-3′ for Gb’per (GenBank/EMBL/DDBJ accession no. BAG48878), 5′-CCAAAGACAGGAAGCAGACC-3′ and 5′-GAATCCCAACACCAAAATGG-3′ for Gb’tim (GenBank/EMBL/DDBJ accession no. BAJ16356), and 5′-GCTCCGGATTACATCGTTGC-3′ and 5′-GCCAAATGCCGAAGTTCTTG-3′ for Gb’rpl18a (GenBank/EMBL/DDBJ accession no. DC448653). The results were analyzed using the software associated with the instrument. The values were normalized with the values of Gb’rpl18a at each time point. Results of 3 or 8 independent experiments were used to calculate the mean ± SEM.
RNAi
Double-stranded RNA (dsRNA) for Gb’Clk, Gb’per, and DsRed2, derived from a coral species (Discosoma sp.), were synthesized using MEGAscript High Yield Transcription kit (Ambion, Austin, TX). For Gb’Clk and Gb’per, using cDNA prepared as described above, PCR was performed with ExTaq DNA polymerase (Takara). The primers used were 5′-TAATACGACTCACTATAGGGTCATAATGAGTTGAGTTCT-3′ (forward) and 5′-TAATACGACTCACTATAGGGAAG GGGTGTCTGTAATCTT-3′ (reverse) for Gb’Clk and 5′-TAATACGACTCACTATAGGGATGTGGCTTGGAAGATCATT-3′ (forward) and 5′-AATTAACCCTCACTAAAGGGTCTCCTTAAGCAAATTCTCA-3′ (reverse) for Gb’per. Amplified Gb’Clk (622 bp) and Gb’per (805 bp) fragments were linearized with phenol/chloroform and precipitated with ethanol. For DsRed2 dsRNA, DsRed2 was linearized from pDsRed2-N1 (Clontech, Mountain View, CA) and amplified with the forward and reverse primers. With each of these linearized fragments as a template, RNA was synthesized with T7 or T3 RNA polymerase. Synthesized RNA was extracted with phenol/chloroform and suspended in 50 µL of TE solution after isopropanol precipitation. The yield and quality of RNA were assessed by absorbance with a spectral photometer (GeneQuant Pro, Amersham Biosciences, Piscataway, NJ), and the same amount of sense and antisense RNA were mixed. The RNA was denatured for 5 minutes at 100 °C and annealed by a gradual cool down to room temperature. After ethanol precipitation, the obtained dsRNA was suspended in Ultra Pure Water (Invitrogen) and adjusted to the final concentration of 20 µM. The dsRNA solution was stored at −80 °C until use. There was 760 nL of dsRNA solution injected with the nanoliter injector (WPI, Sarasota, FL) into the abdomen of eighth instar nymphs or adults anesthetized with CO2.
Behavioral Analysis
Locomotor activities were recorded in the same way as described previously (Moriyama et al., 2008). Briefly, the final instar nymphs or adult crickets were individually housed in a transparent plastic box (18 × 9 × 4.5 cm) with a rocking substratum. The number of times the substratum rocked backwards and forwards was recorded every 6-minute period by a computerized system. Food and water were provided ad libitum. The activity was monitored in an incubator (MIR-153, Sanyo Biomedica, Osaka, Japan) in which the temperature was kept at 25 °C, and lighting conditions were given by a cool white fluorescent lamp connected to an electric timer. The light intensity was 600 to 1000 lux at the animal’s level, varying with proximity to the lamp. The raw data were displayed as conventional double-plotted actograms to judge activity patterns and statistically analyzed by a χ2 periodogram (Sokolove and Bushell, 1978) with ActogramJ (freely available at http://actogramj.neurofly.de/) (Schmid et al., 2011). If a peak of the periodogram appeared above the 0.05 confidence level (α = 0.005), the power value (height of the peak above the confidence level) was greater than or equal to 10, and the width of the peak was greater than or equal to 2; the period for the peak was designated as statistically significant (Kaneko et al., 2000).
Results
Cloning and Sequence of the Clock Gene
Cloning of a Clk homolog was performed with cDNA from the optic lobe of G. bimaculatus by a degenerate PCR strategy with degenerate primers based on conserved amino acid sequences from vertebrate and insect Clk homologs. A cDNA fragment of about 1000 bp including the PAS domain was first obtained. Then, 3′ and 5′ RACE were performed with gene-specific primers designed from the obtained sequence. The obtained full-length cDNA of 3325 bp encodes a protein of 594 amino acid residues with 281 and 1262 bp of 3′- and 5′-UTR, respectively.
A BLAST database search indicated that the amino acid sequence has 65% to 79% of identities along the entire length of the protein with those of known insects’ Clks, including the firebrat Thermobia domestica, the honey bee Apis mellifera, the beetle Tribolium castaneum, the silkmoth Antheraea pernyi, the louse Pediculus humanus corporis, and the fruit fly D. melanogaster (Suppl. Table S1). The deduced protein had 5 highly conserved regions that are characteristic in known CLK proteins (Fig. 1A and 1B): 1) an amino terminal region that includes a bHLH domain; 2) a PAS-A domain; 3) a PAS-B domain; 4) a PAC domain, a region immediately carboxy terminal to the PAS-B domain, which was shown to function as a cytoplasmic localization determinant in D. melanogaster PER (Saez and Young, 1996); and 5) polyglutamine repeats (Poly-Q) in C-termini. The PAC domain had a particularly high identity with those of other insect Clk homologs (Fig. 1A), suggesting its conserved functional role. The glutamine enrichment of C-termini is thought to be indicative of their transactivational function (King et al., 1997; Allada et al., 1998; Darlington et al., 1998). Based on these identities, we concluded that this protein represents a cricket’s CLK (Gb’CLK; GenBank/EMBL/DDBJ accession no. AB738083). A phylogenetic tree based on the amino acid sequences of CLKs from known insect species revealed a close relationship between the cricket and the firebrat (Fig. 1C).
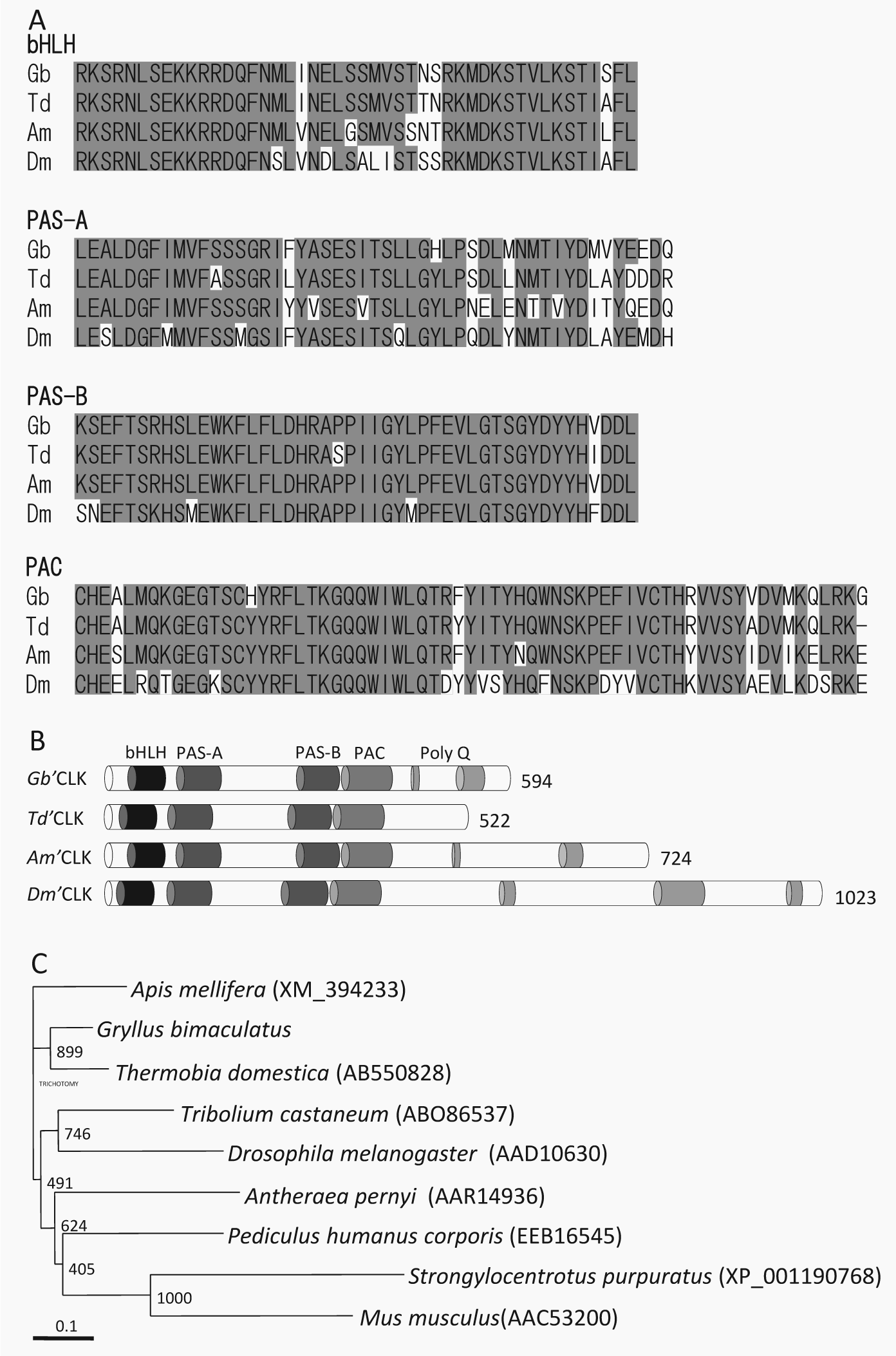
Sequence alignments of conserved domains of Gb’Clk compared with those orthologous sequences from other insects. (A) bHLH, PAS-A, PAS-B, and PAC of Gryllus bimaculatus Clock (Gb) aligned with that of Thermobia domestica (Td), Apis mellifera (Am), and Drosophila melanogaster (Dm). The amino acid residues showing more than 60% of identity are shaded. (B) Schematic structure of various CLK proteins, comparing the organization of the 5 conserved domains: bHLH, PAS-A, PAS-B, PAC, and Poly-Q. (C) A phylogenetic neighbor-joining tree of known insect CLK proteins. CLK amino acid sequences were analyzed, and the neighbor-joining tree was inferred with ClustalW. The GenBank or RefSeq accession numbers are indicated in brackets. A reference bar indicates the distance as number of amino acid substitutions per site.
Gb’Clk Transcript Expression in the Optic Lobe under LD and DD
To determine if Gb’Clk transcripts oscillated in a circadian manner, the levels of Gb’Clk mRNA in the optic lobe were examined under LD 12:12 and in the second day after transfer to constant darkness (DD) by qPCR (Fig. 2). The Gb’Clk mRNA level showed a slight fluctuation but no significant rhythmic profiles both under LD and DD (analysis of variance [ANOVA], p > 0.1).
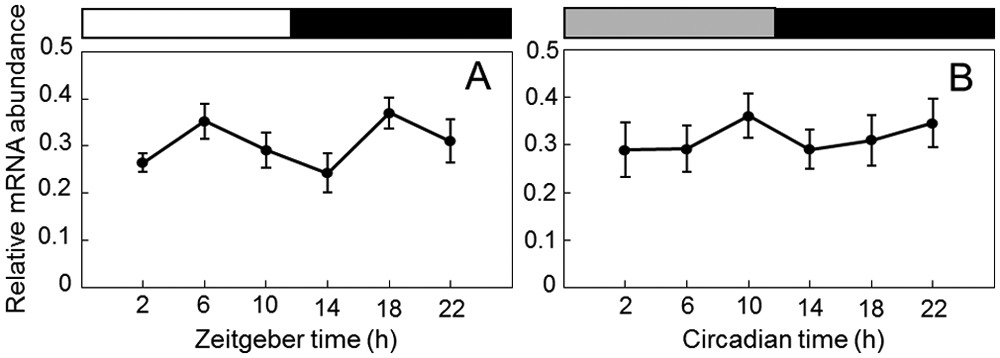
Expression profiles of Clk mRNA in Gryllus bimaculatus optic lobes under LD 12:12 (A) and DD (B). The abundance of Gb’Clk mRNA was measured by quantitative real-time RT-PCR. Total RNA was extracted from the optic lobes that were collected at 4-hour intervals starting at 2 hours after lights-on (ZT 2) or circadian time (CT) 2 in the second day of DD. White, black, and gray bars indicate light phase, night/subjective night, and subjective day, respectively. The data collected from 8 independent experiments were averaged and plotted as mean ± SEM. The abundance of rpl18a mRNA was used as an internal reference. Note that no significant rhythm was found both in LD and DD.
Gb’Clk dsRNA Suppresses Levels of Gb’Clk Transcripts
To examine whether Gb’Clk dsRNA suppressed the Gb’Clk transcript levels, the Gb’Clk mRNA levels were measured by qPCR in the optic lobe of the adult male crickets treated with Gb’Clk dsRNA in the final instar nymphal stage (Fig. 3). The optic lobes were collected more than 7 days after the imaginal molt. As a control, we measured Gb’Clk mRNA levels in crickets treated with DsRed2 dsRNA, and the levels in LD were almost equivalent to those of intact crickets (Fig. 3). In the treated crickets, however, the Gb’Clk mRNA level was significantly reduced to nearly 25% of the peak level of intact crickets at ZT 18 (t test, p < 0.01), and this level was maintained throughout the day, showing no rhythmic changes (ANOVA, p > 0.1). Thus, Gb’Clk dsRNA successfully suppressed the expression of Gb’Clk mRNA through RNAi.
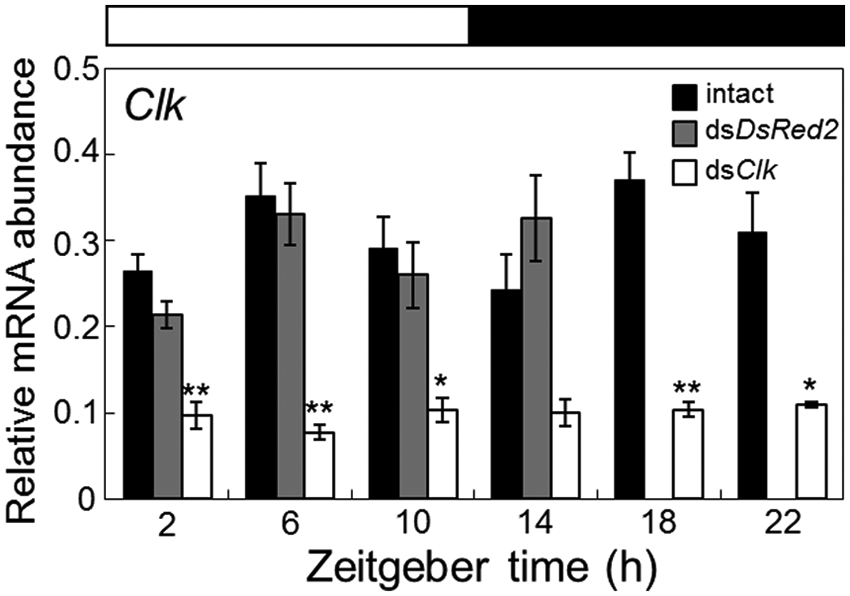
Effects of Clk and DsRed2 dsRNA on the Clk mRNA levels in the optic lobe of adult male crickets (Gryllus bimaculatus). Black, gray, and white columns indicate the mRNA levels of intact, DsRed2, and Clk dsRNA-injected crickets, respectively. The data for intact Clk mRNA are replotted from Figure 2A. In dsRNA-injected crickets, the optic lobes were collected more than 7 days after injection. The abundance of mRNA was measured by quantitative real-time RT-PCR with total RNA extracted from the optic lobes. The data collected from 3 and 8 independent experiments were averaged and plotted as mean ± SEM for dsRNA-injected and intact crickets, respectively. The abundance of rpl18a mRNA was used as an internal reference. Note that Clk dsRNA significantly reduced Clk mRNA levels, while DsRed2 dsRNA had no significant effect. *p < 0.05, **p < 0.01, t test.
Loss of Locomotor Rhythm Induced by Gb’Clk RNAi
To examine the effects of Gb’Clk dsRNA on the locomotor rhythm, locomotor activity was recorded in 20 males injected with Gb’Clk dsRNA into the abdomen at the eighth instar nymphal stage 1 day before the start of recording. Seventy percent of the treated crickets exhibited an activity rhythm under LD, with peaks at lights-on and lights-off as exemplified in Figure 4A, and the remaining 30% were arrhythmic (Table 1). An actogram of the cricket that showed a rhythm under LD is shown in Figure 4A. This particular animal exhibited a weak rhythm with small peaks at lights-on and lights-off and another weak, somewhat dispersed, peak in the middle of the day during the first several days, a pattern characteristic to nymphal crickets (Tomioka and Chiba, 1982). The nymphal activity pattern changed to an adult locomotor pattern with a burst of nocturnal activity at lights-off. In the ensuing DD, however, the nocturnal rhythm disappeared immediately, suggesting that it is a masking effect of light. Likewise, on transfer to DD as adults, more than 80% of the 21 crickets immediately became arrhythmic with activity dispersed over the entire day. The remaining 4 crickets showed a free-running rhythm with a period of 24.1 ± 0.1 hours (mean ± SD) but with a significantly weaker power (Table 1). Six of the arrhythmic crickets were exposed to LD for 12 days after 34 days of DD. They showed an intense burst of activity persisting for a few hours after lights-off, while on transfer to DD, they immediately began to show an arrhythmic activity pattern again, which persisted until the end of the record (Fig. 4A), confirming that the nocturnal activity in LD is a masking effect.
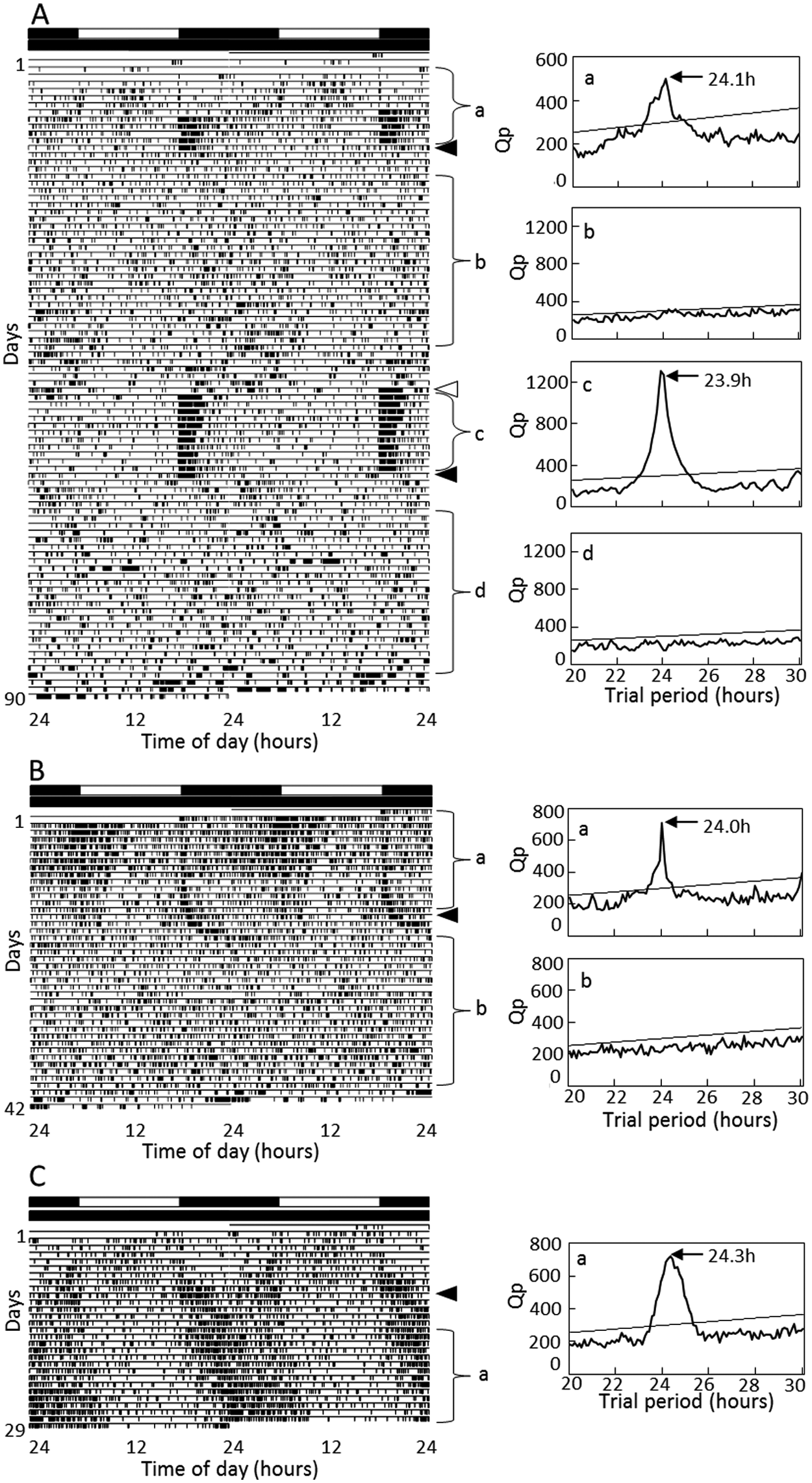
Double-plotted actograms (left) and χ2 periodograms (right) of locomotor rhythms of crickets Gryllus bimaculatus injected with Gb’Clk dsRNA at the nymphal (A) or adult stage (B) or with DsRed2 dsRNA (C) under LD 12:12 and DD at a constant temperature of 25 °C. Filled arrowheads indicate the day when the crickets were transferred from LD to DD and an open arrowhead from DD to LD. a~d indicated in the periodogram correspond to the analyzed time span, a~d, indicated in the actogram. An oblique line in the periodogram indicates a significance level of p < 0.01; a peak value above the line was designated as significant. The cricket showed arrhythmic locomotor activity in DD (A, B), but the insect in A showed a clear rhythm in LD, which disappeared on transfer to DD. The control cricket injected with DsRed2 dsRNA showed a clear rhythm throughout the recording (C). For further explanations, see the text.
Effects of dsClk and dsDsRed2 on the locomotor rhythm of the cricket Gryllus bimaculatus.
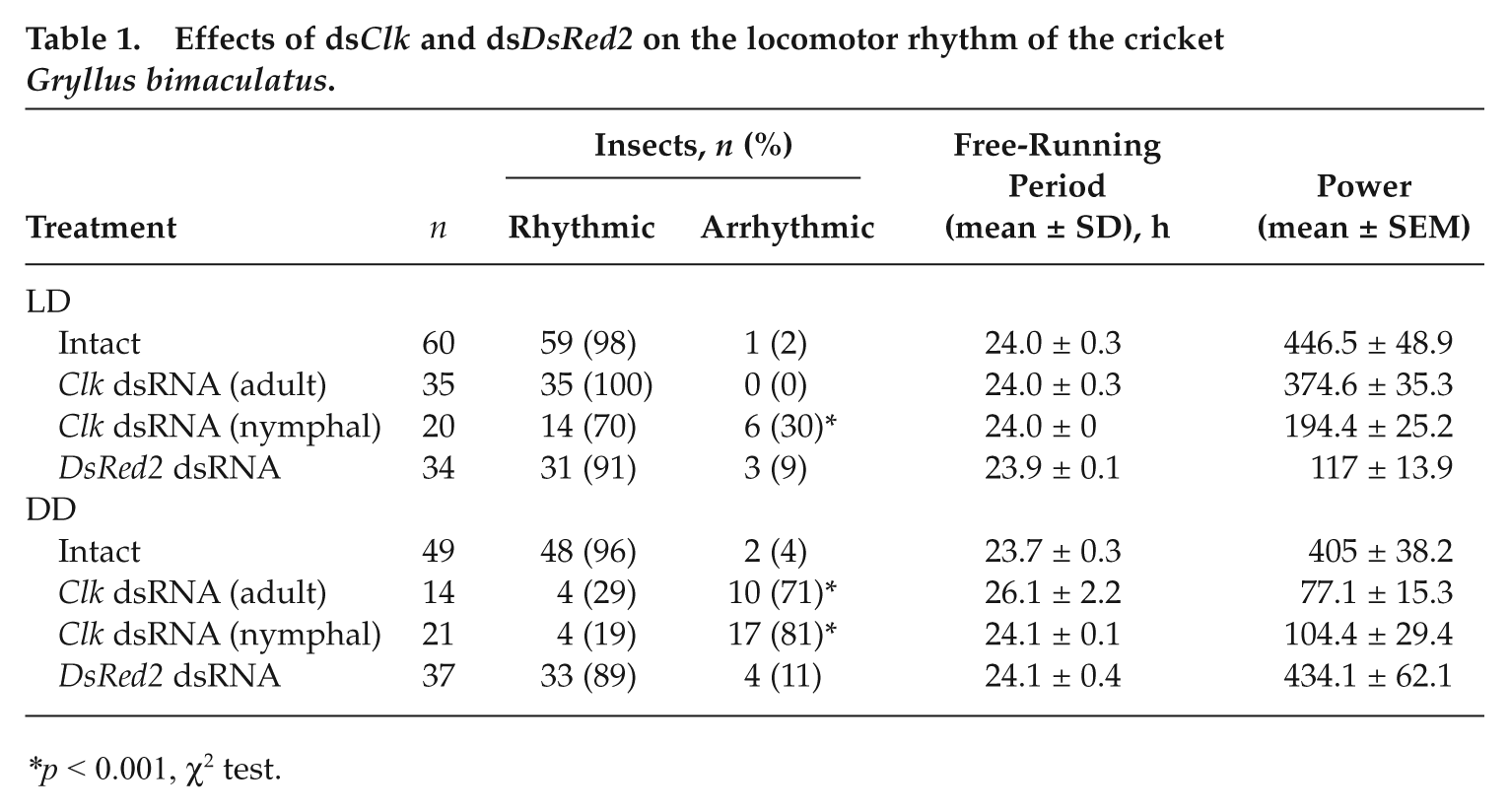
p < 0.001, χ2 test.
In addition, to rule out the possibility that Gb’Clk dsRNA affected the development and induced arrhythmicity in adults, Gb’Clk dsRNA was injected into 35 adult crickets within 3 days after imaginal molt, and their locomotor rhythms were examined. In LD, the crickets showed a clear daily rhythm with peaks at lights-off and around lights-on (Fig. 4B). On transfer to DD, although less than 30% of the Gb’Clk RNAi crickets still maintained the free-running rhythm with a significantly longer period of 26.1 ± 2.2 hours (mean ± SD) and with a weaker power, the remaining more than 70% of the crickets became arrhythmic often with a few residual cycles (Fig. 4B and Table 1), suggesting that the effects of Gb’Clk dsRNA are not developmental.
As a negative control, DsRed2 dsRNA was injected into 37 final instar nymphs, and its effects on the locomotor rhythm were investigated in the adult stage. Figure 4C is an example of the record. This particular animal showed a change in locomotor activity from nymphal weak diurnality to adult nocturnality during the first several days, and the nocturnal rhythm persisted in the ensuing DD. Likewise, most of the control crickets showed a similar nocturnal locomotor rhythm, which free-ran with an average period of 24.1 ± 0.4 hours (n = 33) (Table 1).
Gb’Clk dsRNA Suppresses Levels of Gb’per and Gb’tim Transcripts
In Drosophila, CLK is known to form a heterodimer with CYC to activate the transcription of per and tim (Tomioka and Matsumoto, 2010). To examine whether Gb’Clk is involved in the regulation of these genes, the levels of Gb’per and Gb’tim mRNA in the optic lobe were measured by qPCR in adult male crickets injected with Gb’Clk dsRNA in the final instar nymphal stage. The optic lobes were collected more than 7 days after the imaginal molt. In the treated crickets, the Gb’per and Gb’tim mRNA levels were both reduced to nearly 30% of the peak level at ZT 18, respectively (t test, p < 0.05), and their rhythmic expression was completely abolished (ANOVA, p > 0.05) (Fig. 5A and 5B). These results suggest the involvement of Gb’CLK in the transcriptional regulation of Gb’per and Gb’tim.
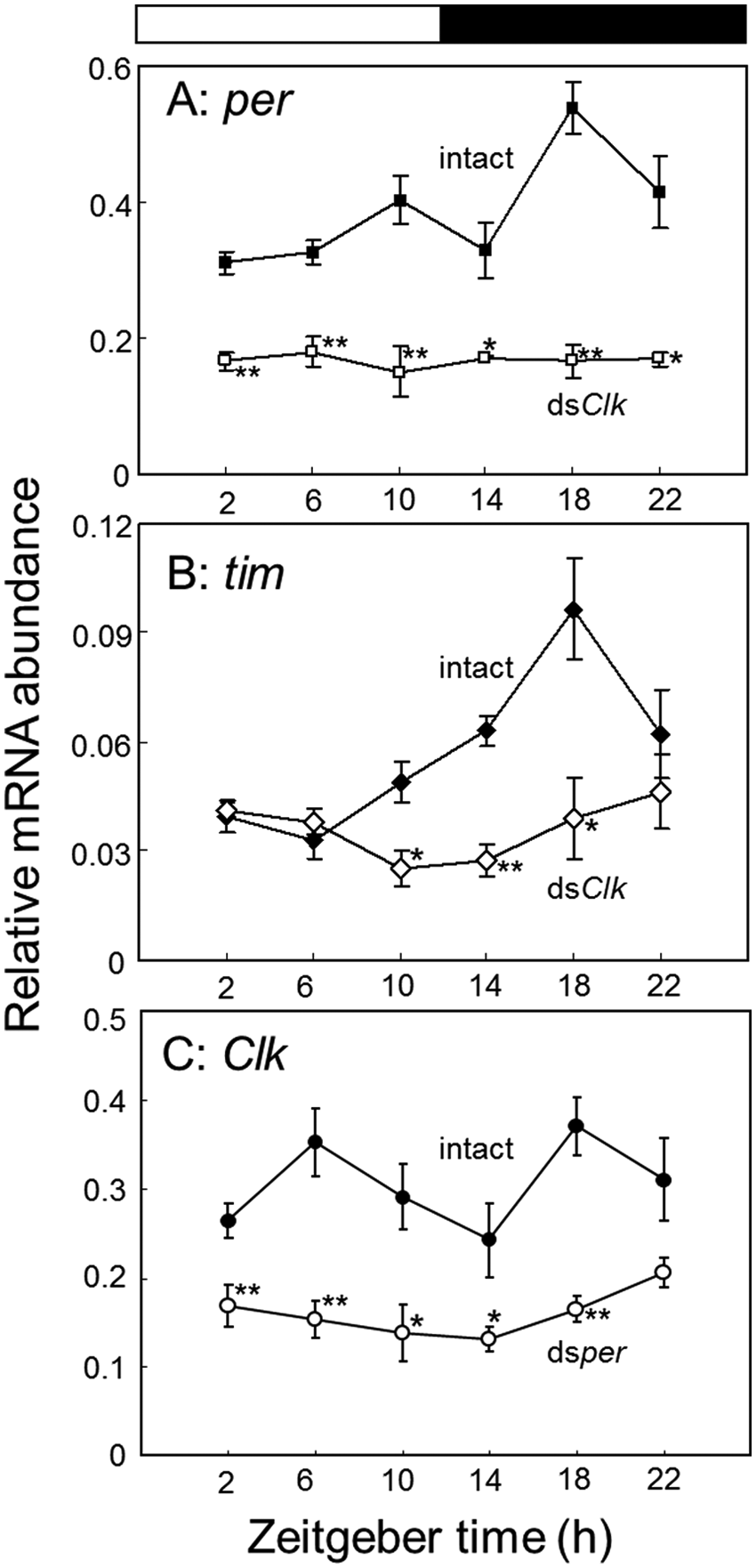
Relative abundance of per (A), tim (B), or Clk mRNA (C) in the optic lobes of intact and Clk dsRNA-injected (A, B) or per dsRNA-injected adult male crickets (Gryllus bimaculatus) (C). The data for intact Clk mRNA are replotted from Figure 2A. Filled and open symbols indicate the mRNA levels of intact and dsRNA-injected crickets, respectively. In dsRNA-injected crickets, the optic lobes were collected more than 7 days after the dsRNA injection. The abundance of mRNA was measured by quantitative real-time RT-PCR with total RNA extracted from the optic lobes. The data collected from 3 and 8 independent experiments were averaged and plotted as mean ± SEM for dsRNA-injected and intact crickets, respectively. The abundance of rpl18a mRNA was used as an internal reference. *p < 0.05, **p < 0.01, t test. Note that Clk dsRNA down-regulates both per and tim mRNA and prevents their rhythmic expression, and per dsRNA reduced the Clk mRNA levels. For further explanations, see the text.
Effects of Dsper on Gb’Clk
In Drosophila, transcription of Clk is hypothesized to be regulated by PDP1ε and VRILLE (VRI), whose transcription is in turn regulated by CLK/CYC. Since transcriptional activity of CLK/CYC is repressed by PER/TIM (Bae et al., 1998), transcription of Clk is indirectly affected by PER/TIM. To test this possibility in the cricket, we measured levels of Gb’Clk mRNA in crickets injected with Gb’per dsRNA under LD. The knockdown of Gb’per by its dsRNA has been confirmed previously (Moriyama et al., 2008). In the treated crickets, Gb’Clk mRNA levels stayed below the basal level of intact animals throughout the day, and no significant rhythm was observed (ANOVA, p > 0.05); the reduction was statistically significant at all the time points except for ZT 22 (t test, p < 0.05) (Fig. 5C).
Discussion
Role of Clock Gene in the Cricket
In the Drosophila circadian clock, at least 3 interdependent feedback loops are involved, and Clk participates as part of the transcriptional activator (positive element) in all 3 loops, regulating the transcription of per, tim, vri, Pdp1ε, and clockwork orange (Tomioka and Matsumoto, 2010). Clk regulates many other clock-controlled genes together with cyc, and its ectopic expression can induce rhythmic expression of clock genes in nonclock cells (McDonald and Rosbash, 2001; Zhao et al., 2003; Kilman and Allada, 2009). It is thus one of the core elements of the clock. The results of our present study clearly indicate that Clk is an essential element of the circadian clock also in the cricket G. bimaculatus.
Our results show that knockdown of Gb’Clk resulted in a loss of rhythms both at behavioral and molecular levels (Figs. 3-5). Gb’Clk RNAi suppresses the mRNA levels of both Gb’per and Gb’tim. This is in good agreement with what is expected from the Drosophila clock model. Together with the previous findings that Gb’per and Gb’tim mRNA show rhythmic expression (Moriyama et al., 2008; Danbara et al., 2010), it is most probable that Gb’CLK acts as a positive element in the clock machinery, transactivating Gb’per and Gb’tim like Drosophila CLK does (Allada et al., 1998). To test this hypothesis, the transcriptional activity of Gb’CLK should be examined.
The Clk gene has been cloned or reported in various insect species, and its function has been studied in the silkmoth and the monarch butterfly using culture cells (Allada et al., 1998; Chang et al., 2003; Rubin et al., 2006; Zhu et al., 2008; Gentile et al., 2009; Kamae et al., 2010; Rund et al., 2011). However, besides Drosophila Clk, in vivo functional analysis of Clk has been performed only in the mosquito Anopheles gambiae (Das and Dimophoulos, 2008) and the firebrat (Kamae et al., 2010). In the former species, its involvement in the clock machinery is only speculated based on the fact that its RNAi-based silencing increased the circadian-based blood-feeding behavior (Das and Dimophoulos, 2008). In the latter species, knockdown of Clk mRNA also resulted in a reduced level of tim mRNA, which led to a loss of locomotor rhythm (Kamae and Tomioka, 2012), as we found here in the cricket. Thus, the role of Clk seems quite consistent across a wide range of insect species from apterygotes to higher pterygotes.
Apart from the clock, it was recently found in the desert locust, Schistocerca gregaria, that nymphal treatment with Clk dsRNA delayed adult emergence or resulted in a high rate of lethality, and Clk was suggested to have a role in postembryonic development (Tobback et al., 2012). However, we have observed no such developmental failures in both crickets and firebrats (Kamae et al., 2010). Thus, the developmental role of Clk seems species dependent.
Regulation of Clk Expression
Interestingly, Gb’Clk showed no significant rhythmic expression in the optic lobe in both LD and DD conditions. This contrasts with the fact that Drosophila Clk mRNA oscillates in both LD and DD with a peak near dawn under a mechanism including vri and Pdp1ε (Bae et al., 1998; Cyran et al., 2003; Glossop et al., 2003), suggesting that the transcription of Gb’Clk may be regulated through a pathway without vri and Pdp1ε or a mechanism in which vri and Pdp1ε play roles different from those in Drosophila. The arrhythmic expression of Clk is consistent with results from the honey bee and the firebrat (Rubin et al., 2006; Kamae et al., 2010) and also similar to that in mammalian species (King et al., 1997). In this respect, the clock of these insects has a molecular mechanism that is more similar to a mammalian one than that of Drosophila (Dunlap, 1999). One may argue that Gb’Clk is rhythmically expressed in a subset of neurons that controls behavior but that this is masked by constitutive expression in other nonpacemaker cells in the optic lobe. However, this seems unlikely since other clock genes such as Gb’per and Gb’tim show robust rhythms of their transcripts in the optic lobe.
A question arising here is how the rhythmic per and tim expressions are regulated without rhythmic expression of Clk. Interestingly, in the firebrat, cyc is rhythmically expressed to regulate the rhythmic transcription (Kamae et al., 2010). Whether a similar system is operated in the cricket’s clock should be examined in future studies. Alternatively, however, constitutive Gb’Clk expression might be sufficient to drive the rhythm since it has been reported in Drosophila that the CLK protein is constitutively expressed in clock neurons (Houl et al., 2006; Yu et al., 2006) and that constitutive, ectopic expression of Drosophila Clk induces rhythmic expression of per, tim, and cry (Zhao et al., 2003). This view is also consistent with the report by Kim et al. (2002) that Clk mRNA rhythms were unimportant for circadian behavioral rhythms.
Interestingly, our data indicate that Gb’per is required for the sufficient expression of Gb’Clk at least in LD (Fig. 5C) like in Drosophila (Bae et al., 1998). It is difficult to give a consistent explanation for this fact, but one may explain according to the Drosophila clock model. Since CLK/CYC transcriptional activity is suppressed by PER/TIM, a reduced level of PER/TIM may increase the transcriptional activity of CLK/CYC. The enhancement results in an increased level of VRI, which may reduce the Clk transcript level. However, this leaves a question of then why the transcriptional activity of PDP1ε is not observed, which might also increase along with VRI. Alternatively, there are other possibilities that the PER/TIM complex shuttles a transcriptional activator into the nucleus, or it acts as a negative regulator of the transcriptional repressor for Gb’Clk as has been suggested in Drosophila (Bae et al., 1998).
Comparison of Cricket’s Clock with Other Insects
The cricket’s clock shares at least 3 clock genes, per, tim, and Clk, with the Drosophila clock but differs in some aspects: Gb’Clk has structural characteristics common to Dm’Clk, but its daily expression profile is different from Dm’Clk. In addition, Gb’tim seems to be not an essential component but may have an important role: a reduced level of its mRNA by RNAi shortens the free-running period (Danbara et al., 2010). The constitutive Clk expression shared with the more ancestral apterygote insect T. domestica. Thus, the cricket’s clock retains an ancestral trait in this aspect, which later may have changed to the Drosophila type in higher insects. However, there is a considerable difference in tim’s role even between these 2 species (Danbara et al., 2010; Kamae and Tomioka, 2012). The role of tim seems greatly diversified among insects with an extreme case in hymenopteran insects where the tim gene has been lost from their genome (Rubin et al., 2006; Zhan et al., 2011).
At the present stage, it is thus still premature to figure out the phylogeny of insect clock mechanisms. More detailed information is necessary on the clock genes and their roles in a variety of insect species to understand the diversification and its biological significance. Research on the cricket clock mechanism would contribute to the comparative approach to the insect circadian clock because the molecular dissection of the clock could be done quite efficiently (Tomioka et al., 2009).
Footnotes
Acknowledgements
The authors thank Drs. Akira Matsumoto and Taishi Yoshii for critically reading an earlier version of the article. This study was supported in part by grants from the Japan Society for Promotion of Science (JSPS) to K.T. Y.M. was a JSPS postdoctoral research fellow, and Y.K. and O.U. are JSPS research fellows.
Conflict of Interest Statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article. [This has been confirmed.]
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.