
Abstract
Planktonic foraminifera exhibit species-specific responses to temperature, salinity, nutrient supply, and carbonate chemistry, making them sensitive recorders of past ocean conditions. This study investigates the late Pleistocene calcification and dissolution patterns in planktonic foraminifera using two deep-sea sediment cores (BC 37, water depth: 4,252 m; SVBC 37, water depth: 3,992 m) from the Central Indian Basin (CIB). We analysed shell weight, test size, and scanning electron microscopic (SEM) evidence in three species with varying susceptibility to dissolution, namely Globorotalia menardii, Pulleniatina obliquiloculata, and Neogloboquadrina dutertrei. The results reveal species-specific calcification responses to glacial-interglacial climate forcing and a strong post-depositional overprint driven by deep-water carbonate saturation. Notably, enhanced shell weights during glacial stages in G. menardii and N. dutertrei indicate improved preservation associated with higher concentrations of carbonate ions. At the same time, the SEM image confirms progressive test degradation during dissolution-prone intervals, such as Marine Isotope Stage (MIS) 11. Weak correlations between shell weight and size underscore the influence of diagenesis and test wall thinning near the lysocline. Variations in the lysocline and carbonate compensation depth (CCD), together with changes in monsoon-driven productivity, further modulate preservation patterns. These findings highlight the complex interplay between calcification, dissolution, and carbonate chemistry across climate cycles, providing valuable insights into biogenic carbonate dynamics and their implications for both past and future ocean acidification scenarios.
Keywords
INTRODUCTION
Planktonic foraminifera are unicellular marine protists that precipitate calcareous shells, playing a key role in marine biogeochemical cycles (Schiebel, 2002). They contribute significantly to the oceanic carbonate budget, accounting for nearly half of the total CaCO3 production in the open ocean (Honjo et al., 1995; Schiebel, 2002). The calcareous shells of planktonic foraminifera, once deposited and preserved in marine sediments, serve as powerful archives of past oceanic and climatic changes, facilitating reconstructions of sea surface temperature (SST), productivity, and carbonate chemistry (Gebregiorgis et al., 2016; Schmidt et al., 2004). Due to their ecological sensitivity and widespread distribution across diverse oceanic regions, planktonic foraminifera are extensively used as proxies in palaeoceanographic reconstructions (Gayathri et al., 2022; Kucera, 2007; Rebotim et al., 2017). Moreover, by contributing to both the biological pump (via organic carbon export) and the carbonate pump (through the production of calcareous shells), planktonic foraminifera play a crucial role in regulating the global carbon cycle and mediating the exchange of CO2 between the ocean and the atmosphere (Ravelo & Hillaire-Marcel, 2007). These dual functions underline their importance not only in palaeoclimate reconstruction but also in understanding feedback mechanisms within the Earth system.
The processes of foraminiferal calcification and post-depositional dissolution can be effectively assessed through an integrated approach combining morphometric and geochemical analyses. Morphometric methods involve measurements of shell weight, size, and density, which serve as proxies for calcification intensity and preservation state (Beer et al., 2010). Shell weight and size are established indicators of calcification intensity and are influenced by seawater carbonate ion concentration (CO32−), saturation state, and temperature (Beer et al., 2010; de Villiers, 2004). Recent advancements enable the reconstruction of original shell weights, taking into account carbonate loss due to dissolution (Qin et al., 2020, 2023). Qin et al. (2020) showed that Pulleniatina obliquiloculata shell weight correlates with SST, and Qin et al. (2023) found that Neogloboquadrina dutertrei shell weight reflects deep-water carbonate ion concentration after adjusting for dissolution effects. Additionally, test size in species like Globorotalia menardii serves as a proxy for ecological stress and upper-ocean stratification, with larger specimens typically indicating more favourable surface-water conditions (Thunell et al., 1983). Scanning electron microscopy (SEM) offers high-resolution visual evidence of foraminiferal test preservation, allowing the identification of microstructural features indicative of dissolution, such as surface etching, wall thinning, pitting, and fragmentation (Berger, 1970; Fehrenbacher et al., 2017). These microscopic alterations provide qualitative support to morphometric and geochemical indicators, helping to distinguish between primary calcification signals and post-depositional alteration.
Numerous studies have investigated carbonate dissolution using various faunal groups, individual species, and geochemical proxies. However, a majority of the research focusing on carbonate dissolution during the Quaternary period has relied predominantly on foraminiferal dissolution indicators (Berger, 1968, 1970; Droxler & Cullen, 1990; Kawagata et al., 2006; Mekik & Raterink, 2008; Murray & Prell, 1991; Nath et al., 2013; Oba, 1969; Peterson & Prell, 1985; Rai et al., 2011; Sijinkumar et al., 2022; Wang et al., 2021). While traditional dissolution indices based on species abundance and preservation state are widely used, integrating individual shell weight and size measurements with established proxies offers a more comprehensive approach to understanding foraminiferal dissolution processes (Beer et al., 2010; Qin et al., 2023; Thunell & Honjo, 1981). Shell weight analysis of planktonic foraminifera such as Trilobatus sacculifer, Globigerinoides ruber, and P. obliquiloculata has been effectively used to infer surface-water carbonate ion concentration and assess dissolution intensity in the Arabian Sea and western Indian Ocean, providing a robust proxy for reconstructing past carbonate chemistry and preservation conditions (Godad et al., 2017; Naik et al., 2010; Yadav et al., 2022).
SITE LOCATION AND OCEANOGRAPHIC SETTINGS
The Central Indian Basin (CIB) encompasses a diverse range of sediment types, including terrigenous-siliceous, calcareous, siliceous, and red clay deposits (Nath & Mudholkar, 1989; Purnachandra Rao & Nagender Nath, 1988). The basin’s bathymetry is marked by distinct topographic variation (Figure 1). High-relief features dominate the western sector, the eastern part exhibits moderate-relief structures, and the central region consists mainly of extensive planar formations (Kodagali et al., 1992). Geographically, the CIB is bounded by the Central Indian Ridge (CIR) to the west and the Ninetyeast Ridge (NER) to the east, with the latter serving as a prominent physiographic barrier separating it from the deeper Wharton Basin (Sclater & Fisher, 1974). Sediment input into the basin is primarily sourced from the Ganges-Brahmaputra river system, with turbidity currents acting as the main transport mechanism (Nath et al., 1989). The CIB has emerged as a key region for palaeoceanographic studies owing to its sensitivity to both tropical surface-ocean dynamics and deep-water carbonate chemistry. However, carbonate-rich sediments are rare across the basin, which poses a challenge to such investigations. Sediments from seamount tops, which often preserve carbonate materials better, provide a valuable alternative for reconstructing past oceanographic conditions (Nath et al., 2013). Positioned at the convergence of equatorial and subtropical water masses, the basin is strongly influenced by the Indian Monsoon Current and incursions of Antarctic Bottom Water (AABW), which together modulate biogenic productivity, sedimentation rates, and carbonate preservation (De Sousa et al., 2001; Nath et al., 2013). The CIR also acts as a hydrodynamic barrier, restricting the inflow of bottom-water currents from the Madagascar and Crozet basins. Previous studies have reported significant spatial variability in the lysocline and carbonate compensation depth (CCD) across the region. Kolla et al. (1976) identified the lysocline and carbonate critical depth (CCrD) at approximately 4,000 m and 5,050 m, respectively, along a transect from 10° to 20°S, while Banakar et al. (1998) reported the lysocline at ~4,400 m and the CCD at ~4,700 m, indicating active carbonate dissolution processes that vary with latitude and water mass properties. Variations in remineralisation rates and surface productivity contribute to temporal and spatial shifts in the CCD and lysocline, affecting the preservation potential of biogenic carbonates (Derry, 2022; Schulte & Bard, 2003). Although several studies have addressed sediment provenance, deposition (Nath & Mudholkar, 1989; Nath et al., 2005; Purnachandra Rao & Nagender Nath, 1988), their transport pathways (Nath et al., 1992, 2007), radiolarian or radiochemistry-based palaeoceanography (Gupta, 2002, 2003), long-term, species-specific records of foraminiferal calcification and dissolution in the CIB remain limited. Carbonate preservation studies on seamount top records have linked intensified dissolution during Marine Isotope Stages (MIS) 11, 9, and 7 to the increased influence of AABW (Nath et al., 2013). Further insights from dissolution proxies such as the G. menardii fragmentation index and perfect test ratio have revealed selective dissolution in CIB sediments, highlighting the necessity of using robust, dissolution-resistant taxa for reliable palaeoenvironmental reconstructions (Sijinkumar et al., 2022).
Geographic location of sediment cores BC37 and SVBC37 recovered from the Central Indian Basin (CIB), Indian Ocean. The inset map displays detailed multi-beam bathymetry of the core sites, highlighting local seafloor topography and core positioning within the basin.
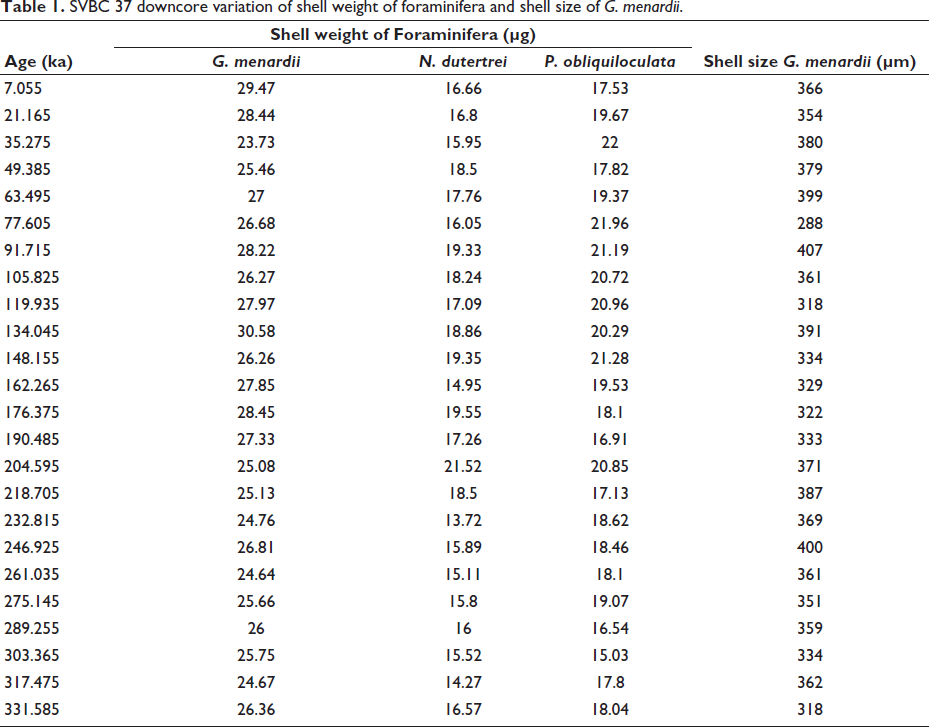
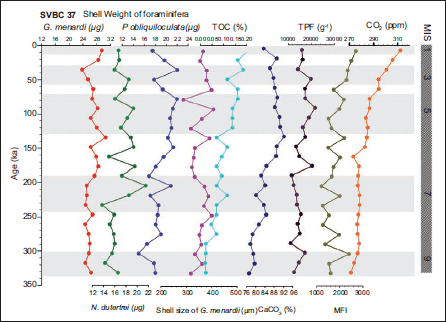
SVBC 37 downcore variation of shell weight of foraminifera and shell size of G. menardii.
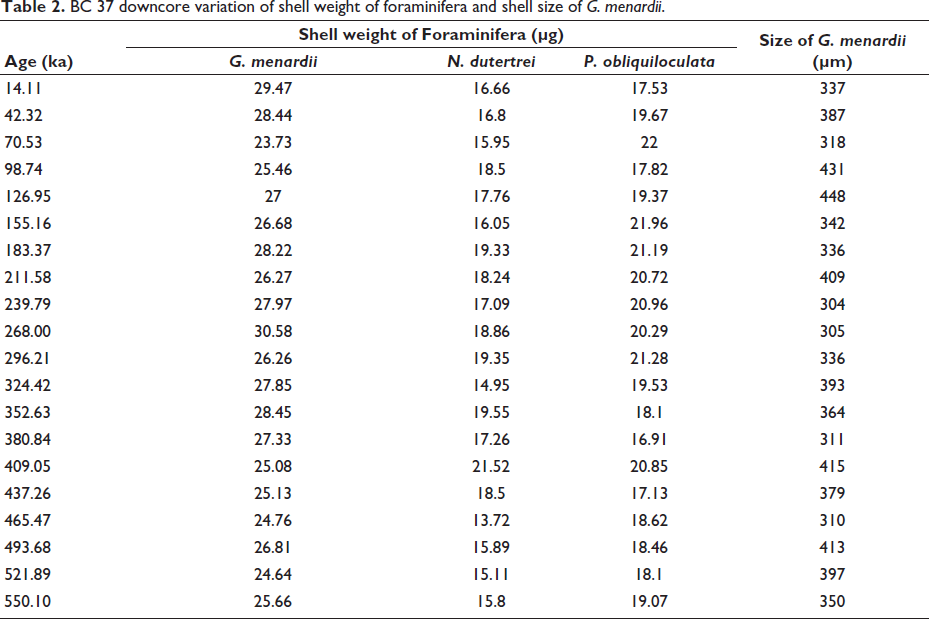
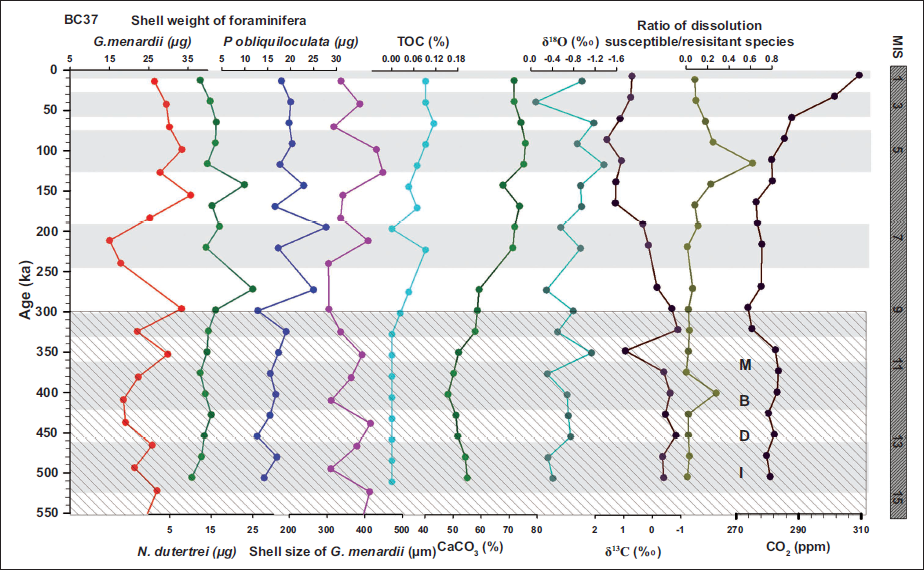
BC 37 downcore variation of shell weight of foraminifera and shell size of G. menardii.
The present study explores a multi-species, multi-proxy approach integrating shell weight, shell size, dissolution features, and planktonic foraminiferal species abundance data to investigate patterns of calcification and preservation in two sediment cores from the CIB, spanning the last ~600 ka. We focus on three species of planktonic foraminifera with varying susceptibility to dissolution, namely, P. obliquiloculata, G. menardii, N. dutertrei, to reconstruct changes in foraminiferal test characteristics across glacial-interglacial cycles. By combining morphometric measurements with SEM and faunal assemblage analysis, this study provides insights into the evolution of surface-ocean carbonate chemistry and deep-sea dissolution processes (Figure 2). The key objectives are to examine species-specific variability in calcification and dissolution across multiple climatic intervals, to evaluate the effectiveness of shell-based morphometric proxies in quantifying carbonate preservation and to infer broader implications for oceanic carbon cycling and atmospheric CO2 regulation during the Quaternary. These findings aim to enhance our understanding of tropical ocean-climate interactions and the role of biogenic calcifiers in modulating the long-term carbon cycle, particularly through their responses to glacial-interglacial variations in carbonate dissolution and preservation.
Conceptual illustration showing the vertical structure of the water column in the CIB, highlighting key oceanographic features such as the lysocline and CCD. The schematic also illustrates the methodological approach used for assessing foraminiferal shell weight and shell size, including sampling depths and analytical techniques.
MATERIALS AND METHODS
Sampling and age model
This study utilises two box cores, BC37 (16°06.031′S, 75°26.040′E; water depth: 4252 m; core length: 0.40 m) and SVBC37 (16°06.943′S, 75°25.083′E; water depth: 3,992 m; core length: 0.25 m) retrieved from the seamount in the CIB during the 4th and 26th expeditions of RV Akademik Boris Petrov (Figure 1). The seamount rises approximately 1,200 m above the surrounding seafloor (~5,100 m depth) and is situated along the flanks of the 76°30′E fracture zone (Nath et al., 2013). Core BC37 was subsampled at 2 cm intervals, and SVBC37 at 1 cm intervals, for the present investigation. The lithology of BC37 reveals two distinct sedimentary units; the upper 0–16 cm consists of foraminifera-rich, homogeneous greyish-orange-pink sediments, while the lower part is composed of moderately brown to yellowish-brown sediments intercalated with dark yellowish-orange layers. SVBC 37 consists of brown to white foraminiferal ooze. The positioning of the cores relative to the seamount summit presents a unique opportunity to investigate the palaeoceanographic conditions that influenced carbonate sediment deposition within a predominantly siliceous ooze-dominated region (Nath et al., 2013). For chronological control, the study adopts previously published age models for both cores without modification (Nath et al., 2013).
Foraminiferal analysis
The sediment samples were disaggregated by soaking in distilled water and then wet-sieved using a mesh with a pore size of >63 µm. The coarse fraction was oven-dried at approximately 60°C and then further dry-sieved to isolate the 250–350 µm size fraction. From this fraction, planktonic foraminifera species P. obliquiloculata, G. menardii, and N. dutertrei were picked under a stereo-zoom binocular microscope (Olympus SZ61), with ~50 specimens selected per sample for shell weight analysis. Specimens with secondary calcite overgrowths, textural changes due to recrystallisation, or infillings of detrital particles and coccoliths were carefully excluded. Shell weights were measured using an Aczet CM-6 microbalance with a precision of ±1 µg. Shell size measurements were carried out for G. menardii using specimens from the fraction >250 µm. We measured the maximum diameter of each G. menardii specimen. At least 10 well-preserved individuals were measured per sample using a Leica M205C stereo-zoom microscope equipped with a micrometre scale on the stage. For SEM, selected specimens were gold-coated to enhance conductivity and backscattering of secondary electrons. SEM imaging was performed using a JEOL JSM-IT800 digital SEM with a tungsten filament, allowing detailed observation of microstructural features indicative of dissolution, such as etching, wall thinning, and fragmentation.
RESULTS
Shell Weight Variations
In the shallower core SVBC37, the shell weights of G. menardii, N. dutertrei, and P. obliquiloculata display distinct temporal patterns over the last ~350 kyr (Figure 3 and Table 1). Shell weights of G. menardii range from 23.7 to 30.6 µg, with the lowest value recorded at 35 ka (MIS 3) and the highest at 134 ka (MIS 6). P. obliquiloculata exhibits a weight range of 15.0 to 22.0 µg, with a minimum at 303 ka (MIS 9) and a maximum at 35 ka (MIS 3). N. dutertrei ranges between 13.7 and 21.5 µg, with the lowest value at 232 ka and the highest at 204 ka, both within MIS 7 (Figure 3). In core BC37, a broader range of shell weights is observed. G. menardii shows values between 15.0 µg (211 ka; MIS 7) and 35.7 µg (155 ka; MIS 6). P. obliquiloculata ranges from 12.7 µg (465 ka; MIS 13) to 27.8 µg (211 ka; MIS 7). For N. dutertrei, the lowest weight (12.3 µg) is recorded at 380 ka (MIS 11), while the highest (25.0 µg) occurs at 268 ka (MIS 8) (Figure 4). These variations suggest species-specific responses to changing oceanographic conditions during glacial-interglacial transitions.
Downcore variations in shell weight (µg) of G. menardii (red), P. obliquiloculata (green), and N. dutertrei (blue), together with shell size (µm) of G. menardii (pink), from sediment core SVBC 37, plotted against age (ka). Also shown are total organic carbon (TOC), CaCO₃ content, total planktonic foraminifera (TPF), and the Menardii Fragmentation Index (MFI), adopted from Nath et al. (2013), along with atmospheric CO₂ concentrations derived from the Vostok ice-core record. Marine Isotope Stages (MIS) are indicated on the right; shaded intervals represent interglacial periods, whereas unshaded intervals denote glacial conditions.
Shell size variability in G. menardii
Shell size measurements of G. menardii show clear glacial-interglacial trends in both cores. In SVBC37, the largest shells (~407 µm) occur at 110 ka, likely corresponding to the warm interglacial MIS 5. The smallest size (~288 µm) is recorded at 78 ka, associated with a shift toward glacial conditions (Figure 3 and Table 2). These size variations imply ecological sensitivity to surface-water temperature and productivity. In BC37, G. menardii sizes vary from ~304 to 448 µm. The largest specimens (~448 µm) are found around 127 ka (MIS 5), a period of enhanced warmth and stratification, while the smallest specimens (~304 µm) are found at approximately 240 ka (MIS 7/8), representing a glacial phase (Figure 4). These patterns support the interpretation that G. menardii exhibits larger sizes under warm, productive conditions and reduced sizes during colder, nutrient-stressed intervals.
Downcore variations in shell weight (µg) of G. menardii (red), P. obliquiloculata (green), and N. dutertrei (blue), together with shell size (µm) of G. menardii (pink), from sediment core BC37. Marine Isotope Stages (MIS) are indicated to facilitate interpretation of glacial–interglacial variability. Total organic carbon (TOC), CaCO₃ content, and stable isotope records (δ¹³C and δ¹⁸O) are adopted from Nath et al. (2013), while atmospheric CO₂ concentrations are derived from the Vostok ice-core record.
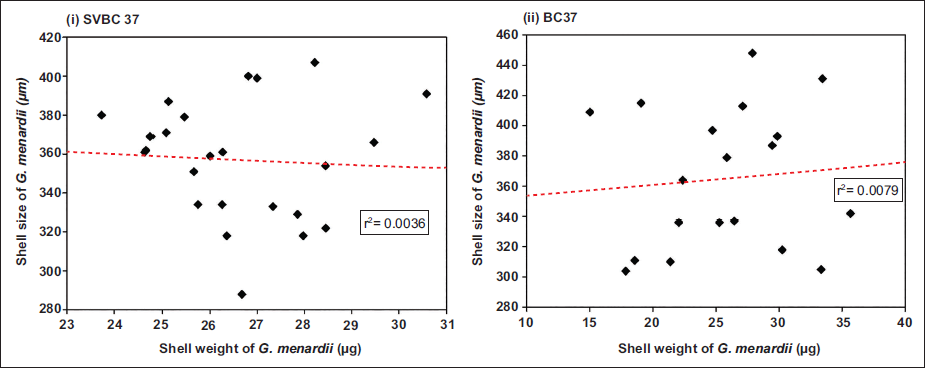
Relationship between shell weight and size
To assess the influence of size on calcification, a correlation analysis was conducted between shell weight and shell size of G. menardii for both cores (Figure 5). In SVBC37, the scatter plot reveals a very weak linear relationship (r2 = 0.0036), indicating that shell weight does not systematically increase with shell size (Figure 5(i)). A similar pattern is observed in BC37, where the r2 value is slightly higher at 0.0079 (Figure 5(ii)), but still negligible. These results suggest that shell weight in G. menardii is not directly controlled by shell size in the studied records.
Scatter plots illustrating the relationship between shell weight and shell size of G. menardii from cores SVBC37 (left) and BC37 (right). Red dashed lines represent linear regression fits. Both plots show weak correlations, suggesting that shell weight is largely independent of shell size in the studied intervals.
DISCUSSION
Planktonic foraminifera: calcification and dissolution
The shell weight records of G. menardii, P. obliquiloculata, and N. dutertrei from the CIB reflect a complex interplay of species-specific ecological preferences, regional hydrography, and basin-scale carbonate chemistry. Shell weight serves as a net signal of both primary calcification in the upper ocean and post-depositional preservation on the seafloor. P. obliquiloculata, a subsurface-dwelling species, aligns with the classical view that warmer, stratified interglacial conditions enhance calcification, as seen during MIS 7. However, its anomalous weight increase during MIS 3 and decline during MIS 13 suggest that thermocline structure and nutrient delivery may modulate calcification independently of surface temperature (Duplessy et al., 1981; Sijinkumar et al., 2022). Conversely, G. menardii and N. dutertrei exhibit shell weight maxima during glacial periods (MIS 6 and MIS 8, respectively), indicating that cooler, denser waters may enhance preservation through increased saturation with carbonate ions. This deviates from the expected patterns observed in other Indo-Pacific sites, highlighting the dominant role of regional factors, such as water mass structure and carbonate chemistry, in the calcification process. These species-specific responses also reflect ecological adaptations to varying depth habitats, with N. dutertrei particularly sensitive to thermocline productivity and subsurface water mass changes (Fehrenbacher et al., 2017; Ravelo & Fairbanks, 1992).
Glacial-interglacial cycles: interplay of productivity and dissolution
The shell weight and size measurements, along with planktonic foraminiferal assemblage data, reveal how glacial-interglacial cycles modulate the interplay between surface-ocean productivity and deep-water carbonate dissolution, ultimately shaping the preservation patterns of calcareous microfossils. P. obliquiloculata shell weight data support the traditional model of enhanced calcification during interglacial periods (MIS 7), whereas the glacial maxima in shell weights of G. menardii and N. dutertrei point to improved carbonate preservation driven by reduced atmospheric CO2 levels and increased carbonate saturation in bottom waters. The influx of AABW, particularly during specific transition periods and the last interglacial (MIS 5), may have introduced corrosive, carbonate-undersaturated waters into the CIB, thereby amplifying dissolution and altering shell preservation. The sharp weight reductions across all species during MIS 11, corresponding to the Mid-Brunhes Dissolution Interval (MBDI), indicate basin-wide carbonate undersaturation (Barker et al., 2006; Barth et al., 2018; Nath et al., 2013). The observed loss in shell weight likely reflects diagenetic overprinting, particularly post-depositional dissolution, rather than true variability in primary biological calcification. This inference is based on weak correlations between shell weight and size, SEM evidence of surface etching and test thinning, and the positioning of the sediment cores near or below the lysocline and CCD, where dissolution is intensified. This distinction is crucial, as relying solely on shell weight without evaluating the preservation state can lead to misinterpretation of palaeoceanographic signals. Seafloor conditions, including carbonate saturation levels, sediment porewater chemistry, and proximity to critical depth horizons, can significantly alter original calcification signals. Therefore, disentangling primary biogenic inputs from secondary diagenetic effects is crucial for accurately reconstructing past ocean carbonate chemistry and climate variability. When interpreted within the framework of glacial-interglacial variability, the records reveal a complex interplay between monsoonal productivity, carbonate ion availability, and post-depositional dissolution.
Preservation versus dissolution effects
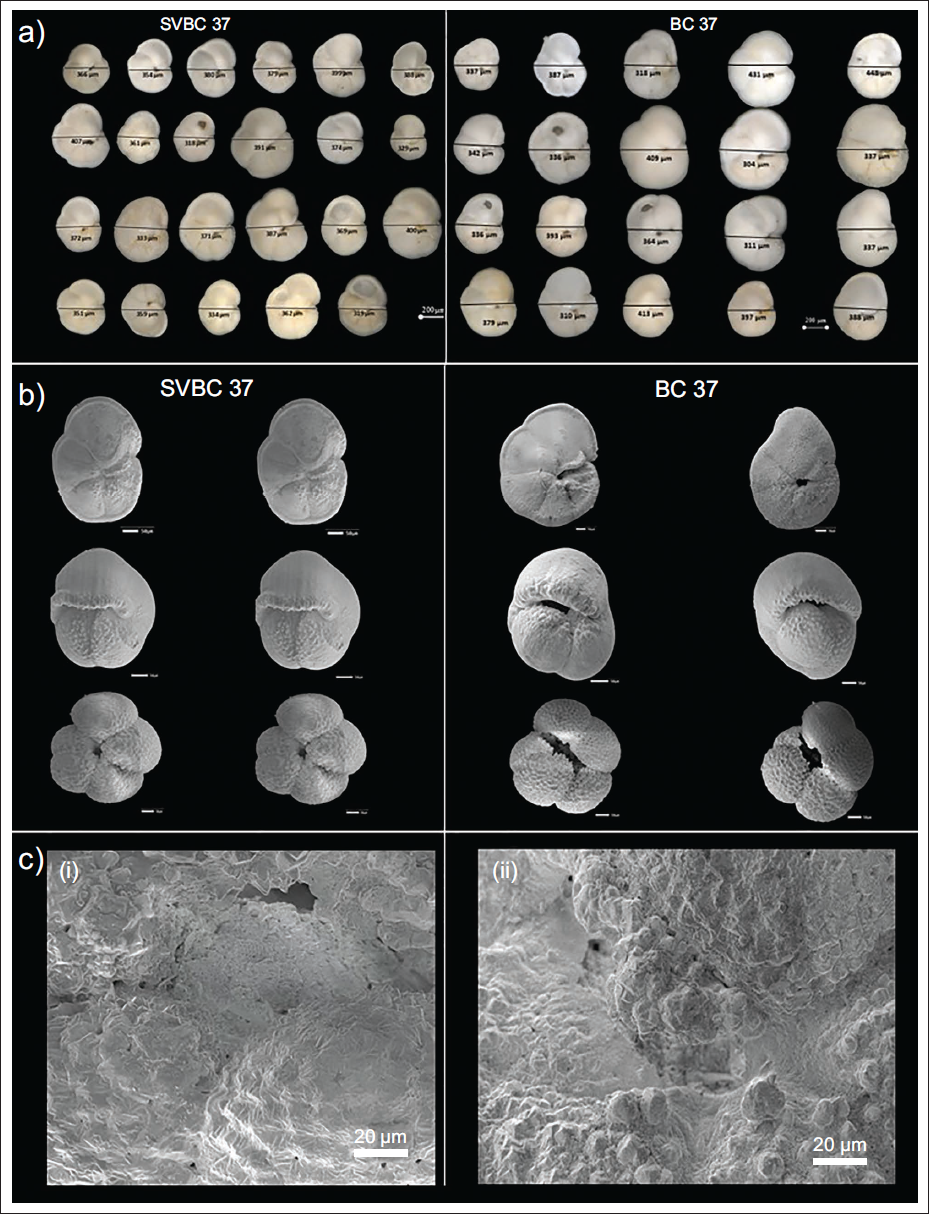
The preservation of biogenic carbonate is strongly tied to water depth, lysocline dynamics, and carbonate saturation levels. SVBC37 (3992 m) lies near the modern lysocline (~4,000–4,400 m), while BC37 (4252 m) is closer to or within it (Banakar et al., 1998; Kolla et al., 1976). The heavier shell weights and foraminiferal ooze in SVBC37 suggest better preservation, whereas BC37 has mixed clay-rich layers that are too conducive to implying more active dissolution (Nath et al., 2013). This depth-dependent dissolution is further influenced by the incursion of AABW (Nath et al., 2013), which contributes to carbonate undersaturation at abyssal depths (Corliss, 1979). The relatively higher dissolution observed during glacial-interglacial transitions and the last interglacial (MIS 5) aligns with periods of more substantial AABW influence, as indicated by reduced shell weight and more intense test degradation. The SEM observations support this at MIS 7 (~204 ka), N. dutertrei displays features of incipient dissolution such as etching, wall thinning, and weakened ornamentation (Figure 6). At MIS 11, more pronounced dissolution, manifesting as honeycomb textures and the complete loss of surface detail, points to prolonged exposure to corrosive pore waters or bottom water under saturation (Fehrenbacher et al., 2017; Mekik et al., 2012). These findings offer critical insights into the preservation dynamics of biogenic carbonates across glacial-interglacial cycles, with broader implications for the long-term carbon cycle and the ocean’s buffering capacity in response to shifts in deep-water carbonate chemistry.
(a) Shell size measurements of G. menardii from (i) SVBC 37 and (ii) BC 37 cores. (b) Scanning Electron Microscope (SEM) images of G. menardii, P. obliquiloculata, and N. dutertrei from (i) SVBC 37 and (ii) BC 37 cores, (c) Detailed SEM images of N. dutertrei from BC37 illustrating preservation states: (i) Specimen at ~204 ka showing early-stage dissolution features-etched surface texture, test wall thinning, loss of original ornamentation (broken/flattened spines), and signs of structural weakening. (ii) Specimen at ~407 ka showing advanced dissolution, honeycomb-like surface texture, nodular corrosion, circular pitting, and smoothed test surface indicative of prolonged exposure to corrosive bottom waters.
Shell weight and size in G. menardii: environmental and diagenetic controls
Shell size analysis was conducted on individuals from the >250 µm size fraction, effectively normalising the overall test size range. Within this fraction, 10 randomly selected specimens were measured for size, while shell weight was calculated as an average of 50 specimens from the same size class. Given the potential variability within the 250–350 µm range, a strong correlation between individual shell size and average shell weight is not necessarily expected. Indeed, the extremely weak correlation observed in G. menardii (r2 < 0.01) supports this expectation and suggests that test diameter alone does not account for variability in shell weight. This disconnect likely reflects the influence of other factors such as test wall thickness, internal porosity, and particularly post-depositional dissolution processes, which reduce shell mass while leaving outer dimensions essentially unchanged (Mekik & Francois, 2006). Furthermore, larger shell sizes observed during interglacial periods may reflect more favourable ecological conditions for calcification, yet this increase in size does not consistently correspond to heavier shells, underscoring the role of differential dissolution and preservation variability (Barrett et al., 2025; Davis et al., 2019). The high total organic carbon (TOC) and low CaCO3 concentrations in the BC37 core sediments further reinforce this interpretation, indicating active carbonate dissolution in undersaturated bottom-water conditions, particularly where elevated biogeochemical activity enhances porewater acidity (Nath et al., 2013). Taken together, these findings underscore that both environmental drivers and diagenetic overprinting, such as shifts in the lysocline and proximity to the CCD, collectively modulate calcification and preservation dynamics.
CONCLUSIONS
This study presents planktonic foraminiferal shell weight and size records spanning the last ~600 ka to unravel patterns of calcification and dissolution in the CIB. The species-specific data from G. menardii, P. obliquiloculata, and N. dutertrei reflect both ecological responses to surface-water conditions and post-depositional modifications influenced by deep-sea carbonate chemistry. Enhanced shell weights during glacial periods, particularly in G. menardii and N. dutertrei, suggest improved preservation linked to higher carbonate saturation states. In contrast, marked reductions in shell mass during MIS 11 indicate intense dissolution associated with the Mid-Brunhes climatic shift. SEM observations confirm progressive test degradation, reinforcing the role of lysocline proximity and corrosive deep-water incursions in shaping preservation trends. Overall, our data indicate better preservation and reduced carbonate dissolution during glacial periods compared to interglacial stages, likely due to shifts in deep-water chemistry and carbon cycling. The decoupling of shell weight and size, on the other hand, underscores the importance of diagenetic overprints and bottom-water chemistry. Overall, this study enhances our understanding of carbonate preservation dynamics and their modulation by offering valuable insights into the response of marine calcifiers to both past and future shifts in ocean carbonate chemistry.
Footnotes
Acknowledgements
The Ministry of Earth Sciences (MoES), Government of India, New Delhi, is gratefully acknowledged for providing ship time aboard the RV Akademik Boris Petrov and for funding the project through PMN-EIA. We sincerely thank the captain and crew of the vessel, as well as the cruise participants from CSIR-NIO, for their support during the expeditions. AVS acknowledges the ANRF, New Delhi, for the financial support under the CRG Grant (CRG/2023/002751). This is NIO’s contribution number 7514.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This research was supported by the ANRF, New Delhi, under the CRG Grant (CRG/2023/002751).