
Abstract
Circadian rhythms represent an adaptive feature, ubiquitously found in nature, which grants living beings the ability to anticipate daily variations in their environment. They have been found in a multitude of organisms, ranging from bacteria to fungi, plants, and animals. Circadian rhythms are generated by endogenous clocks that can be entrained daily by environmental cycles such as light and temperature. The molecular machinery of circadian clocks includes a transcriptional-translational feedback loop that takes approximately 24 h to complete. Drosophila melanogaster has been a model organism of choice to understand the molecular basis of circadian clocks. However, alternative animal models are also being adopted, each offering their respective experimental advantages. The nematode Caenorhabditis elegans provides an excellent model for genetics and neuro-behavioral studies, which thanks to its ease of use and manipulation, as well as availability of genetic data and mutant strains, is currently used as a novel model for circadian research. Here, we aim to evaluate C. elegans as a model for chronobiological studies, focusing on its strengths and weaknesses while reviewing the available literature. Possible zeitgebers (including light and temperature) are also discussed. Determining the molecular bases and the neural circuitry involved in the central pacemaker of the C. elegans’ clock will contribute to the understanding of its circadian system, becoming a novel model organism for the study of diseases due to alterations of the circadian cycle.
The rotation of the Earth causes predictable changes in several variables, such as light and temperature, in our natural environment. Accordingly, natural selection has favored the evolution of biological circadian (from the Latin circa, meaning “about,” and dies, meaning “day”) clocks. Such clocks are autonomous and generate an about-24 h rhythm that needs to be entrained to the environment in order to adapt to external daily changes of light, temperature, and other environmental factors. Circadian rhythms have some common basic characteristics: they persist for long periods in the absence of environmental signals, their period is temperature-compensated maintaining a similar value over a wide range of temperatures, and they synchronize to daily environmental cycles, using light and temperature as major entrainment agents or zeitgebers. Most animals have a hierarchical network of central and peripheral clocks. In both vertebrates and invertebrates, a group of neurons control circadian behaviors and can convey information about the time-of-day to peripheral clocks located in different organs and tissues. (Ahmad et al., 2021; Hastings et al., 2018; Helfrich-Forster et al., 1998; Patke et al., 2020). The circadian clock is genetically controlled, and mutations in so-called clock genes can change rhythmic behavior in animals, plants, fungi, and bacteria (Patke et al., 2020; Young and Kay, 2001).
C. elegans is a model organism used in different fields of research including neurobiology, developmental biology, and genetics. Different circadian rhythms have been described in C. elegans ranging from behavior to gene expression, including locomotor activity, abiotic and biotic stress tolerance, olfaction, and metabolism (Figure 1). In addition, bioinformatic analysis has identified C. elegans proteins of relatively high homology to clock components from other species (Romanowski et al., 2014). However, most of the studies have consisted mainly of a description of circadian outputs, applying classical in vitro methods or behavioral recordings, but the molecular components of its central clock are currently unknown. This review discusses the experimental approaches and findings related to C. elegans circadian rhythms, lists the nematode proteins with relatively high identity to the mammalian/fly core clock genes, and discusses possible synchronizing signals such as light and temperature.
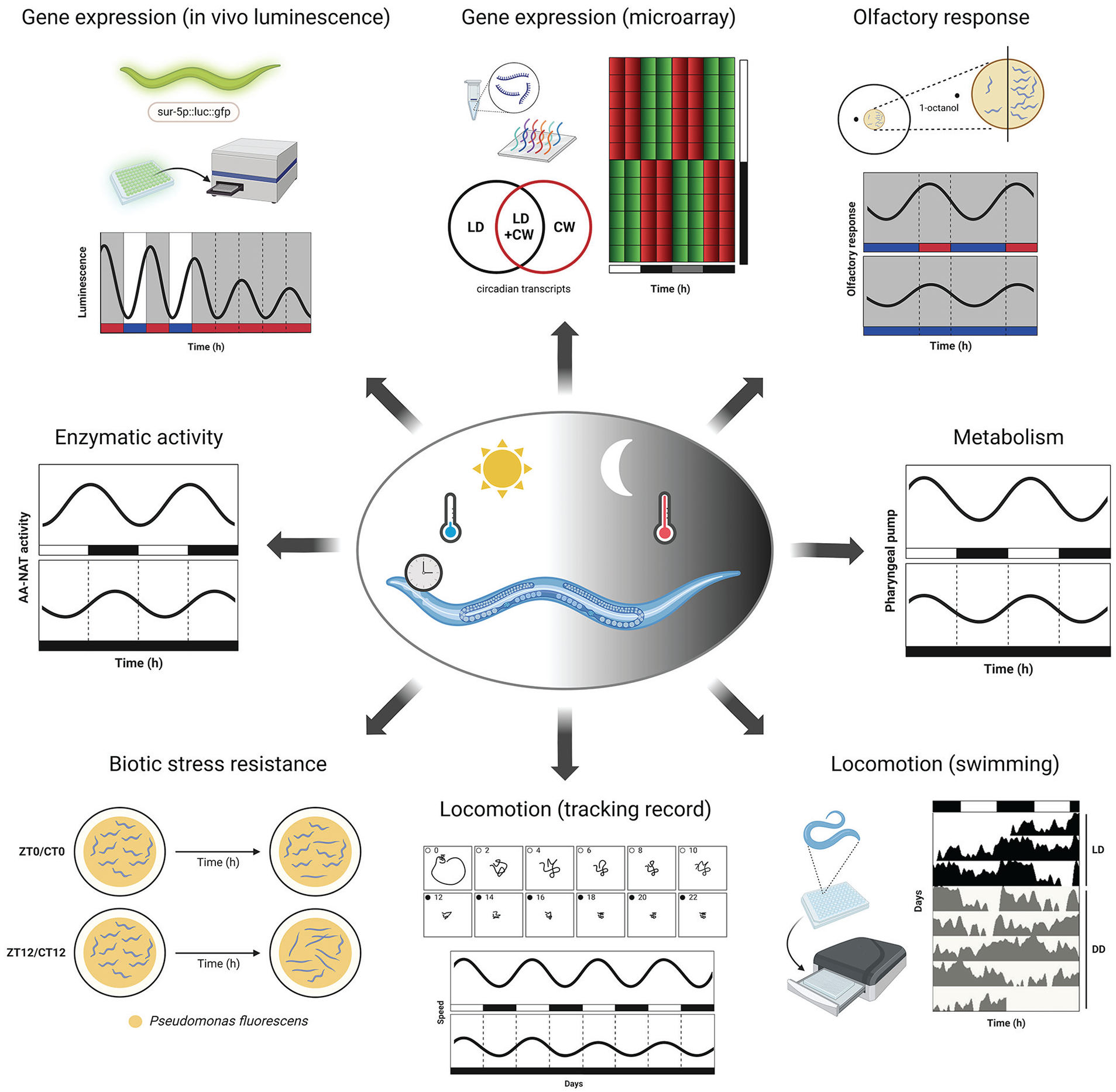
Circadian rhythms characterized in C. elegans. Numerous circadian cycles have been described in the nematode at different levels, including locomotory behavior (Saigusa et al., 2002; Simonetta and Golombek, 2007; Simonetta et al., 2009; Winbush et al., 2015), olfaction (Olmedo et al., 2012), metabolism (abiotic and biotic stress tolerance, food and oxygen consumption, pharyngeal pumping rate) (Migliori et al., 2011; Romanowski et al., 2011; Simonetta et al., 2008), protein activity and regulation (Migliori et al., 2012; Olmedo et al., 2012), and gene expression (Goya et al., 2016; van der Linden et al., 2010). Black/white bars indicate dark/light periods; blue/red bars indicate cold/warm periods. Abbreviations: LD = light-dark; CW = cold-warm; CT = circadian time; ZT = zeitgeber time; DD = dark conditions.
Advantages of C. elegans as Model Organism
C. elegans is a powerful and attractive model for the study of chronobiology because it is an organism with a completely sequenced genome (C. elegans Sequencing Consortium, 1998), a compact and simple nervous system (302 neurons), and a fully known neuronal connectome (Bhattacharya et al., 2019; Flavell and Gordus, 2022). It is also especially amenable to genetic modifications by classical mutagenesis, RNA interference (RNAi), and CRISPR. Furthermore, several aspects of its basic biology, such as its small size (adults are approximately 1 mm in length), short lifespan (15-20 days at 20 °C), ability to self-fertilize and high reproductive rate (~300 offspring per hermaphrodite), and its transparency, make it an ideal model for behavioral and molecular genetics studies. Moreover, nematodes show different and easily measurable patterns of behavior, including locomotor activity, feeding, and defecation (Avery and Horvitz, 1989; Simonetta et al., 2009; Thomas, 1990). Another major advantage of these nematodes is the possibility to study the function of specific neurons and neuronal circuits by programmed cell death induced by the specific expression of human caspases (Chelur and Chalfie, 2007), or to control neuronal activity using the histamine-gated chloride channel HisCl1 from Drosophila and exogenous histamine (Pokala et al., 2014). In addition, C. elegans is an exceptional model organism for biomedical research because roughly 30%-60% of genes in nematodes have orthologs or strong homologs in mammals (Apfeld and Alper, 2018; Shaye and Greenwald, 2011). In this sense, determining the molecular bases and the role of specific neurons in the biological clock of C. elegans will contribute to the understanding of general principles of circadian biology, and will also enable it to be established as a model organism for the study of diseases associated with alterations of the circadian system.
Chronobiology and Sleep in C. elegans
Biological rhythms were originally being characterized in C. elegans based on the study of the ultradian defecation clock (i.e., period less than 24 h) (Branicky and Hekimi, 2006; Branicky et al., 2001; Iwasaki et al., 1995). However, in 2002, two independent studies provided the first descriptions of circadian rhythms in C. elegans, demonstrating that the nematode exhibited circadian rhythms in behaviors such as locomotion or responses to osmotic stress in L1 larvae (Kippert et al., 2002; Saigusa et al., 2002). One of the reasons it took so long to study circadian rhythms in these animals was the lack of robust and reliable methods to measure daily rhythms. In this sense, our laboratory first characterized circadian rhythms of locomotion in adult nematodes—single worms or populations—(Caldart et al., 2020; Simonetta and Golombek, 2007; Simonetta et al., 2009) through the development of a locomotor activity recording system (Wmicrotracker, Phylum Technologies, Argentina). Winbush et al. (2015), used an imaging-based system for long-term automated recording and analysis of locomotion data of multiple nematodes on the surface of an agar plate (Winbush et al., 2015). Moreover, through the development of a bioluminescent reporter-based method, our group was able to demonstrate circadian rhythms of gene expression under rhythmic and constant environmental conditions (Goya et al., 2016). Furthermore, daily rhythms in pharyngeal pumping (Migliori et al., 2011), in the production of the hormone melatonin (Migliori et al., 2012) and in resistance to different types of abiotic and biotic stress (Romanowski et al., 2011; Simonetta et al., 2008) have been characterized in C. elegans.
Although long-term recording methods allow obtaining reliable data and, in some cases, even following rhythms from single-worm, results of entrainment under light and/or temperature cycles and free-running suggests that the circadian system of C. elegans exhibits properties of a weak oscillator, characterized by significant masking and damping effects under free running, at least under laboratory conditions assayed so far. Population experiments often show a low proportion of real entrainment to the external inputs, under both light and temperature cycles (Goya et al., 2016; Simonetta et al., 2009). One of the possible reasons for this is that the maybe synchronization conditions or zeitgebers used in the lab aren’t the optimal for the entrainment of the worm’s clock, and they are still far from the ones sensed in their natural niche. This is specifically critical in population assays, where most individuals need to be synchronized relative to one another and to the external output to discern daily oscillations. The lack of entrainment of most of the individuals in population assays could also contribute to the quick damping seen in free running, as a result of an averaging effect.
On the other hand, the C. elegans reference strain commonly used in laboratories corresponds to the N2 strain (wild-type), which was obtained from mushroom compost in Bristol, England, and later provided by Ellsworth Dougherty to Sydney Brenner in 1964 (Riddle et al., 1997). This strain was grown for many years under constant temperature on agar plates with bacteria until it was frozen for the first time, so it is considered a domesticated strain. Different studies show that recent isolates of C. elegans are highly divergent at the genomic level with respect to the N2 strain (Vergara et al., 2014; Weber et al., 2010) due to the accumulation of numerous mutations that also generate phenotypic differences. In this sense, it would be interesting to study circadian rhythms in wild isolates of C. elegans to analyze possible changes in the pattern of oscillations and to see whether the variability and lack of robustness observed in some rhythmic behaviors of the laboratory strain are related to genetic variations accumulated after domestication. Although in C. elegans (N2 strain) circadian outputs do not appear to be robust and there is large inter-individual variability (Herrero et al., 2015; Kippert et al., 2002; Simonetta and Golombek, 2007; Winbush et al., 2015), the basic circadian properties have been studied and identified in these nematodes by different research groups (Table 1).
Circadian properties and entrainment protocols used in our and other studies.
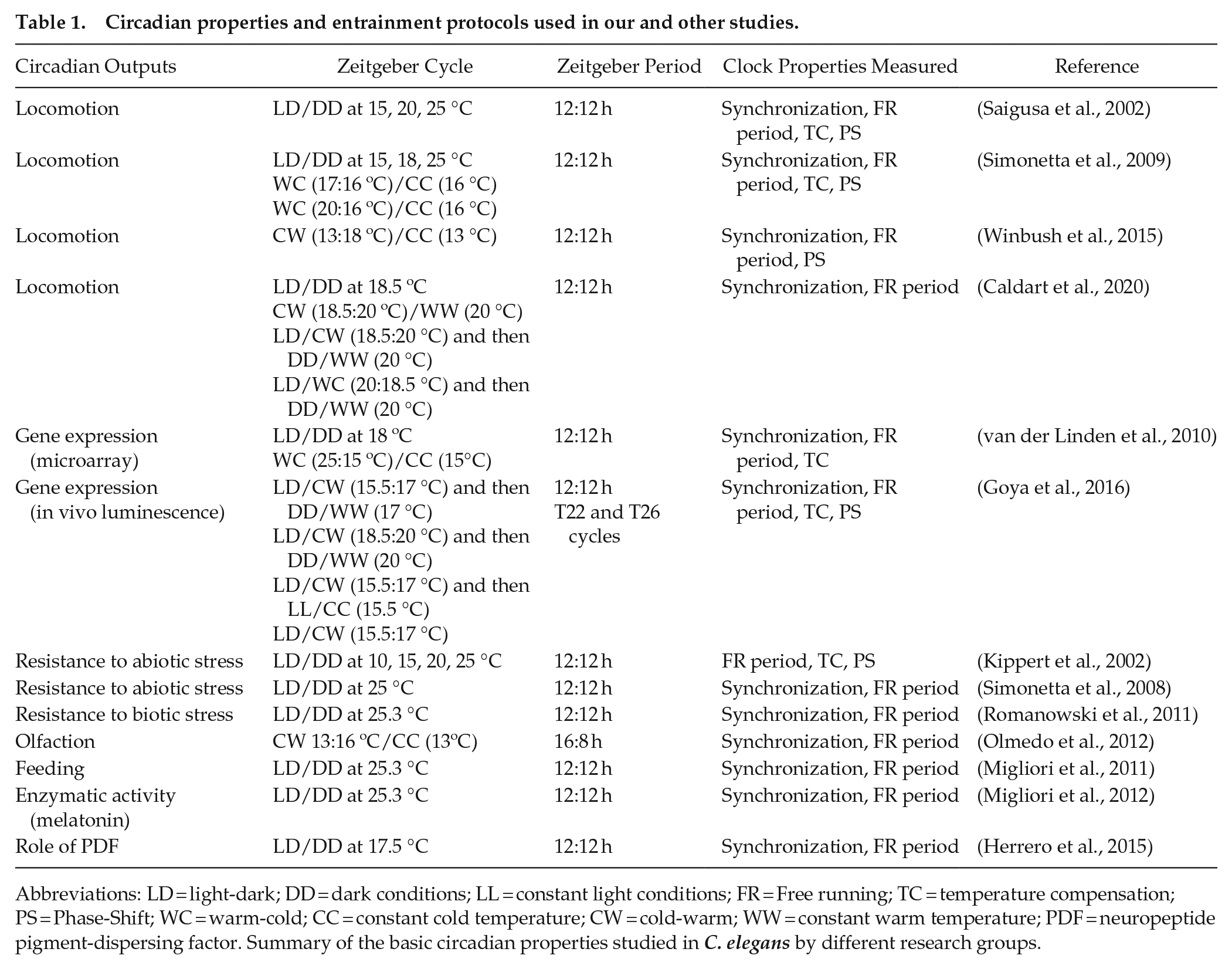
Abbreviations: LD = light-dark; DD = dark conditions; LL = constant light conditions; FR = Free running; TC = temperature compensation; PS = Phase-Shift; WC = warm-cold; CC = constant cold temperature; CW = cold-warm; WW = constant warm temperature; PDF = neuropeptide pigment-dispersing factor. Summary of the basic circadian properties studied in C. elegans by different research groups.
In mammals and Drosophila, sleep and wake states are governed by a dual process involving homeostatic state and the circadian clock. C. elegans has two states of sleep: developmentally timed sleep (DTS) or lethargus, and stress-induced sleep (SIS) (Trojanowski and Raizen, 2016). During lethargus, which lasts almost 3 h, nematodes show evident changes in development and morphology (including vulval eversion, cuticle shedding/remodeling, as well as regulated seam cell and hypodermal/cuticle cell divisions). Feeding and locomotion cease during DTS. Unlike the circadian sleep in Drosophila and mammals, DTS occurs with an ultradian periodicity (it occurs four times during development, with a period of 8-10 h depending on the cultivation temperature) (Mata-Cabana et al., 2022) and is linked to the molting cycle rather than the 24 h light/dark cycle. Despite this difference, many molecular pathways regulating sleep in mammals and flies also regulate sleep in C. elegans, such as the PDF, PKA/cAMP, EGFR, dopamine, and PKG signaling pathways (Moosavi and Hatam, 2018; Trojanowski and Raizen, 2016). The nematodes greatly reduce the release of PDF-1 during DTS suggesting that PDF signaling promotes arousal in C. elegans, as they do in Drosophila (Renn et al., 1999). PKA/cAMP signaling pathway is known to promote wakefulness in mammals and flies (Graves et al., 2003; Hendricks et al., 2001). C. elegans mutants with increased PKA signaling are hyperactive during DTS (Trojanowski and Raizen, 2016). Moreover, dopamine release promotes wake in mammals and Drosophila; in nematodes, mutations in the dopamine D1 receptor gene dop-1 cause increased quiescence (Trojanowski and Raizen, 2016). On the other hand, EGF signaling promotes sleep and PKG activity is associated with sleep or sleep intensity. The overexpression of the EGF homolog LIN-3 causes cessation of feeding and locomotion in C. elegans (Moosavi and Hatam, 2018). EGF signaling appears primarily necessary for SIS, rather than DTS. In sensory neurons of C. elegans, PKG functions to induce quiescence and reduced responsiveness during lethargus (Raizen et al., 2008). However, in this nematode, the timing of lethargus appears to be regulated by developmental stage, an internal time giver, rather than the light/dark cycle, an external time giver. On the other hand, SIS occurs when nematodes are exposed to environmental factors that generate cellular stress, and its duration depends on the severity of the stressor (Hill et al., 2014; Jones and Candido, 1999). SIS is also characterized by a cessation in locomotion and feeding and can occur at any time during the development of the larvae and the adult stage (Hill et al., 2014). Although C. elegans shows circadian rhythms of behaviors, metabolism, physiology, and gene expression (Migliori et al., 2012, 2011; Olmedo et al., 2012; Saigusa et al., 2002; Simonetta et al., 2009, 2008; van der Linden et al., 2010; Winbush et al., 2015), it has not been demonstrated that SIS and sleep behavior are under circadian control.
Clock Homolog Genes in C. elegans
The biological clock in C. elegans has not yet been characterized at the molecular level. However, some proteins have been described in C. elegans as putative homologs to the mammalian/flies core clock genes, such as AHA-1, LIN-42b/c, TIM-1, and KIN-20, which show different degrees of homology to CLOCK/BMAL1, PER, TIM y CK1ε/δ, respectively (Romanowski et al., 2014; Temmerman et al., 2011) (Table 2). These genes have key roles in the development of nematodes (Chan et al., 2003; Jeon et al., 1999; Qin and Powell-Coffman, 2004; Tennessen et al., 2006), as their counterparts do in mammals, including during the synthesis of the new cuticle—molts—(Monsalve et al., 2011), but their role in the circadian clock of C. elegans has not yet been studied in depth. Furthermore, circadian rhythmicity at the mRNA level has not been found for any of these possible homologous genes in adult worms (Olmedo et al., 2012; van der Linden et al., 2010), at least under classic light/dark or temperature cycles and in total RNA extracts of whole animals. It remains possible, however, that orthologous clock genes cycle in a small subset of cells or tissues in worms. Moreover, hybridization-based approaches such as microarrays have been the standard gene expression profiling technology for use in research; however, they have low sensitivity and only profiles predefined transcripts/genes through hybridization. Another alternative approach to identify light- and temperature regulated transcriptional rhythms in C. elegans could be through RNAseq.
Homologous proteins of biological clock core and accessory clock proteins in Drosophila and mammals.
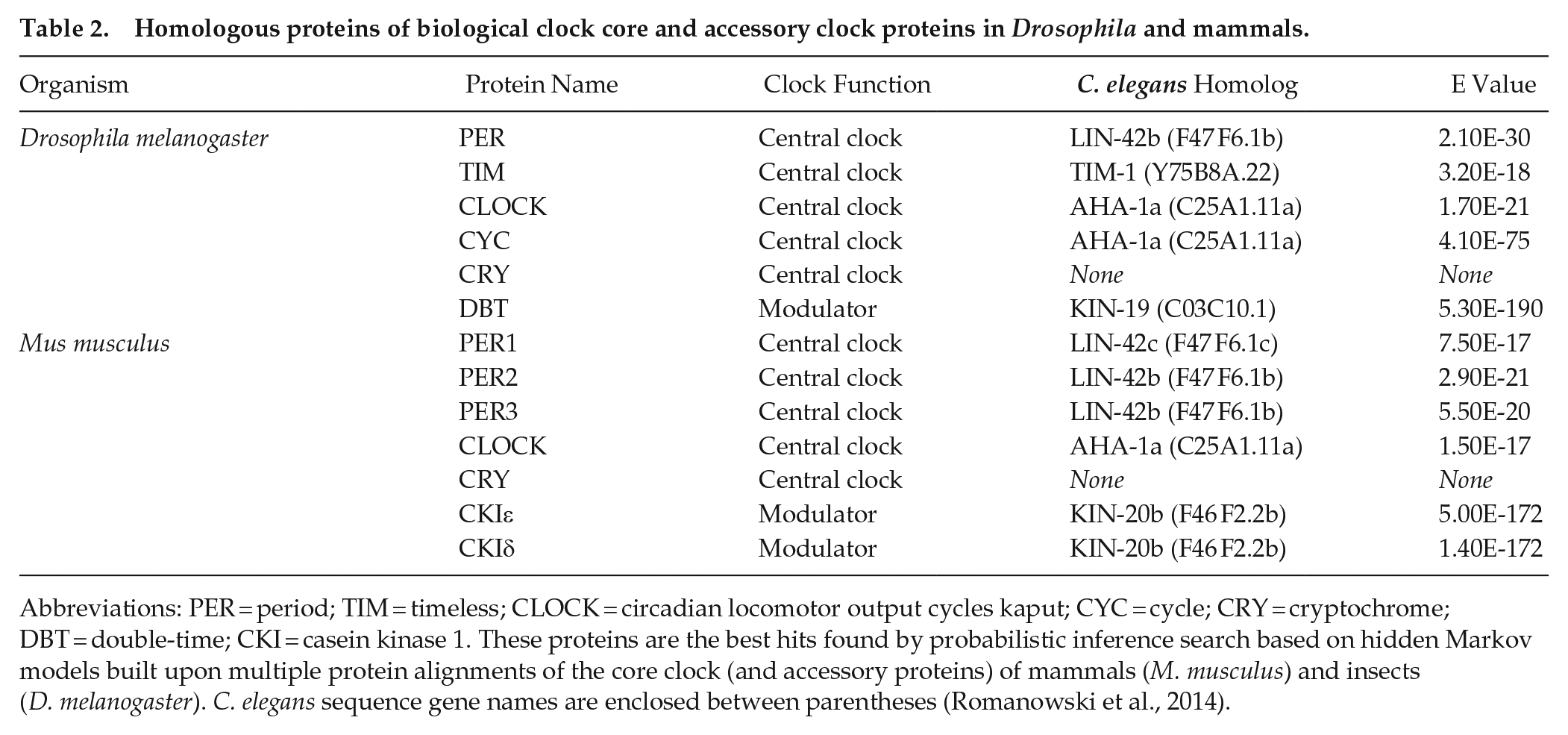
Abbreviations: PER = period; TIM = timeless; CLOCK = circadian locomotor output cycles kaput; CYC = cycle; CRY = cryptochrome; DBT = double-time; CKI = casein kinase 1. These proteins are the best hits found by probabilistic inference search based on hidden Markov models built upon multiple protein alignments of the core clock (and accessory proteins) of mammals (M. musculus) and insects (D. melanogaster). C. elegans sequence gene names are enclosed between parentheses (Romanowski et al., 2014).
As stated before, CKIε plays an important role in the regulation of core clock proteins of mammals. C. elegans expresses only one homologous protein to the mammalian CK1ε/δ (albeit with several isoforms), encoded by the protein kinase gene kin-20 (Romanowski et al., 2014). KIN-20 functions as a member of the heterochronic pathway which regulates developmental timing, intervening in the morphogenesis of the body, embryonic and larval development, growth and reproduction (Banerjee et al., 2005). We have shown that treatment with a CK1ε/δ inhibitor lengthens the period of the luminescent rhythms in C. elegans (Goya et al., 2016). These results suggest the existence of a conserved mechanism of post-translational modulation of the circadian clock in C. elegans.
On the road toward clock genes discovery in the nematode, mutagenesis and genetic screening are invaluable tools. Not only does C. elegans have a completely sequenced and annotated genome, but a wide-array of different mutants are available (from small- and large-scale mutant screens) to perform genetic and molecular studies. To identify possible components of the circadian molecular clock of C. elegans, nematode mutants of homologs of other organisms’ clock genes can be used to measure possible outputs of the clock. In C. elegans, the PER homolog lin-42 is an important member of the heterochronic pathway. While LIN-42 does not exhibit rhythmic expression in whole animal profiling experiments under LD (light-dark) or WC (warm-cold) cycles (van der Linden et al., 2010), evidence in favor of the possible role of LIN-42 in the central clock of C. elegans comes from the observation that nematodes mutant for this gene show a longer period of locomotor activity as compared to wild-type nematodes (Simonetta et al., 2009).
The N-terminal of LIN-42 contains PAS domains similar to the ones found in PER (Romanowski et al., 2014), while the C-terminus of LIN-42 also contains two highly conserved kinase binding motifs that interact with mammalian CK1, suggesting that the domains that anchor the kinase to PER2/LIN-42 may work in a similar way.
Synchronization of Circadian Rhythms
In mammals, light is transmitted by means of the retinohypothalamic tract (RHT) to the central clock located in the suprachiasmatic nucleus (SCN). The SCN act as a master pacemaker to synchronize peripheral oscillators (Green et al., 2008). Drosophila possesses peripheral clocks that can be directly entrained by environmental cues, located in the head and body (George and Stanewsky, 2021). In these animals, the Cryptochrome act as blue light photoreceptors—in addition to being part of the molecular oscillation mechanism—and are encoded by the cry gene.
In addition to light, which is considered the most important zeitgeber, it has been shown that temperature cycles can also entrain locomotor rhythms in vertebrate and invertebrate organisms (Dubruille and Emery, 2008; George and Stanewsky, 2021; Winbush et al., 2015). In Drosophila, temperature differences of 2 to 3 °C are sufficient to entrain locomotor activity rhythms, but the molecular mechanism is not yet understood in detail (George and Stanewsky, 2021). In C. elegans, different studies showed that both light and temperature are very important stimuli capable of entraining the central clock, both for rhythms of locomotor activity (Caldart et al., 2020; Simonetta and Golombek, 2007; Simonetta et al., 2009) and gene expression (Goya et al., 2016; van der Linden et al., 2010). Light and temperature signals are also integrated by the central clock in Drosophila and both stimuli act cooperatively to entrain the circadian clock and generate patterns of rhythmic activity (Boothroyd et al., 2007).
In C. elegans, LITE-1 and GUR-3 (both noncanonical photoreceptors of the GPCR gustatory receptor family) are capable of sensing light directly or indirectly, respectively (Bhatla and Horvitz, 2015; Edwards et al., 2008; Gong et al., 2016). LITE-1 is a receptor with high affinity for ultraviolet light (UVA and UVB), which mediates the negative phototaxis induced by short wavelengths (ultraviolet, violet and blue, 400-500 nm) and the inhibition of pharyngeal pumping in the presence of daylight (Bhatla and Horvitz, 2015). GUR-3 functions as a high affinity receptor for hydrogen peroxide, which can be generated as a byproduct of light, and is also involved in photic inhibition of pharyngeal pumping. Unlike LITE-1, GUR-3 is activated only by very bright unnatural light; therefore, this pathway could function naturally in environmental sensing of hydrogen peroxide produced by soil microorganisms and/or other reactive oxygen species and not in natural light detection (Bhatla and Horvitz, 2015). Although photoreceptors are expressed in many neurons in the nervous system of the nematode, the ASJ, AWB, ASK, and ASH sensory neurons have been reported to play a major role as photoreceptor cells for phototaxis in a redundant manner (Ward et al., 2008). The phototransduction cascade has been studied in more detail for the ASJ neuron. Light is sensed by the LITE-1 photoreceptor, generating downstream activation of at least two G alpha proteins (GOA-1 and GPA-3) and the guanylate cyclases DAF-11 and ODR-1. This leads to the upregulation of the second messenger cGMP followed by opening of cGMP-sensitive CNG channels formed by TAX-2/TAX-4. The increase in intracellular Ca+2 induces a downstream signaling cascade that ends with negative phototaxis (Liu et al., 2010) (Figure 2a).
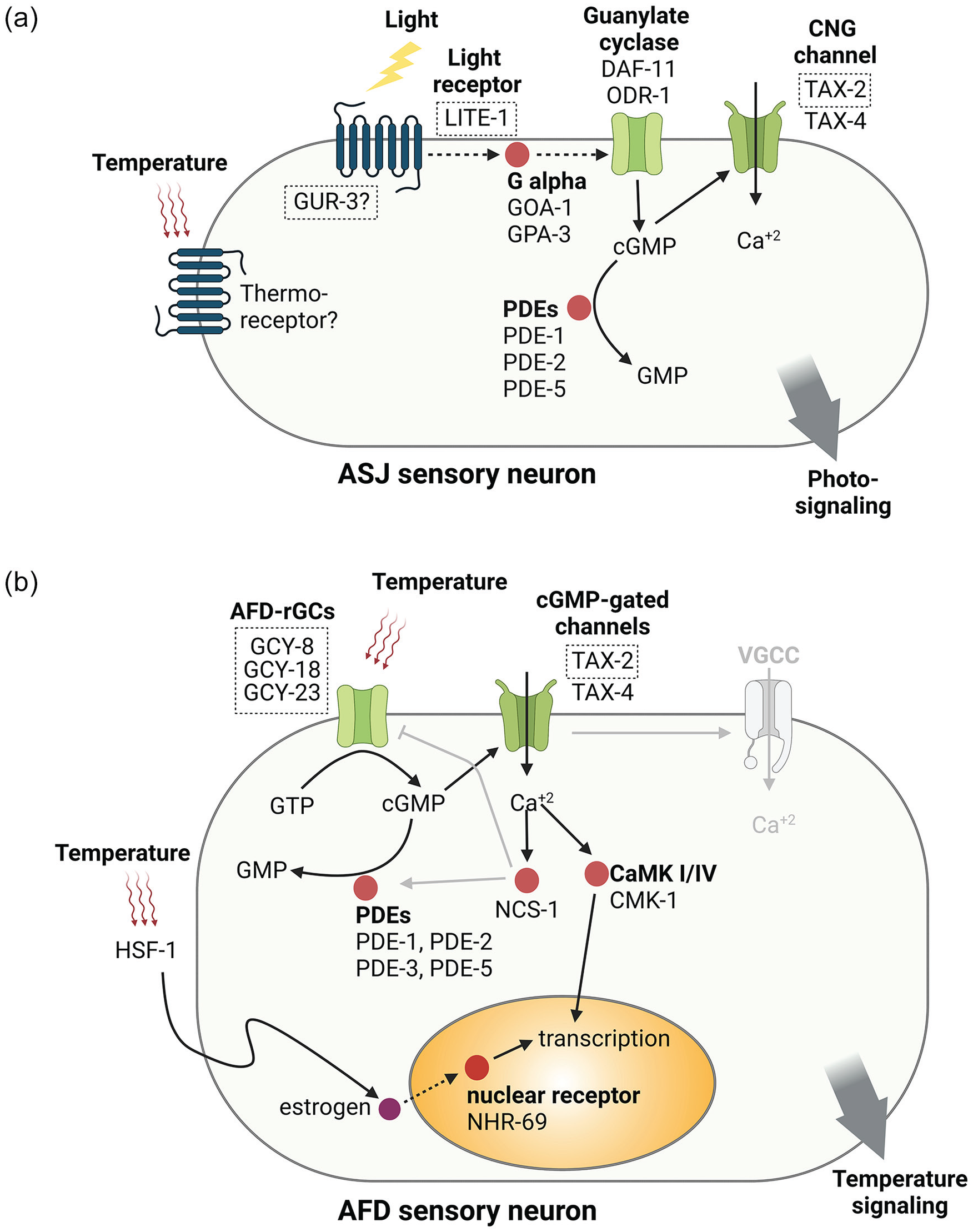
Temperature and light signaling models in the sensory neurons ASJ and AFD. a) Light is sensed by the photoreceptor LITE-1 generating downstream activation of at least two G-alpha proteins (GOA-1 and GPA-3) and the guanylate cyclases DAF-11 and ODR-1. This leads to the up-regulation of the second messenger cGMP followed by opening of cGMP-sensitive CNG channels formed by TAX-2 and TAX-4. PDEs (PDE-1, 2 and 5) acts as an inhibitor of light signaling by hydrolyzing cGMP to GMP. ASJ also contributes to thermosensory signaling via a G protein-coupled temperature receptor, which remains to be identified, for performing cold tolerance. b) In sensory neuron AFD, temperature is sensed by rGCs (GCY-8, 18 and 23) that convert GTP to cGMP, which then opens cGMP-gated channels (TAX-2 and 4). Calcium influx promotes CMK-1 translocation to the nucleus, which drives gene expression. cGMP is degraded by PDEs (PDE-1, 2, 3 and 5). The involvement of VGCCs and the feedback mechanism of NCS-1 are still unclear. Systemic estrogen signaling also mediates gene expression in AFD to regulate thermotaxis. Molecular names shown inside dotted boxes indicate C. elegans proteins that have been recently shown to be necessary for circadian clock synchronization in nematodes. Abbreviations: PDEs = phosphodiesterases; AFD-rGCs = AFD specific guanylyl cyclases; VGCCs = voltage-gated calcium channels; cGMP = cyclic guanosine monophosphate; CNG = cyclic nucleotide-gated; GTP = guanosine-5’-triphosphate.
On the other hand, nematodes are very sensitive to changes in temperature (Goodman and Sengupta, 2019). The main neurons involved in thermoreception are the sensory neurons AFD, along with other sensory neurons, AWC, and ASI. Genetic or laser ablation of AFD suppresses all thermotaxis behaviors (Takeishi et al., 2020). In the AFD neuron, temperature activates three guanylate cyclases (GCY-8, GCY-18, and GCY-23) generating an increase in cGMP and activation of the TAX-2/TAX-4 channel (Takeishi et al., 2020); however, purely thermosensory proteins are still unknown (Figura 2B). Although the molecular mechanisms and neural circuits involved in sensing light and temperature are known, the interaction between these and the C. elegans circadian system are still not understood. Some components involved in sensing light (LITE-1 and GUR-3), and in light and temperature transduction (TAX-2), have recently been shown to be necessary for clock entrainment (Caldart et al., 2020; Goya et al., 2016; van der Linden et al., 2010; Winbush et al., 2015).
Circadian Rhythms of Locomotor Activity in C. elegans
C. elegans adults reach a length of about 1 mm, and this characteristic has been a problem for the recording of its locomotor activity. The first work in which locomotor activity rhythms were measured in these nematodes consisted of filming their movement and the subsequent digital processing of the images obtained (Saigusa et al., 2002); however, it is not particularly useful for mutant screening and circadian characterization. Different systems have been developed for long-term recordings of C. elegans locomotor activity, including infrared light sensor-based trackers (Simonetta and Golombek, 2007), which allow ~2 week recordings under different experimental conditions, such as different light and temperature schedules, as well as the screening of specific mutants. In addition, an imaging system for long-term automated locomotor activity recordings was described, which also allows for long-term recording (Winbush et al., 2015). Both systems are simple and quick to execute and allow the rhythms of animals in free movement to be recorded; however, the circadian rhythms of locomotion do not appear to be robust since they do not persist for more than 1 day under free running conditions. One of the possible reasons for this could be that in both systems the signal-to-noise ratio is low and makes it difficult to record for long periods of time. Because of the importance of locomotion as a circadian readout, we describe in this section how to determine circadian period and study light and temperature inputs.
Synchronization to Light-Dark Cycles
Circadian rhythms need to be continuously entrained (synchronized) with the environment in order to cope with daily changes in UV light intensity, food availability, or the presence of predators. A shift in the synchronizer (zeitgeber, such as the light-dark [LD] or temperature cycle) results in a gradual alignment of the clock to the new cycle. Furthermore, behavioral patterns can also respond to changes in the environment independently of the circadian clock, a phenomenon called masking (Golombek and Rosenstein, 2010). Previous studies have shown that C. elegans perceives its environment through chemosensation, mechanosensation, osmosensation, and thermosensation (mechanisms commonly used by subterranean animals) (Iliff and Xu, 2020); however, C. elegans may spend much of its time above ground, living on small surface-dwelling animals or their carcasses (Schulenburg and Felix, 2017). C. elegans may therefore be frequently exposed to direct sunlight, which can damage cells by photooxidative reactions (De Magalhaes Filho et al., 2018).
Indeed, our laboratory demonstrated that locomotor activity in adult C. elegans (wild-type strain) is entrained to LD cycles (Caldart et al., 2020; Simonetta and Golombek, 2007; Simonetta et al., 2009). Nematodes subjected to a normal 24 h LD cycle showed an activity pattern of 24 h in at least 50% of the populations tested, 40% of which maintained such rhythmicity in constant conditions (DD). Under this entrainment, higher locomotor activity was registered during the dark phase of the cycle, suggesting that C. elegans is nocturnal under laboratory conditions (circadian periodicity showed a population distribution of periods centered at 23.9 ± 0.4 h and was temperature-compensated) (Simonetta et al., 2009). While about 50% of the populations were clearly entrained by light, the remaining populations analyzed showed a masking phenomenon: when the zeitgeber was removed, the free-running rhythms began in different phases than the one determined by the LD cycle. In addition, worms re-entrained after 6 h phase shifts in the zeitgeber. For analysis purposes, we defined “synchronized populations” as the number of populations with a period and phase set by the zeitgeber conditions, and “entrained populations” as the number of populations that retained their circadian phase when placed under constant conditions (free running). Finally, synchronized populations with a free running phase that is not determined by the previous cycling conditions are defined as “masked populations” (please refer to Bittman, 2021). As for single-worm assays, the average period for locomotor activity of individual nematodes was 24.2 ± 0.44 h in LD and 23.1 ± 0.40 h in DD (Simonetta and Golombek, 2007).
Population recordings (approximately 50 nematodes per well) can provide valuable information about social entrainment and the effect of inter-individual interactions. It should be noted that single-nematode recordings, although possible under certain conditions, are quite challenging to perform because of the small size of the nematodes and their high motility rate. Moreover, signal/counts from single nematodes are far lower compared to those of a population, which greatly reduces the signal-to-noise ratio and makes it difficult to register for long periods of time. Even so, single-nematode assays are of great value to analyze individual response to the action of a particular zeitgeber and to estimate circadian period more precisely. This allows to avoid the averaging effect that results in the loss of information caused by individual desynchrony within a population. Yet, single-nematode assays generally show more variability and require a larger number of replicates (Goya et al., 2016; Simonetta and Golombek, 2007).
Locomotor activity rhythms under LD cycles and subsequent DD in wild-type worms have also been characterized by other laboratories. Saigusa et al. used a bug tracker to record the footprints of 3rd and 4th stage larvae and adult worms in order to analyze rhythmic locomotion patterns, again finding ~24 h rhythms peaking during the day and sustained under constant dark conditions (Saigusa et al., 2002). C. elegans switches between distinct forms of locomotion—crawling versus swimming—when transitioning between solid and liquid environments. These forms of locomotion are distinguished by different underlying patterns of neuromuscular activity (Pierce-Shimomura et al., 2008). When the locomotor activity of nematodes is recorded on solid medium under LD cycles, they show high locomotor activity during the day (Saigusa et al., 2002). Interestingly, when the recordings are carried out in liquid medium, this locomotor behavior pattern is altered, showing greater activity at night (Simonetta and Golombek, 2007; Simonetta et al., 2009). While both types of movements can be used to characterize locomotor activity rhythms in nematodes, the results obtained cannot be compared.
Although the molecular mechanisms and neural circuits involved in sensing light are known in C. elegans, the fundamental components of the entrainment pathways to the LD cycle are still unknown. Our laboratory performed circadian screenings in order to assess the entrainment capability of different mutant strains that exhibit defects in photic responses (Caldart et al., 2020). A lower responsiveness to light was observed for different strains carrying mutations for lite-1/gur-3. As stated before, these genes have been involved in sensing light.
Synchronization to Temperature Cycles
In their ecological niche C. elegans are subjected to daily temperature fluctuations as a consequence of the solar energy’s incidence on the ground (Robinson, 1994). Thus, temperature might be of relevance to entrain its circadian clock. Our laboratory demonstrated that C. elegans populations showed entrainment to temperature cycles of 17:16 °C (WC 12:12 h) under constant dark conditions (Simonetta et al., 2009). Under WC cycles with larger temperature variations (20:16 °C), the amplitude of the rhythms was higher and a decrease in activity was observed after the transition from higher to lower temperature values. Other temperature cycles were also tested in our laboratory in order to find the best temperature conditions capable of entraining rather the locomotor activity rhythms in adult worms (Caldart et al., 2020). Moreover, Winbush et al. (2015) used a new method based on an imaging system for long-term locomotor activity recordings. With this new setup, when nematode populations were entrained to temperature cycles of 13:18 °C (WC 12:12 h) under constant dark conditions, robust rhythms with peak activity during the warm (18 ºC) phase were observed, with a 24.2 h period. A 6-h phase shift in temperature cycles suggested that locomotor activity shows both entrainment and masking features (Winbush et al., 2015). Worms present anticipatory activity several hours before the cold-warm shift, which corresponds to entrainment, and a high amplitude response during the actual shift between temperatures, which corresponds to masking. Under constant conditions (13 °C), nematodes maintained their rhythms for 1 day, but these dampened quickly after. All these data suggest that temperature is also a strong signal for the circadian system of C. elegans, capable of entraining locomotor activity rhythms in these nematodes.
On the other hand, our laboratory performed circadian screenings in order to assess the entrainment capability of different mutant strains that exhibit defects in thermal responses (Caldart et al., 2020). A lower responsiveness to temperature was observed for different strains carrying mutations for gcy-8, 18 and 23. As stated before, these genes have been involved in sensing temperature. tax-2 (p671) mutants, which have impaired thermoreception, were also tested by van der Linden and colleagues (Winbush et al., 2015). Their results showed that loss of tax-2 alters locomotor activity of C. elegans during temperature entrainment, although period analysis showed no clear circadian pattern. All these results show that this group of genes could be involved in the synchronization pathway of this nematode, modifying its behavioral output.
Dual Zeitgeber Condition for Entrainment
As mentioned before, C. elegans is exposed to daily changes in both light and temperature, as well as in oxygen supply, humidity and nutrients; all these factors could contribute to circadian entrainment. Considering nematode entrainment to light-dark or temperature cycles, this opens the door to an interesting question: could a combination of both zeitgebers robustly entrain the biological clock of C. elegans? Recently, our laboratory designed a different behavioral approach in order to complete the characterization of circadian entrainment of adult C. elegans under light and temperature cycles. In its ecological niche, and in particular beneath the soil, the temperature increase is phase-delayed with respect to sunrise, reaching its maximum in the late afternoon (Monteith and Unsworth, 2013). Thus, the hours with higher temperature are concentrated toward the end of the day, when solar radiation is minimal but the soil has absorbed most of it. In this context, we designed a dual zeitgeber condition to emulate the nematodes underground environment, where higher temperatures correspond to darkness and lower temperatures to light, normal 24 h LD cycle / CW (18.5:20 °C,) (Caldart et al., 2020; Goya et al., 2016). Under this condition, approximately 80% of the populations assayed are synchronized to the dual cycle; 50% of which maintained such rhythmicity in constant conditions (DD/WW), suggesting that nematodes are able to entrain to the dual cycles, even if part of the populations exhibits masking. The advantage of the dual cycles lies in their ability to entrain the nematode from two different pathways. Different neurons are responsible for thermoreception and photoreception, yet they are integrated by the neurons RIM, RIA, and RIB, which lead to locomotor output (Takeishi et al., 2020). This way, mutants such as lite-1/gur-3, which have impaired photoreception, and mutants like gcy-8, 18 and 23, with impaired thermoreception, can still entrain via the alternative zeitgeber (Caldart et al., 2020). Understanding how environmental cues are integrated by the clock is important if we are to elucidate the mechanisms required for entrainment under natural conditions.
Circadian Variation in Stress Resistance
As mentioned before, circadian rhythms control several behaviors through neural networks, hormones, and gene expression. One of these outputs in animals is the stress resistance behavior. In many organisms, abiotic and biotic environmental stressors play a key role in their ecophysiology, and since stress-inducing agents vary along the day, it is logical that organisms must be able to predict these periodic changes in order to survive. Kippert et al. (2002) provided the first evidence of a circadian clock in C. elegans larvae, by studying the response to hyperosmotic stress throughout the day, which was rhythmic under LD and DD conditions and could be entrained to an experimental photoperiod (Kippert et al., 2002). We have also been able to describe the rhythmic resistance to osmotic and oxidative stress in adult C. elegans measuring withdrawal in response to tapping after a stressful condition (Simonetta et al., 2008). In addition, circadian expression of genes related to the control of stress responses have been shown in adult nematodes (Simonetta et al., 2008). Similarly, the biotic stress response caused by Pseudomona fluorescens also follows a circadian pattern in adult worms (Romanowski et al., 2011).
Feeding, Olfaction and Oxygen Consumption
In many organisms, physiological and metabolic processes are driven by the circadian clock (Dubowy and Sehgal, 2017; Leypunskiy et al., 2017; Serin and Acar Tek, 2019) and C. elegans is no exception. Thus, the circadian control of these variables is important so that nematodes can optimize the use of their energy, along with the ability to adapt to the cyclical changes of light, temperature, and nutrients in their environment. On an agar surface in the presence of abundant bacteria, normal worms exhibit a continuous pharyngeal pump activity at an average rate of about 260 pumps/min (Avery and Horvitz, 1989; Trojanowski et al., 2016). This behavior can be measured throughout the day counting the number of pharyngeal muscle contractions for a specific time period. Our laboratory demonstrated daily and circadian rhythms in muscle contraction of the pharynx in adult nematodes under LD and DD conditions (Migliori et al., 2011). Furthermore, this study showed daily variations in feeding rate and defecation behavior of adult nematodes under LD cycles. Circadian rhythms in oxygen consumption have also been studied in C. elegans. By using a polarographic O2 electrode, adult nematode populations showed a daily fluctuation in oxygen consumption under an LD cycle, which was maintained under constant conditions (DD) (Migliori et al., 2011).
Merrow and colleagues (Olmedo et al., 2012) used a C. elegans chemorepellent, 1-Octanol, in order to measure the variation of the olfactory response along the day. In this work, a culture protocol was designed using temperature as a zeitgeber capable of entraining the circadian clock of the nematodes and mimicking the regular daily cycles that would occur in the soil. Thus, daily and circadian oscillations in olfaction were observed in adult worms, suggesting that this sensory function could be regulated by the circadian clock. Indeed, olfaction is also clock-regulated in insects and mammals (Granados-Fuentes et al., 2006; Tanoue et al., 2008). C. elegans uses olfaction in order to spatially orient within their environment, and thus colonize new food sources (Zhang et al., 2005), and find sexual partners (Edison, 2009). In this sense, the circadian control of olfaction would allow nematodes a better adaptation/performance in their ecological niche.
A Bioluminescence Reporter-Based Method
The development of methods based on high-throughput analysis, such as long-term recordings of the locomotor activity of C. elegans, made possible the screening of circadian mutants, as well as mutant strains with defects in photic and thermal responses (all available at the Caenorhabditis Genetics Center). Our laboratory also developed a luminescence-based method to monitor rhythms of either single worms or populations of around 100 animals. Using this system, it was shown that fundamental properties of circadian rhythms present in other organisms such as entrainment, rhythmicity under free running, entrainment after a 6-hour phase-shift, and temperature compensation, also apply to C. elegans (Goya et al., 2016).
Luciferases are a class of enzymes that produce light upon oxidation of their substrates, and they are commonly used in biology as reporters of transcription or translation. Luciferase-based methods are broadly used to monitor circadian clocks in various organisms (Welsh et al., 2005). We generated transgenic strains carrying the firefly luciferase cDNA fused to the ORF of the green fluorescent protein (GFP) under the control of a fragment of the sur-5 gene promoter (Goya et al., 2016). This particular promoter was chosen because it exhibits a strong, constitutive, and ubiquitous somatic expression throughout all developmental stages of C. elegans. Our study using such a luminescence-based transgene showed that both population and single nematodes express ~24 h rhythms, which can be entrained by LD and CW cycles, and that these rhythms are temperature compensated and can be re-entrained after phase changes of synchronizing agents. Later, the expression of the bioluminescence reporter in different mutants deficient in thermo—and/or photo—reception was analyzed to further understand how the C. elegans’ clock senses environmental cues (Goya et al., 2016). Thus, our bioluminescence reporter revealed that LITE-1, GUR-3 and TAX-2 proteins are components involved in the photic and temperature synchronization pathways of the central oscillator. This method proved to be more reproducible than locomotion-based methods, allowing to record robust circadian rhythms in gene expression which persists even after 2 days under free running conditions, even from single worms. It is also simple and fast to run and allows to record rhythms from free moving animals. However, luminescence recordings also showed a low proportion of entrainment among populations assays, of around 50% under the best entrainment conditions found and a high variability of the period under free running conditions, making it very challenging for screening purposes. It should be noted that although this could be a consequence of using the promoter of an output gene, which expression could be not as robust as a central clock gene, this seems not be a limitation of the method per se, but rather a common feature seen with all the methods so far related to the intrinsic biology of the worms. A systematic analysis of different entrainment conditions such as by using different light intensities and temperature variations combined with this very sensitive luminescence-based method could perhaps help find better entrainment conditions to reduce inter-individual variability. As mentioned before, another alternative approach, would be to study luminescent rhythms in less domesticated strains, with the hope of finding less variability from animals with a more ancestral clock, recently exposed to natural conditions. In addition, adapting this method for doing RNAi based on feeding, which could allow the use of commercially available libraries to knock down most of the genes of C. elegans’ genome, will make it more amenable for a whole-genome screening. This will greatly increase our changes of unveiling components of the C. elegans clock.
Output Signals of the Circadian Clock
In mammals, environmentally cued circadian rhythms of behavior and physiology are generated by the outputs of the core clock circuit, which are organized by peptidergic signaling from a few pacemaker neurons (Aton and Herzog, 2005). In many insects, this peptide is PDF (Pigment Dispersing Factor), whose role is strikingly similar to VIP in mammals. The circadian functions of PDF have been studied in detail in Drosophila (Yoshii et al., 2009). In the fly, small ventral lateral neurons (s-LNv) are major pacemaker neurons that are essential for robust rhythmic activity under constant darkness. Most s-LNv express PDF, which synchronizes oscillations throughout the brain, and also influence oenocyte rhythms in the fly body. The PDF receptor (PDFR) is expressed in a subset of clock neurons distributed throughout the network. Flies with mutations in either PDF or PDFR have altered behavioral phenotypes, with no morning peak of activity and an early evening peak of activity in LD, and short-period rhythms that dampen quickly in DD (Yoshii et al., 2009). In C. elegans, two pdf homologs—pdf-1 and pdf-2—have been identified (Janssen et al., 2009), which resemble the crustacean PDH (Pigment Dispersing Hormone) system (Meelkop et al., 2012a) and whose products interact with different splice forms of the specific PDF receptor (PDFR) (Janssen et al., 2008). The C. elegans PDF peptides differ in length when compared with arthropod family members; however, the core sequence necessary for binding to a crustacean PDH receptor is preserved in C. elegans PDF-1 peptides (Meelkop et al., 2012a). In nematodes, PDF is involved in different biological processes such as male reproduction, egg laying, and locomotion (Barrios et al., 2012; Meelkop et al., 2012b). The role of PDF in the circadian system of nematodes was approached in recent years. Janssen et al. (2009) provided the first evidence that all three C. elegans pdf transcripts (pdf-1a, b and pdf-2) are consistently expressed over the course of 24 h in LD, which is in agreement with the observations in Drosophila (Park and Hall, 1998). Also, the C. elegans PDFs are expressed throughout postembryonic life in at least 19 types of neurons and a few non-neuronal tissues (Janssen et al., 2009). Moreover, we have demonstrated the role of PDF in the genesis and entrainment of locomotor activity circadian rhythms in C. elegans. pdf-1 and pdf-2 mutant strains showed abnormal locomotor activity outputs. In particular, pdf-1 mutant induced circadian arrhythmicity under both LD and DD conditions, which was rescued by a genomic copy of the pdf-1 locus (Herrero et al., 2015). The evidence that nematodes possess a functional PDF signaling pathway, and that PDF-1 is involved in the control of daily locomotor rhythms in C. elegans, suggests that this neuropeptide could contribute to the synchronization and transmission of signals from the circadian clock of nematodes as was shown in Drosophila.
Also, melatonin is a molecule that has a prominent role within circadian rhythms. This indoleamine’s expression peaks during the night and is suppressed by light, which effectively means that melatonin signals darkness. Furthermore, melatonin has a ubiquitous distribution in nature, as it has been found in bacteria, fungi, plants, invertebrates, and vertebrates (Hardeland et al., 2006). In vertebrates, this hormone is synthesized from tryptophan, through several reactions including hydroxylation, decarboxylation, N-acetylation, and O-methylation. The N-acetylation of serotonin by the arylalkylamine N-acetyltransferase (AA-NAT) is the limiting step in this pathway. This enzyme is inhibited by light in vertebrates, giving melatonin its characteristic rhythms (Amaral and Cipolla-Neto, 2018). In the mammalian circadian system, melatonin acts as a neuroendocrine hormone that transduces signals from the environment through membrane-bound receptors (MT1 and MT2 receptors), and several other binding sites whose function is not completely understood (Amaral and Cipolla-Neto, 2018). Tanaka et al. (2007) examined the effects of exogenous melatonin on C. elegans behavior and found that this molecule affects locomotion rate via MT1-like receptors. Furthermore, it was reported that nematodes are able to synthesize melatonin and a gene that possibly encodes a C. elegans ASMT enzyme has been identified (Tanaka et al., 2007). Other authors have shown daily variations of melatonin levels in C. elegans, which under LD conditions showed a rhythmic pattern with high levels of the indoleamine mostly during darkness (Balzer et al., 2000). We have described daily variations in melatonin synthesis in adult nematodes under LD cycles, finding higher levels of the hormone around the middle of the night (Migliori et al., 2012). Furthermore, we have shown a daily and circadian variation in AA-NAT activity with higher levels of activity during the early night, both under LD and DD conditions. In vertebrates, higher plants, and invertebrates of different taxa, melatonin acts as a free radical scavenger and antioxidant. We have reported that C. elegans is more resistant to oxidative stress during the night (Simonetta et al., 2008), and this temporal pattern of resistance correlates with the rhythm we found in the melatonin synthesis, suggesting that it might participate in protection mechanisms against oxidative stress in nematodes. Furthermore, C. elegans could use melatonin as a neuroendocrine molecule capable of sensing time, and thus adapt to changes in the environment by regulating its locomotion as well as other cyclical activities of the body.
Conclusions
Various model organisms have been used to decipher the molecular bases of circadian clocks, including cyanobacteria, Drosophila, zebrafish, and mice. C. elegans is a model organism that in recent years has been used for circadian research as well. Numerous advantages favor its use in chronobiology and make it a model of choice in studying circadian rhythms. Cycles in behavior, physiology, and gene expression have been reported in C. elegans by several groups. Furthermore, nematodes are able to entrain with a single zeitgeber (light or temperature), as well as to a particular combination of zeitgebers. The lack of robust and reliable methods to measure daily rhythms in C. elegans delayed the study of circadian rhythms in these animals. This is particularly relevant given that there is a large variability between individuals in C. elegans, yielding circadian outputs that may not appear as robust (Herrero et al., 2015; Kippert et al., 2002; Simonetta and Golombek, 2007; Winbush et al., 2015). In this sense, the locomotor activity recording system (Simonetta and Golombek, 2007) and the bioluminescence reporter for measuring gene expression in vivo (Goya et al., 2016) have provided a great opportunity to further circadian studies in the worm. Both methods complement each other and have allowed, by recording of either single worms or whole populations, to further understand the molecular bases of the circadian clock and the neuronal circuitry involved in synchronization.
Exhaustive bioinformatics analysis has identified proteins of relatively high homology to clock components from other species. Table 2 shows that C. elegans has almost all the protein components constituting the core of Drosophila and mammalian circadian clocks except for CRY (Romanowski et al., 2014). It is, however, interesting to note that a novel family of photoreceptors, unknown to other species, has been described in C. elegans, which may possibly fulfill a role related to CRY (Bhatla and Horvitz, 2015; Edwards et al., 2008; Gong et al., 2016). Furthermore, given that C. elegans orthologs of the clock proteins are involved in development and developmental timing, and the lack of rhythmicity in the expression of the lin-42 (per), tim-1 (tim) and aha-1 (clk/cyc) genes in whole-worm animal profiling experiments (Olmedo et al., 2012; van der Linden et al., 2010), we could hypothesize that in C. elegans the circadian clock might actually have a core design entirely different from that of insects and mammals. It remains possible, however, that orthologous clock genes cycle in a small subset of cells or tissues in worms, or that they are regulated distinctly by either post-transcriptional modifications (without relaying on transcriptional regulation) or by using alternative heterodimerization partners, as is the case in other organisms (Young and Kay, 2001). Nevertheless, different research groups have shown that the nematode’s circadian clock ticks through all stages of life from hatching through adulthood, demonstrating circadian rhythms in resistance to hyperosmotic stress in starved larvae (L1) after hatching, locomotor behavior in the adult stage, metabolism, and gene expression. In particular, daily oscillations found in the accumulation of RNA suggest that circadian regulation in C. elegans has some similarities to other clock model systems, at least at the level of clock regulation via transcription (Goya et al., 2016; Olmedo et al., 2012; van der Linden et al., 2010). Likewise, the circadian regulation in the redox state of the metabolic enzymes peroxiredoxins (PRX), which oscillates between reduced and oxidized forms in organisms from all phyla (Edgar et al., 2012), represents another phylogenetically conserved molecular marker of the circadian clock in C. elegans (Olmedo et al., 2012). The existence of multiple core designs of animal circadian clocks poses a further challenge to understanding the evolution of animal circadian clock systems.
Deciphering how the C. elegans’ circadian clock is able to sense environmental signals and regulate its rhythmic behaviors opens the door to discovering in which tissues and/or neurons are clock genes expressed. In most animals studied so far, there is a pacemaker and a molecular clock located in neurons; therefore, we could hypothesize that this will also be the case for homologous clock genes in C. elegans. As stated before, C. elegans is able to perceive light and temperature signals through photo and thermoreceptor neurons, respectively (Kimura et al., 2004; Mori and Ohshima, 1995; Ward et al., 2008). Moreover, both environmental stimuli are enough to entrain rhythmic behaviors in worms. Thereby, the zeitgebers—in this case daily cycles of light-darkness and temperature—could be sensed by specific photo and thermoreceptor neurons of the nematode that finally carry through the entrainment of the central clock. Using bioinformatic tools that allow us to find all genes expressed in a given cell type, such as the ceNGEN (C. elegans Neuronal Gene Expression Network), we generated a list of candidate neurons in which tax-2, tax-4 and lite-1 genes are expressed (Table 3). This list was then contrasted with functional and expression studies based on the literature. In the future, removing, turning off, or affecting the function of these neurons could help us identify the ones involved in the generation of rhythmic responses in nematodes.
C. elegans neurons in which tax-2, tax-4 and lite-1 genes are expressed.

Summary of neurons in which tax-2, tax-4 and lite-1 genes are expressed. Data were obtained using ceNGEN (C. elegans Neuronal Gene Expression Network) and then contrasted with functional and expression studies based on the literature.
Finally, some molecules (PDF and melatonin) have been identified as possible output signals of the central clock that would deliver the circadian message to the whole organism. Both systems could play an important role in C. elegans’ adaptation to its surrounding environment, allowing this organism to adequately predict the daily changes occurring in nature. The complete genome sequence and annotation for C. elegans, together with the powerful molecular genetic techniques for generating transgenic animals and analyzing gene function, promise to facilitate its genetic analysis and thus further increase the use of this model to understand key aspects of circadian clocks. Indeed, since many of the recordable outputs (behavioral, physiological and/or molecular) have been introduced rather recently, including our own bioluminescent assay, it is the right timing for a high-throughput mutant screen in order to continue the identification of the key genes and pathways underlying circadian rhythmicity in this model.
Footnotes
Acknowledgements
Studies in authors’ lab were supported by the National Science Agency (ANPCyT), the National Research Council (CONICET) and the Universidad Nacional de Quilmes, Argentina.
Conflict of Interest Statement
The author(s) have no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.