
Abstract
The present investigation was aimed at testing whether the lizard sky polarization compass is time compensated. For this purpose, ruin lizards, Podarcis sicula, were both trained and tested for orientation inside a Morris water maze under clear skies with the sun not in view. During training, lizards showed a striking bimodal orientation along the training axis, demonstrating their capability of determining the symmetry plane of the sky polarization pattern and thus the use of polarization information in orientation. After reaching criteria, lizards were kept 7 days in a 6-h fast clock-shift treatment and then released with the sun not in view. Six-hour clock-shifted lizards showed a bimodal distribution of directional choices, which was oriented perpendicularly to the training axis, as it was expected on the basis of the clock-shift. The results show that the only celestial diurnal compass mechanism that does not need a direct vision of the sun disk (i.e., the sky polarization compass) is a time-compensated compass.
Keywords
Animals can determine compass bearings either by using the azimuth of the sun or the pattern of deflected, polarized sunlight through the atmosphere. Like the sun azimuth compass, the sky polarization compass requires time compensation to take into account changes in E-vector direction and pattern rotation, as the sun changes elevation and moves horizontally across the sky (Brines and Gould, 1982). Previous investigations in the ruin lizard, Podarcis sicula, showed the use of polarized light for compass orientation and that an intact parietal eye is necessary to perform this task (Beltrami et al., 2010, 2012). Since all orientation tests were carried out indoors under a fixed source of artificial polarized light, it was impossible to decide whether the sky polarization compass is truly time compensated. To establish whether the sky polarization compass compensates for the regular daily rotation of the skylight E-vector patterns, the present investigation tested the orientation behavior of 6-h clock-shifted lizards outdoors, under clear skies, in situations where the sun disk was not in view, so that animals were prevented from using the sun azimuth compass. A pilot experiment was carried out, in which ruin lizards were individually trained to steer a single goal platform at the periphery of a hexagonal Morris water maze positioned at the geographic North. (For details, see the supplementary online material [SOM].) Training took place from 1030 to 1400 h, always with the sun disk visible. Training lasted about 20 days (2 trials per day). Nine of 20 lizards met learning criteria (scoring system in SOM and in Suppl. Fig. S1A). After reaching criteria, lizards were first tested under the sun. For each lizard, orientation tests always consisted of 1 trial in the absence of the goal platform. Lizards’ directional choices showed a unimodal distribution (Suppl. Fig. S1A), which deviated from uniform (Hodge-Ajne test, p < 0.004), and also the V test was statistically significant (u9 = 4.22, p < 0.0005). A further orientation test was carried out between 0800 and 1000 h under clear skies, with the sun not in view. At those times of day, direct vision of the sun was impaired by buildings, vegetation, and the fences surrounding the maze. In such a situation, the directional choices of lizards became bimodally distributed (Fig. 1B) and, after doubling the angles, deviated from uniform (Hodge-Ajne test, p < 0.004; V test, u9 = 2.23, p < 0.025). The striking bimodal orientation—toward/away from the goal platform—in the absence of the sun disk is explained by assuming that lizards orientate themselves by using sky polarization alone. In fact, the E-vector pattern alone does not allow discrimination between the solar and antisolar meridian, and such a 180° ambiguity imposes an axial-bimodal orientation instead of a unimodal one (Brines and Gould, 1982).
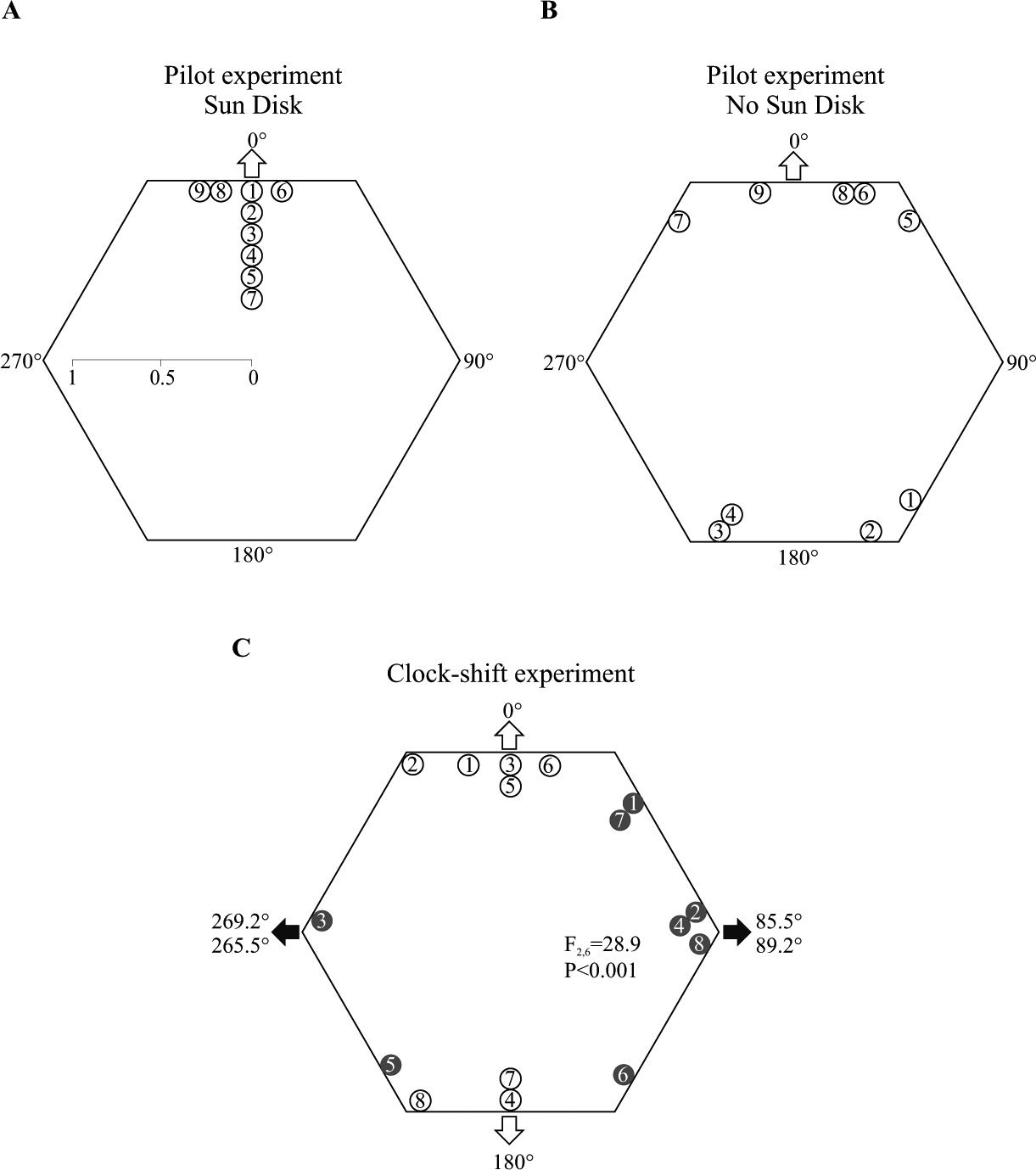
Orientation behavior of lizards. The circular diagrams plot individual directional choices of lizards. They are the traditional way of presenting orientation data and allow evaluating both the sample size and its spatial distribution. Upper diagrams report the orientation performances of lizards under the sun (A) and with the sun not in view (B) in the pilot experiment. Each empty circle indicates the directional choice of a single lizard identified by its number. The outer arrow marks the expected direction of orientation. The lower diagram (C) shows the clock-shift experiment. Each empty circle reports the directional choice of a lizard before clock-shift and each solid circle identified by the same number the directional choice of the same lizard after clock-shift. The empty outer arrows and the solid outer arrows mark the expected axis of orientation before and after clock-shift, respectively. Further information is given in the text.
In the 6-h clock-shift experiment, lizards were both trained and tested for orientation between 0830 and 1030 h with the sun not in view, a condition in which they were already shown to express an axial-bimodal orientation (Fig. 1B). Lizards were therefore subjected to axial training by positioning 2 identical goals in contact with the center of 2 opposite-side walls (0°-180°) of the maze along the geographic north-south axis. Training lasted 14 days (2 trials per day). Eight of 20 lizards met learning criteria (scoring system in SOM and Suppl. Fig. S1B). Orientation tests were carried out in absence of the goal platforms. Lizards reaching criteria at the end of their axial training were subjected to an orientation test, in which they showed, as expected, a bimodal distribution of directional choices along the training axis (Fig. 1C). After doubling the angles, the directional choices deviated from uniform (V test: u8 = 3.46, p < 0.0001). After testing, lizards were immediately housed in an environmental chamber, where the artificial day started and ended 6 h before the natural light phase (6-h fast clock-shift). Previous investigations showed that 7 days are enough to reentrain lizard orientation rhythms to a 6-h clock-shift (Foà et al., 2009). After the clock-shift treatment, the directional choices of lizards were bimodally distributed along an axis that was rotated perpendicularly with respect to the training axis (Fig. 1C). After doubling the angles, the directional choices of clock-shifted lizards deviated from uniform (V test, u8 = 1.95, p < 0.025). The directional choices of lizards before clock-shift were significantly different from the directional choices of the same lizards after clock-shift (Hotelling test for paired data: F2,6 = 28.9, p < 0.001). This difference is made clear in Figure 1C, showing that the directional choice of each lizard before clock-shift does not overlap with the directional choice of the same lizard after clock-shift. Considering the amplitude of the clock-shift (6-h phase-shift), the time of the year (from September 22 to October 17), and the geographic coordinates of Ferrara (longitude: 11°37′E, latitude: 44°49′N), we expected a deviation in axial orientation of about 85.5°-89.2°/265.5°-269.2° (Fig. 1C) (http://www.esrl.noaa.gov/gmd/grad/solcalc/azel.html). After the 6-h clock-shift, the observed deviation in lizards’ orientation was not statistically different from the expected one (one sample test for the mean angle). The axial nature of the observed orientation response does not allow distinguishing between the behavioral effects of a fast clock-shift (expected counterclockwise deflection) and those of a slow clock-shift (expected clockwise deflection). In any case, we performed a fast clock-shift and not a slow clock-shift because previous investigations clearly showed that slow clock-shift is much less effective on orientation behavior (Foà et al., 2009); this is true also in homing pigeons (Ioalè et al., 2000). The drastic changes in orientation observed after clock-shift were not due to lack of retention of the learned spatial task, since previous results in ruin lizards showed that 7 days with no training do not impair or substantially change orientation performances and directional choices (Foà et al., 2009).
A time-compensated sky polarization compass is a very high-level feature, since it requires knowledge (1) of the solar azimuth with respect to the animal, (2) of the time of day, (3) of the local ephemeris throughout the year, and (4) the capability of detecting the E-vector of polarized light to be used in orientation. The data gathered so far in P. sicula showed that this lizard has all these capabilities: (1) it is equipped with a time-compensated sun compass (Foà et al., 2009); (2) it possesses a circadian clock, allowing to exactly determine the time of day (Minutini et al., 1995); (3) it is equipped with a circannual clock to determine seasons throughout the year and hence the local ephemeris (Bertolucci et al., 1999; Foà and Bertolucci, 2003); and (4) it can use the E-vector direction of polarized light for compass orientation (Beltrami et al., 2010, 2012). The present results added further knowledge, as they showed that after clock-shift, the directional choices of lizards released with the sun not in view are not statistically different from those expected on the basis of the applied clock-shift treatment. This demonstrates for the first time that the only known celestial diurnal compass mechanism that does not need direct vision of the sun disk (i.e., the sky polarization compass) is a time-compensated compass.
Previous data in ruin lizards demonstrated that the parietal eye plays a central role in mediating functioning of both the sun compass and the sky polarization compass. In fact, lizards’ directional choices become randomly distributed outdoors, with the sun disk visible as soon as the parietal eye is surgically removed, or indoors, under a single E-vector of polarized light, as soon as the parietal eye has been painted black (Foà et al., 2009; Beltrami et al., 2010). The fact that in all experimental conditions applied so far, preventing lizards from using the parietal eye in orientation resulted in random distributions of directional choices discouraged us from further behavioral investigations. However, we retain that functional/molecular studies on the parietal eye may be more promising. Future investigations should be aimed at testing whether spatial distribution of photopigments within the parietal eye previously identified (Frigato et al., 2006) were functionally and anatomically suited to accomplish the “matched filtering” of the E-vector pattern in the sky, which was already shown to occur in ommatidia (specifically the POL area) of desert ants and honey bees (Wehner, 1989).
Footnotes
Conflict of Interest Statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.