
Abstract
High-resolution planktonic foraminiferal assemblage data from ODP Hole 846B provide new insights into Quaternary palaeoceanographic variability in the Eastern Equatorial Pacific (EEP). The record reveals that the EEP was a highly dynamic system characterised by recurrent shifts in water column structure, upwelling strength, and faunal composition in response to both tropical and high latitude climatic forcing. The repeated appearance of the temperate species Globorotalia (Globoconella) inflata identifies distinct episodes of cold water incursions into the equatorial zone, reflecting enhanced eastern boundary currents and strong teleconnections with Antarctic climate processes. The long-term dominance of Neogloboquadrina dutertrei indicates the persistent influence of the wind driven equatorial cold tongue, although its asynchronous relationship with coastal upwelling taxa points to a decoupling between equatorial and Peruvian margin upwelling systems. Variations in mixed-layer and thermocline dwellers document alternating El Niño and La Niña like conditions, tracking changes in upper ocean stratification and thermocline depth. A marked reorganisation of faunal patterns after the Mid-Pleistocene Transition (MPT) signals a fundamental shift in the dominant controls on EEP oceanography, coinciding with intensified glacial interglacial cycles. Overall, this study refines the understanding of the EEP as a climatically sensitive region governed by the complex interaction of high-latitude forcing, tropical atmospheric variability, and boundary current dynamics.
INTRODUCTION
The Equatorial Pacific Ocean, extending from the Peru–Chile coast to the Indonesian archipelago, has a dominant influence on global ocean–atmosphere interactions. It hosts several tightly linked phenomena, including oscillations in thermocline depth, the Western Pacific Warm Pool (WPWP), El Niño Southern Oscillation (ENSO) variability, the Walker circulation, and changes in high-latitude overturning circulation. These processes collectively shape the global climate by modulating heat distribution, ocean productivity, and atmospheric CO2 exchange. The Eastern Equatorial Pacific (EEP) is particularly important for understanding these linkages due to its strong upwelling, high nutrient concentrations, and persistently shallow thermocline (Wyrtki, 1966). The region is highly sensitive to both tropical and extratropical forcing, making it a key site for studying feedback between ocean circulation and climate variability (Farrell et al., 1995). Palaeoceanographic records (Bin Shaari et al., 2013; Etourneau et al., 2013; Pallone et al., 2025) from the EEP provide valuable insights into how equatorial dynamics have evolved over glacial interglacial cycles and influenced global climate through changes in ocean circulation and biogeochemical cycle, influencing the variation in productivity during the Quaternary. Calcite export fluxes during the Last Glacial Maximum (LGM) were notably lower than during the Holocene, likely reflecting reduced nutrient supply from the Southern Ocean (Loubere et al., 2004). Boron isotope based reconstructions suggest that deep-ocean pH increased during glacial periods, indicating shifts in carbonate chemistry and carbon storage, whereas surface pH remained relatively stable (Sanyal et al., 1997). Despite high macronutrient concentrations, productivity in the EEP is often limited by iron availability, underscoring the importance of nutrient dynamics and ocean–atmosphere carbon exchange in this region (Wells et al., 1999). Although the large scale physical and biogeochemical processes governing EEP variability are increasingly well understood, the detailed faunal and ecological responses to these changes remain less constrained. Understanding these relationships is crucial for interpreting how ocean structure, upwelling strength, and frontal dynamics have evolved over time.
This study examines Quaternary palaeoceanographic variability in the EEP, with a focus on the impact of the Southern Polar Front (SPF). By analysing planktic foraminiferal assemblages, species composition, and abundance patterns from ODP Hole 846B (Figure 1), we aim to reconstruct past surface and thermocline conditions and assess how equatorial ocean dynamics responded to glacial interglacial forcing.
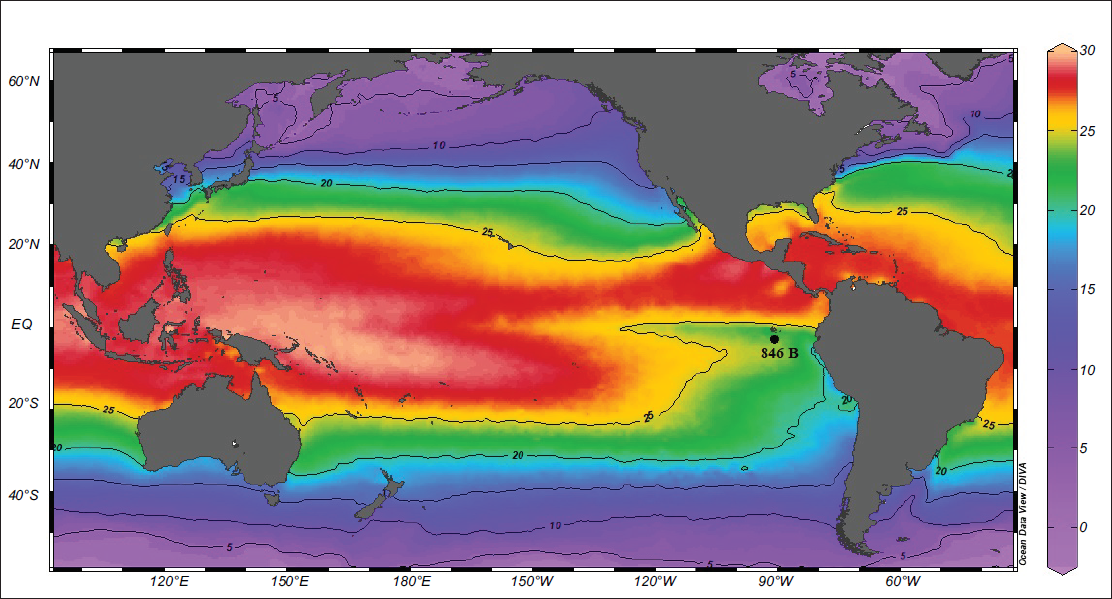
Modern annual Sea Surface Temperature (°C) from the World Ocean Atlas 2023 (Reagan et al., 2024), visualised using Ocean Data View (Schlitzer, 2023) with ODP site 846B (black dot).
MODERN OCEANOGRAPHIC SETUP
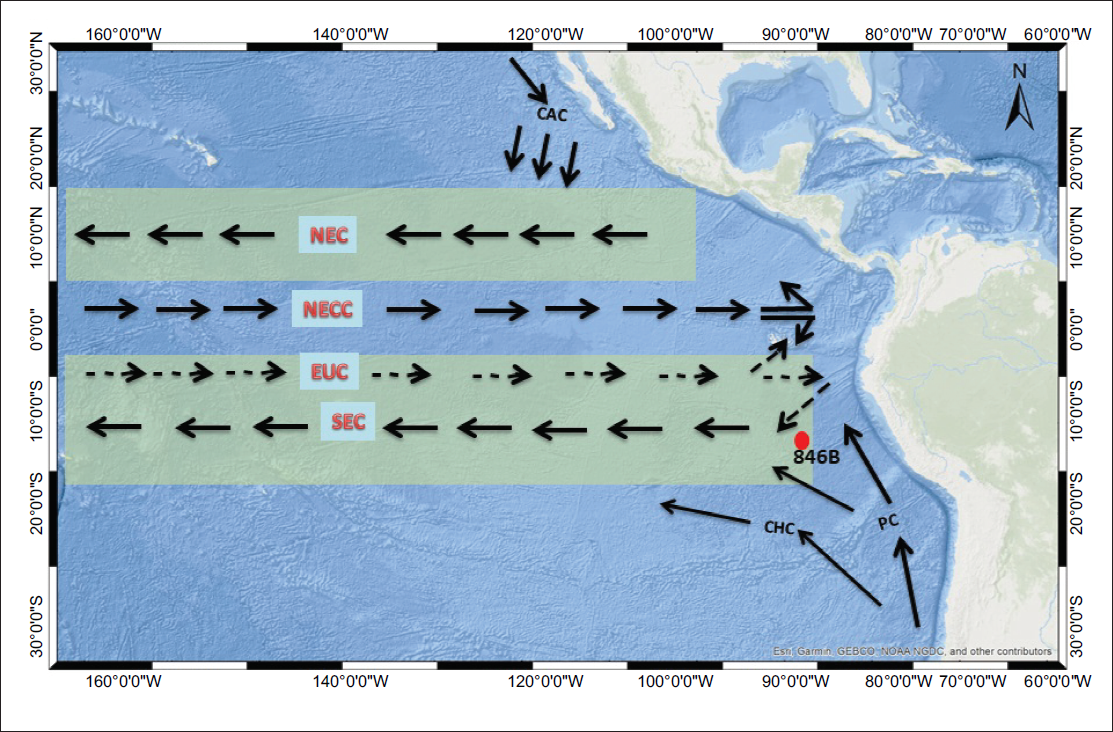
In the trans-tropical Pacific, oceanic circulation is largely influenced by tropical atmospheric dynamics and changes in the sign of the Coriolis force across the equator. A defining feature of this circulation is the asymmetry of surface currents on either side of the equator, which corresponds to the mean latitudinal position of the Intertropical Convergence Zone (ITCZ), the region where the northeast and southeast trade winds converge. Along the equator, wind driven divergence depresses the sea surface topography, generating a pressure gradient that drives a westward geostrophic flow on both sides of the equator, known as the South Equatorial Current (SEC). The primary source waters for the SEC are derived from the Peru Current (PC), which transports cold, nutrient rich waters from higher latitudes, and from the eastward flowing subsurface Equatorial Undercurrent (EUC), which contributes to surface waters through upwelling (Mayer et al., 1992).
Site 846B (3°5.696′S, 90°49.078′W) was drilled during Ocean Drilling Programme (ODP) Leg 138 at a water depth of 3,307 m in the EEP. The site lies approximately 300 km south of the Galápagos Islands, within a zone where the SEC interacts with the PC (Figure 2).
Location of ODP Site 846B and present-day oceanographic setup of the Eastern Equatorial Pacific Ocean. Surface current shown as solid arrows; subsurface current as dashed arrows. CAC (California Current); NEC (North Equatorial Current); NECC (North Equatorial Counter current); EUC (Equatorial Undercurrent); SEC (South Equatorial Current); PC (Peru Current). Shaded areas illustrate the general latitudinal extent of the SEC and NEC. (Adopted and modified from Mayer, L., Pisias, N., Janecek, T., et al., 1992 Proceedings of the Ocean Drilling Program, Initial Reports, Vol. 138).
At present, the EEP ‘cold tongue’ functions as a major ocean to atmosphere CO2 source (Takahashi et al., 2009), sustained by the persistent upwelling of cool, nutrient rich subsurface waters supplied primarily by the EUC (Fiedler & Talley, 2006; Kessler, 2006; Toggweiler et al., 1991). These waters originate largely in the Southern Ocean’s Subantarctic Zone, linking high latitude nutrient cycling to low latitude productivity (Rodgers et al., 2003; Toggweiler et al., 1991). Despite elevated macronutrient concentrations, primary productivity in the EEP is constrained by iron and silicic acid limitation, resulting in relatively low phytoplankton biomass even under high nutrient supply (Brzezinski et al., 2011; Dugdale et al., 1995; Quirós-Collazos et al., 2020). The site 846 lies on the southern flank of the Carnegie Ridge, underlain by oceanic crust generated at the Galápagos Spreading Centre. Its palaeopositions have been quantitatively constrained using Pacific–Nazca plate rotation models in conjunction with Pacific hotspot reference frames. Paleogeographic reconstructions indicate that Site 846 has remained within a narrow latitudinal band near its present position throughout much of its geological history (Farrell et al., 1995). This stability makes it a key site for evaluating the temporal evolution of equatorial current systems and large scale ocean circulation patterns.
MATERIALS AND METHODS
The studied core reveals alternating layers of carbonate-rich and silica-rich sediments. Deep sea samples were collected from a 90 meter long core at 30 centimeter intervals. Each sample was processed using the wet sieving method following international standard protocols. Planktic foraminifera retained on the ≥150 µm sieve were studied following Peeters et al. (1999). The samples were weighed and stored in labelled tubes. These specimens were then split to obtain aliquots containing at least 300 individuals, following the methods of Imbrie and Kipp (1971) and Imbrie et al. (1973), and were mounted on assemblage slides. The foraminiferal species were classified based on their water mass and depth habitat preferences within the ocean water column. Taxonomic identification to the species level followed the frameworks of Kennett and Srinivasan (1983), Bolli and Saunders (1985), and Schiebel and Hemleben (2017).
Age Model
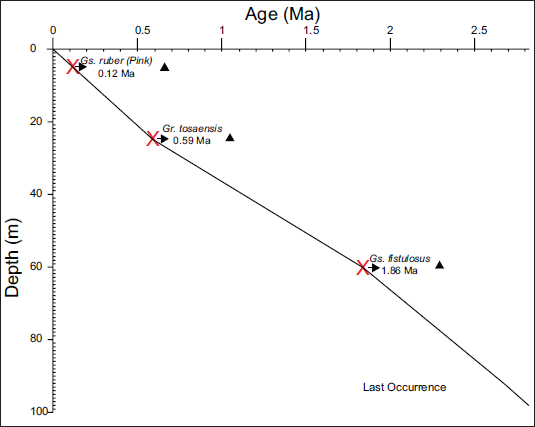
Sediment accumulation rates in pelagic settings are primarily controlled by carbonate productivity (Berger & Winterer, 2009). Shackleton et al. (1992) and the Shipboard Scientific Party (Mayer et al., 1992) developed a consistent method to calculate sedimentation rates across multiple drill sites from ODP Leg 138, using a unified stratigraphic framework based on Gamma Ray Attenuation Porosity Evaluator (GRAPE) density measurements. Key stratigraphic events were selected for their close association with well-dated magnetic reversals and biostratigraphic datums, ensuring accurate age control. At ODP Site 846B, sediment accumulation has remained relatively uniform throughout the Quaternary (Figure 3). In the present study, an age model was constructed using established phylogenetic knowledge to assess the reliability of planktonic foraminiferal events.
Nearly uniform sediment accumulation rate at ODP site 846 B.
RESULTS
In this study, 266 ODP samples were analysed. The assemblage is dominated by planktonic foraminiferal species such as Neogloboquadrina dutertrei (d’Orbigny, 1839), Globigerina bulloides (d’Orbigny, 1826), Globigerinoides ruber (d’Orbigny, 1839), Globigerinita glutinata (Egger, 1893), Orbulina universa (d’Orbigny, 1839), Globorotalia menardii (d’Orbigny, 1826), Globorotalia tumida (Brady, 1877), and Neogloboquadrina pachyderma (Ehrenberg, 1862), which together account for nearly 75% of the total population.
For palaeoceanographic reconstruction during the Quaternary, the planktonic foraminifera were categorised by ecological habitat into three groups: Mixed Layer Dwellers (MLD), dominated by G. ruber; Thermocline Dwellers (TD), dominated by N. dutertrei; and Upwelling Indicators (UI), dominated by G. bulloides. The relationships among key species within these groups form the basis for the interpretations presented in this study. These ecological groups were selected based on earlier studies by Kennett and Srinivasan (1983), Keller (1985), Chaisson and Ravelo (2000), and Aze et al. (2011).
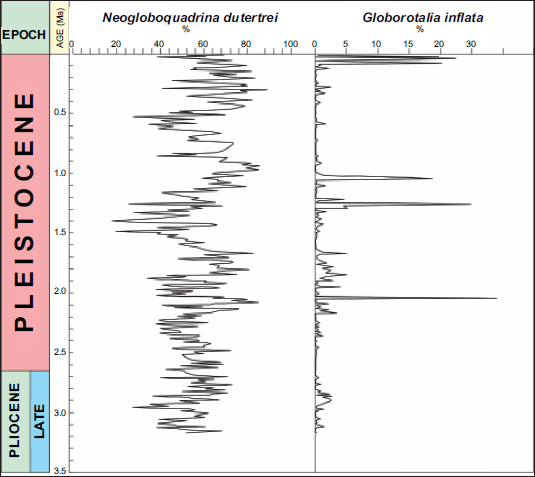
Relative abundance of Neogloboquadrina dutertrei (Thermocline Dweller) and Globorotalia (Globoconella) inflata (Temperate Group)
According to Loubere (2001), in the EEP, N. dutertrei inhabits the thermocline zone, typically between 40 and 80 metres in depth, occupying a stable position along the temperature gradient from the surface to about 100 metres. Its oxygen isotope values indicate equilibrium with waters in the lower part of this profile. In contrast, carbon isotope data show that it reflects regional nutrient gradients below 50 metres, confirming its preference for nutrient rich thermocline waters (Loubere, 2001). The species maintained a consistent depth habitat both under modern conditions and during the LGM, demonstrating ecological stability despite climatic fluctuations. Regional variations in its isotopic composition further suggest that N. dutertrei is sensitive to local changes in thermocline structure and nutrient supply rather than to large scale global water mass shifts. Plankton tow and sediment trap data from the Panama Basin further indicate that N. dutertrei primarily resides within the thermocline, with its abundance increasing when the thermocline shoals, reflecting favourable conditions for growth (Mix et al., 1995; Thunell & Reynolds, 1984). N. dutertrei is also classified by Aze et al. (2011) within Eco-Group 3, representing taxa characteristic of the open ocean thermocline.
Relative abundance of N. dutertrei and temperate fauna in sediments from ODP Site 846B. The left panel shows the percentage relative abundance of N. dutertrei (black line) with fluctuations between ~20% and 90%. In contrast, the right panel shows the relative abundance of temperate fauna, dominated by G. inflata during four distinct intervals correlating with estimated glacial periods at 2.1 Ma, 1.2 Ma, 1.0 Ma, and the Late Holocene. Age model and stratigraphic calibration follow Gradstein et al. (2020). This pattern indicates episodic cold-water incursions, likely linked to glacial intensifications in the Southern Hemisphere.
Stratified oceanic systems frequently develop a Deep Chlorophyll Maximum (DCM) that remains undetectable by ocean colour satellites. The DCM may correspond to either an actual phytoplankton biomass peak, termed the Deep Biomass Maximum (DBM), or a physiological adaptation to low-light conditions, in which phytoplankton increase their chlorophyll a content relative to carbon, forming a Deep Photoacclimation Maximum (DAM) (Cornec et al., 2021). Several studies have also reported associations between specific zooplankton taxa and the DCM (Fairbanks & Wiebe, 1980). The consistent downcore presence of N. dutertrei at ODP Hole 846B indicates tropical to subtropical subsurface conditions linked to the DCM, positioned near the open ocean thermocline (Schiebel & Hemleben, 2017).
Globorotalia inflata is a temperate species, and its incursion into tropical latitudes has been interpreted as evidence of strengthened cold water advection associated with the northward migration of the Antarctic Polar Front (APF) (Singh & Sinha, 2021; Sinha & Singh, 2007a, 2007b). Although Aze et al. (2011) classified G. inflata within the same ecological group (Eco-Group 3) as N. dutertrei, the former is typically restricted to temperate and subpolar water masses. It is only rarely recorded in modern equatorial regions. Episodic increases in G. inflata abundance during the Quaternary have been documented from the Eastern Indian Ocean (Sinha et al., 2006), where three distinct PL events (PL-1, PL-2, PL-3) were identified and linked to phases of intensified West Australian Current activity driven by the northward shift of the APF (Singh & Sinha, 2021). Le et al. (1995) reported that G. inflata exhibits several distinct abundance spikes at oxygen isotope stage boundaries 3/2, 4/3, 17/16, and 18/17 at ODP Site 846. The species was also notably abundant during MIS 2. Similar patterns have been observed in other cores (ODP Site 847) from the EEP Ocean (McKenna et al., 1995). Le et al. (1995) suggested that the factors driving these sudden abundance spikes during certain glacial interglacial transitions require further investigation. Both studies noted an abundance of G. inflata during glacial stages and during MIS 19 (interglacial stage).
During the Early Pleistocene (2.6–1.0 Ma), N. dutertrei abundance fluctuates between ~20% and 70%, suggesting alternating periods of thermocline strengthening and weakening, likely tied to orbital-scale climate oscillations, ENSO variations and the onset of Northern Hemisphere glaciation. In the Late Pleistocene (<1 Ma), the record remains variable. However, it tends to have overall lower mean values, reflecting greater vertical mixing and cooler surface conditions, consistent with stronger glacial interglacial contrasts.
G. inflata is nearly absent before ~2.5 Ma, implying that temperate or subpolar influences were minimal in the tropical Pacific during the warm Pliocene. After ~2.5 Ma (near the Pliocene–Pleistocene boundary), intermittent but distinct peaks appear, some exceeding 20%. These peaks correspond to cooler intervals or enhanced upwelling episodes, during which subtropical or temperate waters penetrate the equatorial region. Its increased occurrence in the Pleistocene marks enhanced latitudinal temperature gradients and a more dynamic ocean circulation regime.
The overall pattern reflects a long-term cooling trend and increasing climate variability from the Late Pliocene into the Pleistocene. The dominance of N. dutertrei in the Pliocene suggests warm, stratified surface waters, characteristic of a strong equatorial thermocline and reduced upwelling. The emergence and sporadic increases of G. inflata after ~2.5 million years (Ma) indicate a strengthening of upwelling, shoaling of the thermocline, and an increased incursion of cooler waters, consistent with the intensification of the Walker circulation and the development of modern equatorial upwelling systems.
Periods of increased N. dutertrei abundance are occasionally accompanied by a temporary appearance of the temperate species G. inflata. However, during the Late Holocene, N. dutertrei shows a marked decline, while G. inflata exhibits an abrupt increase.
The anomalous occurrences of G. inflata reaching relative abundances of about 20% or higher are observed during four distinct intervals at approximately 2.1 Ma, 1.2 Ma, 1.0 Ma, and the Late Holocene. These episodes likely reflect intensified glacial conditions that facilitated the southward penetration of cold, subpolar waters.
Variations in relative abundance over time highlight the dominance of N. dutertrei and the intermittent invasions of G. inflata at 3°S, which correspond to periods of enhanced glacial influence (Figure 4).
Relative abundance of Mixed Layer Dwellers and Thermocline Dwellers
The MLD group, primarily composed of Globigerinoides species dominated by G. ruber, maintains an average relative abundance of about 10%, reaching a maximum of approximately 37% at three distinct intervals downcore (Figure 5). An analysis of marine dynamics from the mid-Pleistocene to the Late Holocene reveals pronounced fluctuations in both the MLD and TD (Figure 5), marked by alternating dominance in their relative abundances. Around 0.57 million years ago (Ma), the MLD population reached 25.88%, indicating a gradual decline throughout the middle Pleistocene and continuing into the Late Holocene. In contrast, at approximately 27 kyr, the TD attained its peak relative abundance of about 35%, suggesting a shoaling of the thermocline. These variations reflect changes in ocean stratification, likely driven by global temperature shifts and alterations in oceanic circulation.
Relative abundance of Mixed Layer Dwellers (MLD, left panel) and Thermocline Dwellers (TD, right panel) recorded in ODP Site 846B sediment core. Corresponding age (Ma) is shown. Notable intervals of elevated MLD abundance coincide with declines in TD, indicating dynamic stratification and El Niño like conditions during the Quaternary. Stratigraphic context follows Gradstein et al. (2020). Mixed Layer Census data taken from Mallick et al. (2017).
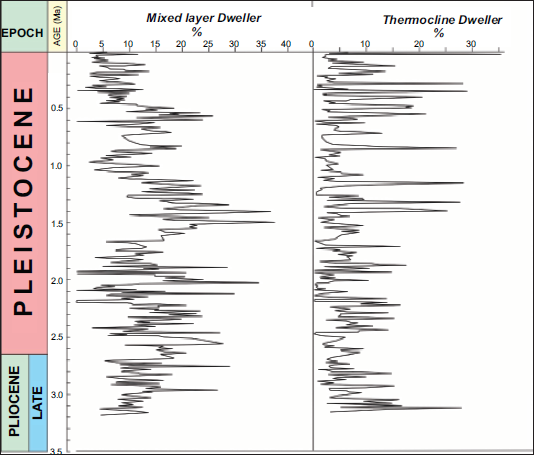
Six distinct intervals were observed at approximately 2.15 Ma, 2.0 Ma, 1.86 Ma, 1.47 Ma, 1.44 Ma, and 0.57 Ma, showing remarkable increases in the relative abundance of MLD, accompanied by corresponding declines in TD (Figure 5). This inverse relationship suggests El Niño like conditions in the EEP. The species interrelationships and their stratigraphic patterns support this interpretation.
Around 0.5 Ma, MLD reached a peak of ~25.9%, followed by a gradual decline into the Late Pleistocene and Holocene. In contrast, in the youngest part of the core, TD reached its highest relative abundance (~35%), suggesting significant reorganisation of the upper water column during this time.
The observed inverse relationship between MLD and TD abundances reflects alternating phases of surface water stratification and thermocline structure in the EEP. Periods dominated by MLD imply enhanced surface water productivity and a deepened thermocline, conditions often associated with El Niño like states. Conversely, intervals of increased TD abundance indicate a shoaling thermocline, characteristic of intensified upwelling and La Niña like conditions. These long-term oscillations correspond to major climatic transitions through the Pleistocene, including shifts in global ice volume and oceanic circulation that modulated the strength and depth of the EEP thermocline. The parallel trends between species groups and stratigraphic intervals therefore record a dynamic interplay between tropical surface water variability and global climate forcing.
Relative abundance of Upwelling Indicator Species (UIS), Mixed Layer Dwellers (MLD), and Neogloboquadrina dutertrei
The figure compares the relative abundances (percentage) of UIS, MLD, and N. dutertrei across the Quaternary (Figure 6), based on core depth/age. UIS show three pronounced intervals of increased abundance: near 2.46 Ma (max ~46%), 1.48 Ma, and 0.2 Ma. The highest UIS value corresponds to a significant decline in MLD, indicating strong upwelling and an enhanced nutrient supply at these times (Figure 6). MLD display an inverse pattern, with lower relative abundance during periods of intense upwelling (notably at 2.46 Ma and 0.2 Ma), and relatively higher values between these upwelling events. N. dutertrei dominates the planktonic foraminifera assemblage throughout the record, with relative abundance fluctuating between approximately 17.7% and 87% (Figure 6). Its maxima tend to coincide with upwelling peaks. The transitions from the Late Pliocene into the Pleistocene (~2.6 Ma boundary) are marked by significant faunal shifts, particularly involving increases in UIS and N. dutertrei.
Relative abundance profiles of Upwelling Indicator Species (UIS), Mixed layer Dweller and N. dutertrei from ODP Site 846B. and corresponding age (Ma). Major UIS peaks are synchronous with reduced MLD abundance, indicating amplified upwelling and nutrient delivery. Mixed Layer Census data taken from Mallick et al. (2017).
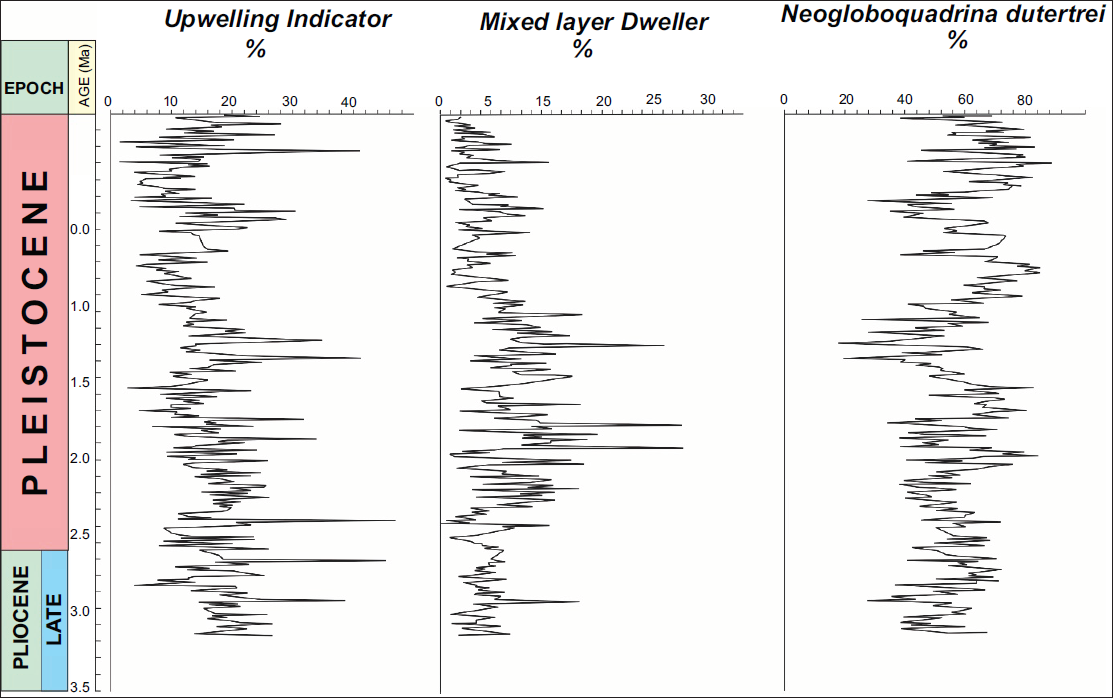
Peaks in UIS percentage are interpreted as episodes of intensified upwelling, likely linked to strengthened trade winds or reorganisations in oceanographic circulation, possibly influenced by major climatic transitions. The marked decrease in MLD during these upwelling intervals indicates increased environmental stress and nutrient enrichment in surface waters, favouring more opportunistic, upwelling adapted taxa. The consistently high abundance of N. dutertrei supports its role as a thermocline or lower MLD, benefiting from the nutrient rich, cooler waters supplied by upwelling. Its dominance underscores the prevalence of Neogloboquadrina assemblages characteristic of the EEP cold tongue. The temporal offsets between UIS and MLD can be interpreted as evidence for alternating dominance of upwelling versus stratified water column conditions, reflecting both global climatic cycles (ice volume, trade winds) and regional oceanographic processes (Peruvian Current versus EUC). The data support a model in which increased N. dutertrei abundance and UIS peaks represent responses to nutrient enrichment resulting from the combined effects of coastal upwelling off Peru and equatorial upwelling generated by the EUC, as described by Keigwin (1976). Such patterns may delineate intervals when the equatorial and coastal upwelling systems were more or less coupled, thus influencing the spatial extent and continuity of the equatorial cold tongue. The lower UIS and relatively higher MLD values during some periods suggest phases of weakened upwelling and/or strengthened stratification.
The comparison thus provides key evidence for the variability and drivers of upwelling in the eastern equatorial during the Quaternary, linking palaeoproductivity changes to global and regional oceanographic controls. The three intervals of maximum UI correspond to major palaeoceanographic reorganisations and can be related to both global climate shifts and changes in regional ocean–atmosphere dynamics.
DISCUSSION
Palaeoceanographic variability in the Eastern Equatorial Pacific
Analysis of planktonic foraminiferal assemblages from ODP Hole 846B documents a dynamic palaeoceanographic evolution of the EEP throughout the Quaternary. The record reveals pronounced fluctuations in water-column structure, upwelling intensity, and intermittent incursions of temperate water masses. When viewed alongside regional and global climate reconstructions, these results highlight the EEP’s sensitivity to both high latitude polar forcing and low latitude atmosphere–ocean interactions.
The EUC serves as the principal source of upwelled water in the eastern Pacific cold tongue (Wyrtki & Kilonsky, 1984). Cromwell (1953) demonstrated that equatorial upwelling arises from the divergence of Ekman transport near the equator. This upwelled water mass, enriched in iron from sedimentary sources along the Papua New Guinea shelf (Ryan et al., 2006), sustains high primary productivity and contributes to deep-ocean CO2 sequestration. Lateral advection from the upper thermocline (Wyrtki, 1981) is consistent with the high relative abundance of N. dutertrei observed in this record, a species characteristic of the upper thermocline environment.
Incursions of temperate fauna and high latitude Linkages
A notable feature of the record is the occurrence of four intervals with anomalously high abundances (≥20%) of the temperate species Globorotalia (Globoconella) inflata at approximately 2.1 Ma, 1.2 Ma, 1.0 Ma, and in the Late Holocene. These events align with the equatorial incursions of G. inflata previously reported by Feldberg and Mix (2003), which were attributed to the intensification of eastern boundary currents in the southeast Pacific. The appearance of this temperate taxon at a near equatorial site (3°S) implies substantial equatorward transport of cold, subpolar waters (Figure 7).
The main surface flow paths, indicated by a solid, thick blue arrow, illustrate the intensified Humboldt Current, which has resulted from the northward migration of the Southern Polar Front driven by the expansion of the Antarctic Ice Sheet (AIS). This phenomenon facilitated the unusual invasion of temperate fauna, G. inflata, at the studied ODP 846B at 3˚S latitude. Principal surface flow paths are: PCC (Peru Chile Current), Peru Coastal Current (PCoastalC), Chile Coastal Current (CCoastalC), Peru Chile Counter Current (PCCC), Equatorial Undercurrent (EUC), modified after Karstensen & Ulloa, 2009.
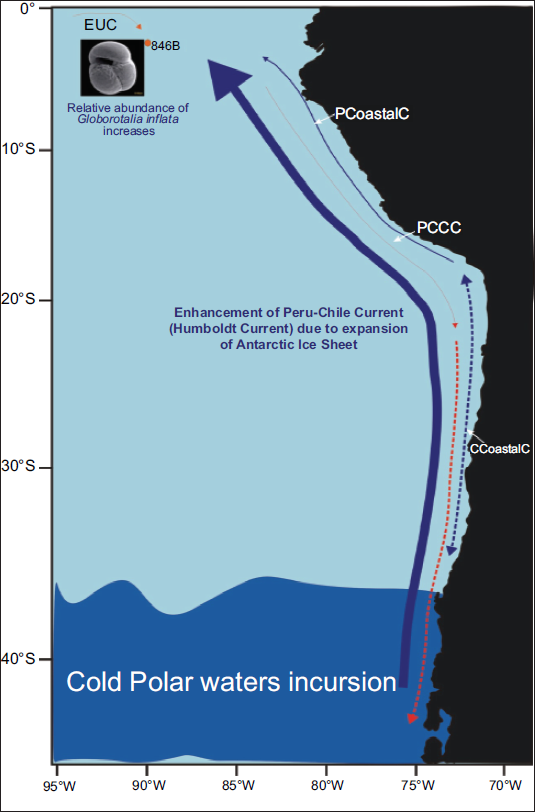
Comparable high latitude linkages have been documented elsewhere in the Southern Hemisphere. Singh and Sinha (2021) identified three major northward migrations of the APF in the southeast Indian Ocean, including one that occurred around 1.2 million years ago (Ma), which coincides with the present event. They associated the proliferation of the Globoconella group with the strengthening of the cold West Australian Current driven by Antarctic ice-sheet expansion. Similarly, reconstructions from the South Tasman Rise indicate that during glacial intervals, the Subtropical Front (STF) shifted northward by up to 5° of latitude, lowering regional temperatures by about 4°C. During the LGM, the frontal system advanced so far north that it became constrained against the Tasmanian landmass (Sikes et al., 2009).
In the EEP, these temperate incursions likely reflect enhanced activity of the Peru–Chile (Humboldt) Current, which advects cold, subpolar water northward (Figure 7). During cooler periods such as the LGM, intensification of this current extended the range of subpolar species along the eastern boundary (Feldberg & Mix, 2003). The synchronous occurrence of temperate fauna in both the Indian and Pacific Oceans suggests a hemisphere-wide oceanographic response to glacial expansion, characterised by stronger eastern boundary currents and increased inter-ocean connectivity (Singh & Sinha, 2021; Sinha & Singh, 2007a, 2007b).
Decoupling of upwelling dynamics
During glacial intervals, particularly the LGM, the EEP experienced pronounced sea-surface cooling, with SSTs decreasing by 5°C–7°C along the equator (Feldberg & Mix, 2003). This was accompanied by a steeper meridional SST gradient south of the equatorial front (Martínez et al., 2003). In the present record, this period is characterised by thermocline shoaling, evidenced by a 35% peak in thermocline dwelling foraminifera, and intensified trade winds, reflected by the dominance of N. dutertrei (Figure 8).
The intervals of high relative abundance of N. dutertrei at the studied site are due to persistent nutrient supply from the EUC, along with advected water from the Peruvian Margin Upwelling system. Principal surface flow paths are: PCC (Peru Chile Current), Peru Coastal Current (PCoastalC), Chile Coastal Current (CCoastalC), Peru Chile Counter Current (PCCC). Equatorial Undercurrent (EUC), modified after Karstensen & Ulloa, 2009.
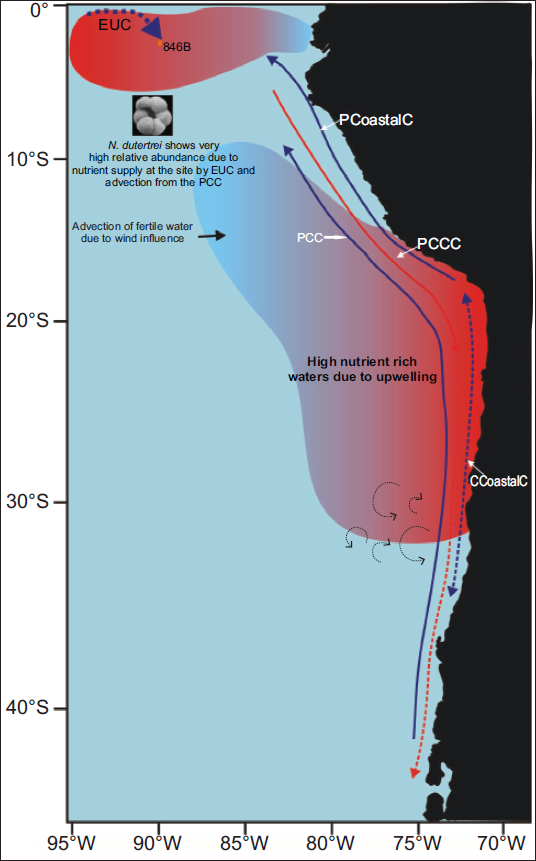
The foraminiferal assemblages indicate that two partially decoupled systems maintained EEP upwelling:
Coastal upwelling along the Peruvian margin, dominated by opportunistic taxa such as G. bulloides and G. glutinata; and Equatorial upwelling, driven by trade-wind-induced divergence and sustained by nutrient rich waters of the EUC, is represented chiefly by N. dutertrei.
Periods of enhanced coastal upwelling did not always coincide with peaks in N. dutertrei, suggesting asynchronous behaviour between the two systems. The highest relative abundance of UIS at 2.46 Ma (46%) corresponds to a reduction in MLD but not to N. dutertrei maxima. This supports a dual-mechanism model of productivity in the EEP: one modulated by high latitude forcing through advective nutrient input from the Peru–Chile Current, and the other governed by local atmospheric forcing through trade-wind variability (Figure 8).
Feldberg and Mix (2003) similarly distinguished between high latitude advective and low latitude upwelling components of SST variability. The faunal decoupling observed here provides independent evidence for these distinct yet interacting drivers of regional productivity. This dynamic interplay may explain why the EEP maintained high productivity even during warm phases, challenging simplified models of tropical ocean dynamics.
Water column structure and climatic implications
Variations in the relative abundances of MLD and TD indicate significant reorganisation of upper ocean stratification through time. Six major intervals approximately 2.15 Ma, 2.0 Ma, 1.86 Ma, 1.47 Ma, 1.44 Ma, and 0.5 Ma—show sharp increases in MLD coupled with declines in TD, interpreted as El Niño like conditions with a deepened thermocline and reduced vertical stratification. Conversely, the TD maximum around 27 kyr suggests thermocline shoaling and stronger upwelling, consistent with La Niña like states.
A long-term decline in MLD abundance after 0.5 million years (Ma), accompanied by an increase in TD, signifies a fundamental shift in the EEP ocean structure. This transition may reflect weakening of the trade winds and reduction of the equatorial cold tongue, coinciding with the Mid-Pleistocene Transition (MPT), when glacial interglacial cycles became more intense and climatic variability amplified globally.
CONCLUSIONS
By integrating detailed foraminiferal assemblage data with broader palaeoceanographic evidence, this study provides a high-resolution biological perspective that refines large-scale physical and geochemical interpretations of the EEP during the Quaternary. The record from ODP Hole 846B demonstrates that the EEP was far from static; it experienced significant reorganisations in water-column structure, upwelling strength, and faunal composition driven by both regional and global climate processes.
The repeated presence of Globorotalia (Globoconella) inflata marks distinct episodes of cold water incursions into the equatorial Pacific, linked to strengthened eastern boundary currents during glacial intervals and high latitude forcing.
The dominance of N. dutertrei reflects the long-term persistence of a wind driven equatorial cold tongue, while its asynchronous relationship with coastal UI supports a dual-control model of EEP productivity.
Variations in mixed layer and TD record oscillations between El Niño– and La Niña like conditions, indicating shifts in upper ocean stratification and thermocline depth.
The faunal reorganisation following the MPT signifies a major restructuring of EEP circulation and upwelling systems in response to intensified glacial interglacial cycles.
Overall, the EEP emerges as a climatically sensitive region shaped by an intricate interplay of high-latitude processes, tropical atmospheric dynamics, and boundary current variability.
Footnotes
Acknowledgement
KRM, PKN, and BM acknowledge the infrastructural support provided by the Department of Geology. AKS and DKS express their gratitude to both the Delhi School of Climate Change and Sustainability and the Department of Geology, University of Delhi, for their infrastructural assistance. The research problem was conceptualised by KRM and AKS, who also prepared the manuscript. DKS contributed to drafting the manuscript and provided valuable feedback. Planktic foraminiferal census analyses were performed by PKN and KRM, with technical support from BM.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: The authors gratefully acknowledge funding support provided under the Palaeoclimate Programme of the Ministry of Earth Sciences, Government of India, for carrying out the research, as well as for the preparation and publication of this manuscript (Sanction No. MoES/CCR/Paleo-4/2019).