
Abstract
We report the presence of two additional echinoid species, Eupatagus faurai (Lambert, 1927) and Echinolampas vilanovae (Cotteau, 1890, Échinides éocènes de la province d’Alicante), from the Sylhet Limestone of Mikir Hills (MH), Assam, North East India. The two species from the middle Eocene of Assam are well recognised under the genera Eupatagus and Echinolampas and are found associated with echinoid fossil spines. These are recorded for the first time from the sediments of the Sylhet Limestone in the MH, Assam. The horizon containing the echinoid faunas described here is recorded in SBZ 16–18, which corresponds to the late Lutetian-Bartonian age of the middle Eocene, as previously reported for the larger benthic foraminifera-bearing bed. The discovery of the present additional echinoids in the Sylhet Limestone of MH provides fascinating evidence for a direct marine connection of the Neo-Tethys Seaway during the Eocene between Northeast India, the northwestern Himalayas, and Western Europe. These fossils support the presence of a warm, shallow marine sea. These findings enhance our understanding of the culmination of the Neo-Tethys Seaway, eventually leading to the formation of the mighty Himalaya.
INTRODUCTION
Echinoids are exclusively marine animals fitting to the phylum Echinodermata and the class Echinoidea, characterised by the presence of a mesodermal endoskeleton with calcareous plates. They are invertebrate and bottom-dwelling benthic organisms. They have a hard shell (referred to as a test) covered with small knobs (tubercles) to which movable spines are attached in living echinoids. The test and spines are the preserved parts usually found as fossils. Another unique feature of echinoids is the water vascular system, consisting of tubes and bladders that perform various functions, including locomotion, respiration, and feeding. Echinoids first appeared in the Ordovician but were not successful, and other groups, such as crinoids, dominated during the Palaeozoic (Srivasatava, 2014; Tappero, 2021). Many of the earlier echinoderm groups were either extinct or declined, and rare occurrences of echinoids are known from the early Jurassic, after which they diversified and have thrived up to the present day (Srivastava, 2014). Echinoids serve as palaeoenvironmental indicators, particularly of sediment type (soft or hard) and substrate granulometry. They provide valuable information about ancient marine environments, inhabiting from the intertidal zone to the deep-sea trenches (Kroh & Mooi, 2025). This article records two echinoid species, Eupatagus faurai and Echinolampas vilanovae, from the fossiliferous sandy limestone horizons of middle Eocene age exposed in Mikir Hills (MH) of Assam, India (Figure 1). These species have also been previously described from Eocene deposits in Spain (Carrasco, 2024). Earlier, Lokho et al. (2023) reported two species of echinoids, namely Porocidaris schmidelii and Ilarionia sindensis, from the same sandy limestone horizon in the studied section of samples Nos. 18 and 19, which are of middle Eocene age, as reported by Biswal et al. (2021).
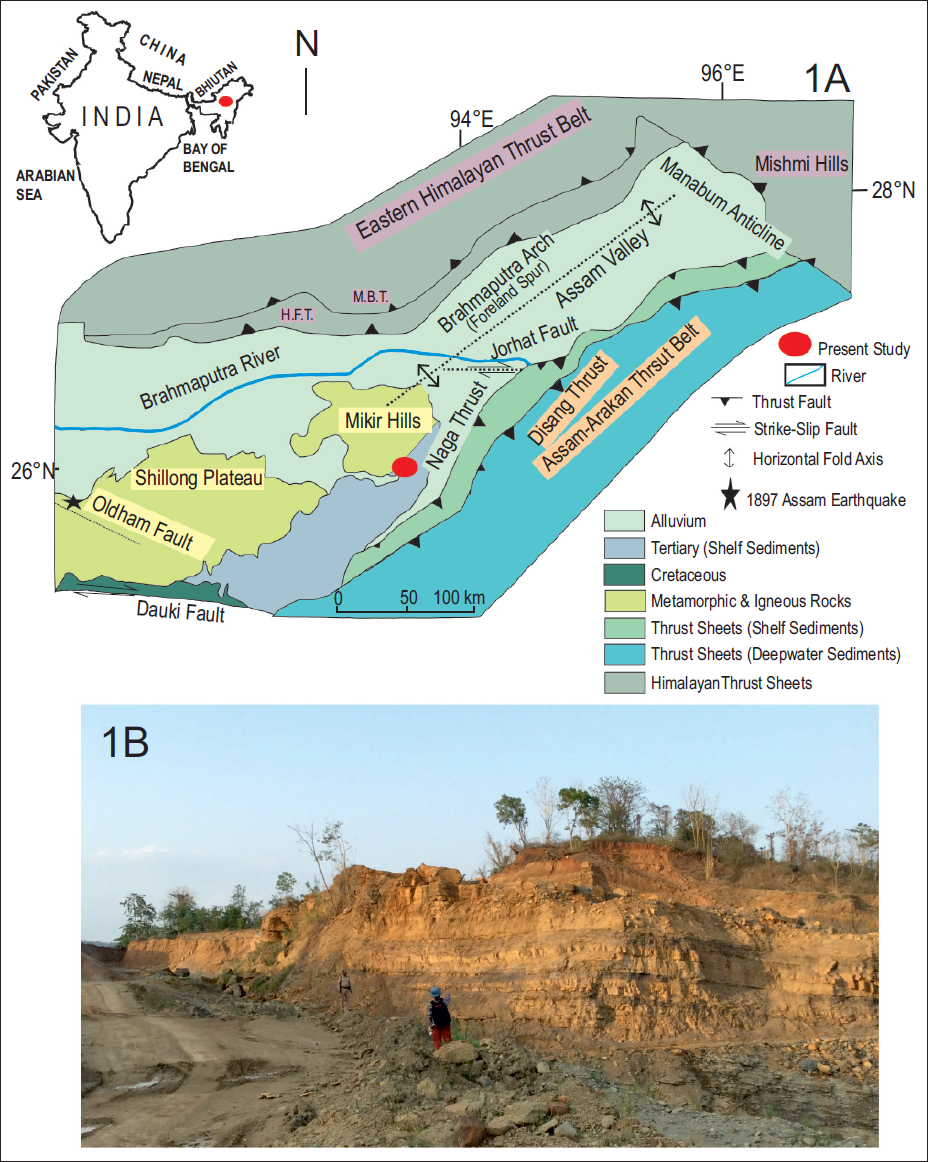
A. Geological map of Assam and surrounding areas (after Berger et al., 1983) showing the location of the Mikir Hills and the present study area with the echinoid-bearing horizon. B. Field photograph of the echinoid-bearing horizon.
In India, the Echinolampas genus was first recorded from the Eocene sediments of Kachchh by d’Archiac and Haime (1853). Further, they were reported from the Eocene of Assam (now in Meghalaya) by Medlicott (1869) and the Miocene rocks of Kachchh by Duncan and Sladen (1883). Roy and Das Gupta (1970) described three forms of Echinolampas from Eocene beds of Cutch. Additional Echinolampas were described from Oligocene rocks of Kutch by Tandon (1973), and a new genus, E. jigniensis, was described from Eocene rocks of Jammu in the NW Himalaya by Srivastava et al. (1992). Srivastava (1988) stated that numerous species of Echinolampas were known from the Tertiary sediments of Kutch, but they have an inadequate stratigraphic distribution, as reviewed in the Tertiary echinoids from India. The present findings on echinoids provide deeper insight into the presence of a warm, shallow marine environment in which they lived in the past. Further, the fossil record provides insight into the demise of the Neo-Tethys Sea. This process ultimately led to the orogeny and the formation of the mighty Himalayan fold mountains.
GEOLOGY OF THE AREA
The Shillong-Mikir Plateau (SMP) is part of the Indian Shield. It is one of the most seismically active zones in the world, separated from the peninsular shield and displaced eastward by approximately 300 km along the Dauki Fault (Baruah et al., 2011; Evans, 1964). The massive E–W trending Dauki Fault separates the plateau to the north and the Bengal basin to the south (Kayal, 2001). The Brahmaputra River, on the other hand, separates the plateau from the Himalaya to the north (Baruah et al., 2011; Kayal, 2001; Nandy, 2017). The MH lie in the East Karbi-Anglong district of Assam and are separated from the Shillong Plateau by the NW-SE trending Kopili Fault (Acharyya et al., 1986; Evans, 1964; Kumar et al., 2017; Sarma & Dey, 1996). The
REPOSITORY AND INSTITUTIONAL ABBREVIATIONS
Illustrated Echinoid specimens are curated in the Wadia Institute of Himalayan Geology (WIHG), Dehradun repository bearing Nos. WIMF/A4927 for E. faurai (Lambert, 1927) and WIMF/A 4928 for E. vilanovae (Cotteau, 1890). The compared studied specimens are curated in the Museo Geológico del Seminario de Barcelona, Spain, bearing numbers 52170MGB for E. vilanovae (Cotteau, 1890) and 7672MGSB for E. faurai (Lambert, 1927).
SYSTEMATIC PALAEONTOLOGY
The systematics proposed by Carrasco (2024) are followed.
Class Echinoidea Leske, 1778
Infraclass Irregularia Latreille, 1825
Subclass Neognathostomata Smith, 1981
Order Spatangoida L. Agassiz, 1840
Suborder Brissidina Stockley et al., 2005
Family Eupatagidae Lambert, 1905
Genus Eupatagus L. Agassiz, 1847
E. faurai (Lambert, 1927)
Plate 1, Figure 1a–e
1970 Eupatagus cossmanni (Lambert, 1902); Roman et Villatte in Reguant et al. p. 909, pl. 34, Figure 4a–4c.
2016 E. faurai (Lambert, 1927; Llansana & Romero, p. 166, Figure n/n.
2021 E. faurai (Lambert, 1927; Carrasco, 2021, p. 131).
2024 E. faurai (Lambert, 1927; Carrasco, 2024, p. 503, Lámina 58, Figure 2a–2d.
The fossil specimen is very similar to Eupatagus (Eupatagus) sp. recorded from the Siju Formation of Garo Hills, Meghalaya, India by Srivastava and Garg (2014) in having Petal III indistinct, Petals I and V longer than the Petals II and IV and close other fossils specimen of E. alatus Arnold Clark described from the Eocene sediments of Jamaica (Arnold & Clark, 1927). However, it differs in having longer Petals (I and V), a posteriorly protruded test, and a lesser number of tubercles. It is also easily distinguished from other species of Eupatagus cf. antillarum (Cotteau, 1890) in having a less broad, oval test, as reported by Donovan and Rowe (2000) from the late Eocene sediments of the Clarimont Formation, White Limestone Group, Jamaica. Our specimen differs markedly from many echinoid specimens of diverse ages such as Eupatagus rostratus described by Srivastava (2004) from the Eocene and Oligocene; E. rostratus by Duncan and Sladen (1884) from Oligocene and Miocene sediments of Kutch; Eupatagus sp. by Srivastava and Singh (2008) from the early Eocene of the Khuiala Formation, Rajasthan; Eupatagus sp. by Srivastava (1981) from Kutch; and Eupatagus singhi by Srivastava (1978) from Middle Oligocene sediments of Kachh, in having a contrasting and distinctive set of morphometric characters of the test and petals.
Class Echinoidea Leske, 1778
Infraclass Irregularia Latreille, 1825
Subterclass Neognathostomata Smith, 1981
Superorder Luminacea Mongiardino Koch et al., 2022
Order Echinolampadacea Mongiardino Koch et al., 2018
Suborder Echinolampadoida Kroh & Smith, 2010
Family Echinolampadidae Gray, 1851
Genus Echinolampas Gray, 1825
E. vilanovae (Cotteau, 1890)
Plate 2, Figure 1a–1e
1890 E. vilanovae (Cotteau, 1890, p.73, pl. IX, Figures 1–5).
1935 E. vilanovae (Cotteau, 1890; Lambert, 1927, p. 367).
1945 E. vilanovae (Cotteau, 1890; Darder, 1945, pl. 10, Figure 217).
2024 E. vilanovae (Cotteau, 1890; Carrasco, 2024, p. 647, Lámina 28, Figure 1a–1d.
Our fossil specimen is similar to Echinolampas qattamiaensis reported from Egypt by Ali (1983), which has a nearly circular test, slightly longer than wide, with a centrally located apical system, petals that almost reach the margin, and a slightly conical lateral profile. The specimen differs from Echinolampus jigniensis reported from Jammu in the NW Himalaya by Srivastava et al. (1992) from the Eocene rocks, which is characterised by globular, large tests with petals reaching up to the ambitus. Our specimen differs from Echinolampus khariensis, reported from Kutch by Srivastava and Singh (1999), in that it has a dome-shaped test, longer than wide and wider than high, with the highest point at the central region; the adoral surface is almost flat but slightly depressed at the peristome. Petals II and IV are broader and larger than the posterior Petals I and V.
DISCUSSION
The Eocene Epoch (56 to 33.9 Ma Ago) represents a dynamic geologic interval characterised by a complex interplay of tectonics and climate. It is characterised by a plethora of climatic conditions that include the transition from a warm, ice-free ‘greenhouse’ to a cool ‘icehouse’ world (Evans et al., 2018; Goldner et al., 2014; Sexton et al., 2011; Sluijs et al., 2013; Villa et al., 2008; Zachos et al., 2008). The Eocene successions are exposed as patches at several places in India: the Subathu Formation of Northwest Himalaya (Gupta & Kumar, 2019; Mathur & Juyal, 2000), the Harudi Formation and Fulra Limestone of Kutch, western India (Samanta & Lahiri, 1985; Saraswati et al., 2018), Bhimanapalli Limestone of the Krishna-Godavari Basin (Singh et al., 2019), and the Pondicherry area of South India (Malarkodi et al., 2013; Venkatachala et al., 1988), Upper Disang Formation of the Naga Hills (Lokho et al., 2004, 2007, 2020 and the references therein), Kopilli Formation of Assam (Evans, 1964) and the Sylhet Limestone of Assam (Biswal et al., 2021, 2022; Lokho et al., 2023).
Wilson and Metre (1953) were the pioneer workers in the classification of the Upper Cretaceous-Eocene sequence of the Assam region, followed by subsequent workers, including Nagappa (1959) and Banerjee et al. (1971). Nagappa (1959) was the first to study the region’s microfaunal assemblages in detail. Later, Samanta (1973) identified five foraminiferal zones of middle to late Eocene age from the Kopili Formation of Mikir-North Cachar Hills, Assam. A fossil mammal from marine Eocene strata (Jaintia Group) of the MH was reported and described by Whiso et al. (2009). Whiso et al. (2003) highlighted the occurrence of Planorotalites palmerae, which defines the upper part of the lower Eocene in the lower succession of the Sylhet Limestone. Recently, Biswal et al. (2021) presented a detailed larger foraminiferal biostratigraphy, lithofacies, and stable carbon isotope analyses of the Sylhet Limestone and assigned a middle Eocene age to the studied succession. The present study is confined to the upper part of the Sylhet Limestone in the southeastern edge of the MH, Assam (Figure 1B). Based on this evidence, the age of the echinoid-bearing horizons is considered to be of Lutetian to Bartonian. Besides foraminifera and vertebrate mammalian lumbar vertebrae, several associated fish teeth were reported by Whiso et al. (2009), and more vertebrate remains were recorded by Biswal et al. (2022).
Earlier, Lokho et al. (2023) described two echinoid taxa, namely I. sindensis Duncan and Sladen (1884) and P. schmidelii Münster in Goldfus (1830) from the same studied section of MH, Assam. Srivastava and Garg (2014) recorded a single specimen of the genus Eupatagus L. Agassiz, 1847, brissid echinoid according to Fischer (1966) and Smith (2011), but Eupatagidae according to Kroh and Mooi (2025) from the fossiliferous limestone-marl succession of middle Eocene age belonging to the Siju Formation exposed along the Dilni River of Garo Hills, Meghalaya, India. The Eocene Epoch witnessed a prolific development of echinoids, with 15 genera recorded from India (Srivastava, 2004). The occurrence of these echinoids indicates shallow-water conditions in the area, characterised by normal temperatures and salinity, with less turbulent water and abundant food, under a tropical to subtropical climate. In India, Eocene echinoids are recorded from Assam and Meghalaya (Spengler, 1923; Srivastava, 2004; Srivastava & Garg, 2014); Kachchh (Duncan & Sladen, 1883; Srivastava et al., 2008) Kohat and Salt Range (Davies, 1943; Theobald, 1881); Sind (d’Archiac & Haime, 1853; Duncan & Sladen, 1882-1886, 1884). An irregular sea urchin, the genus Echinolampas, due to its unique shape and abundant presence in the country’s fossil record, is crucial to decipher the geological past of India. The presence of Echinolampas is a key indicator of a specific geological time period in India, mostly in Tertiary rocks of Gujarat, Rajasthan, Northeast India and Jammu of the NW Himalaya. Echinolampas has traditionally been assigned to the order Cassiduloida, a group of irregular echinoids characterised by bilateral symmetry, an evolutionary adaptation for a burrowing or semi-infaunal habitat. In India, we report, for the first time, the species E. vilanovae. The same species has been recorded from the province of Alicante (SE Spain) in the middle Eocene (Carrasco, 2024).
Eupatagus, which is also an irregular echinoid, exhibits an infaunal or burrowing mode of habitat. It has also been previously reported from the Siju Formation, Meghalaya, where this fossil was found in a highly fossiliferous zone of middle Eocene age. The present study records E. faurai for the first time from the MH, Sylhet Limestone, of the middle Eocene. Carrasco (2024) recorded E. faurai from the province of Alicante (SE Spain) of the middle Eocene. According to Carrasco (2024), E. faurai could be extended into the Middle Eocene of the Island of Mallorca in the Balearic Archipelago, which belongs to Spain and lies very close to the Iberian Peninsula. The previous reports by Biswal et al. (2021) of larger benthic foraminifera from the MH, Assam, indicate a warm, well-lit shallow marine environment, which is supported by the present work. The present discovery of E. faurai and E. vilanovae from the present study adds another layer to our understanding of the rich marine fauna that inhabited the MH of Assam during the Eocene Epoch (56–33 Ma). This reinforces the interpretation of a warm, shallow sea teeming with diverse ancient marine life before the culmination of the Neo-Tethys Sea, which was linked to the epic journey of the Indian subcontinent drifting northward, ceasing with the collision of Eurasia and the subsequent formation of the Himalayas. The presence of similar faunal assemblages, recorded in NE India, including Assam, and as far west as Spain, demonstrates the continuity of the Neo-Tethys Sea (Figure 2).
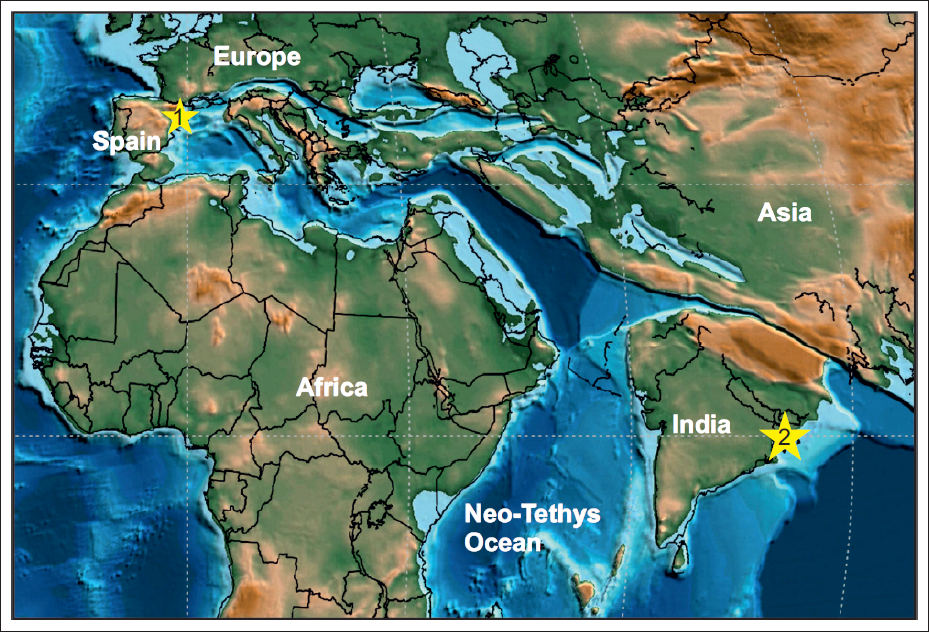
Palaeogeographic map of the Neo-Tethys Ocean showing the distribution of echinoids during the Eocene (adapted from Scotese, 2021). Number 1 marks the location of several regions in the eastern Iberian Peninsula, and Mallorca Island (Spain), and number 2 indicates the Mikir Hills of Assam (India).
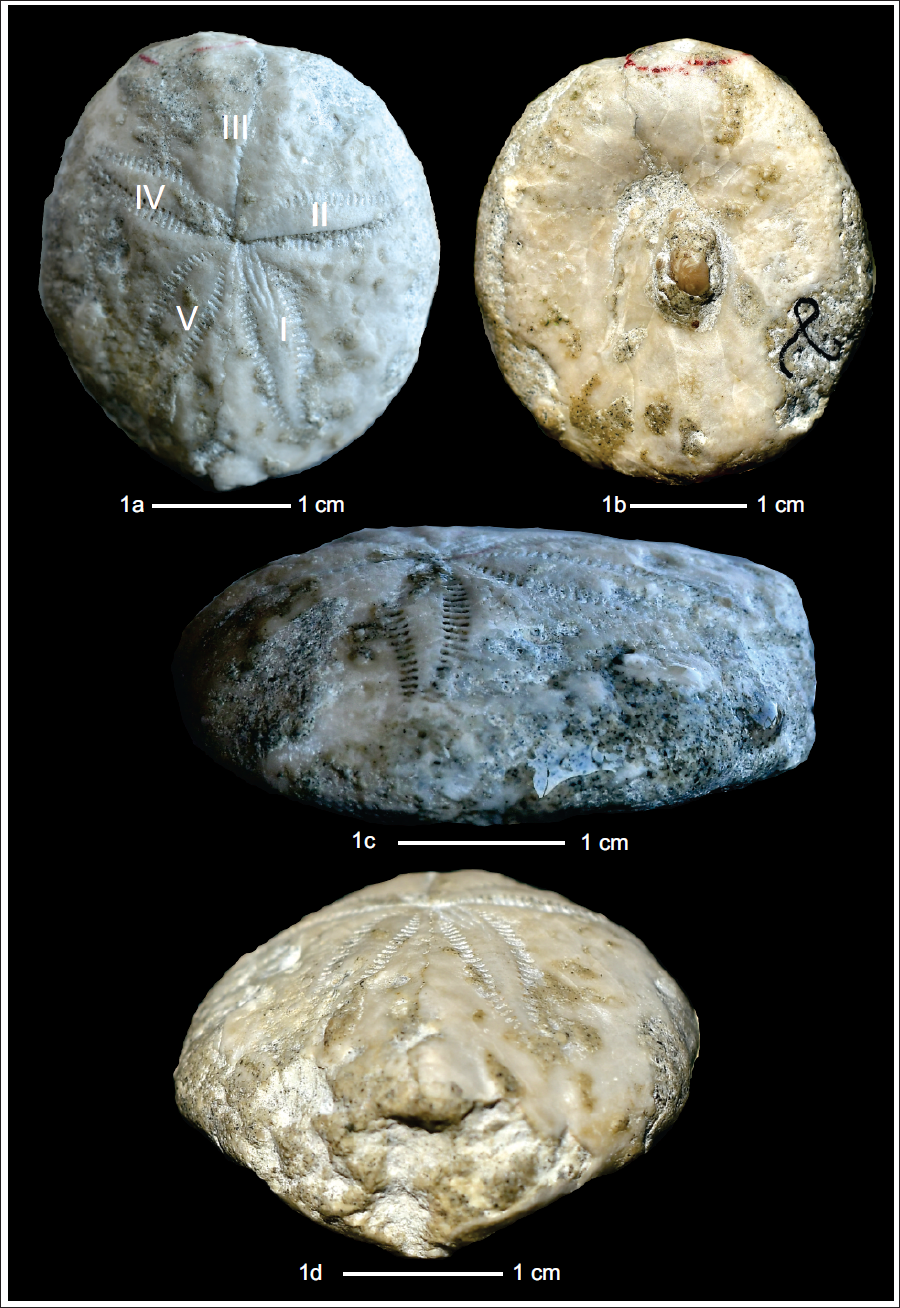
Eupatagus faurai (Lambert, 1927). 1a: aboral view; 1b: adoral view; 1c: lateral view; 1d: posterior view.
Echinolampas vilanovae (Cotteau, 1890). 1a: Aboral view; 1b: adoral view; 1c: Peristome; 1d: anterior view; 1e: lateral view; 1f: posterior view.

CONCLUSIONS
The echinoid taxa E. faurai and E. vilanovae are reported here for the first time from the Sylhet Limestone of MH, Assam, as well as from India.
This record, along with earlier reports of related genera from the Siju Formation of Garo Hills, Meghalaya, the Khuiala Formation of Jaisalmer, Rajasthan and the Subathu Formation of Jammu, (NW) Himalaya, suggests marine connectivity between northeastern, western India and western Himalaya during the middle Eocene, highlighting the palaeobiogeographic significance of the region.
The presence of species common to both the Indian region and Spain provides evidence of a shared marine fauna across the Neo-Tethys Sea during the Eocene, before its culmination, which was associated with the northward epic journey of the Indian subcontinent and the subsequent collision of Eurasia, leading to the formation of the mighty Himalaya.
Footnotes
Acknowledgements
KL is thankful to Dr Vineet K. Gahaluat, Director, Wadia Institute of Himalayan Geology, Dehradun, for providing the facilities and permission to publish this work (WIHG/Publication no. WIHG/0428). Sincere thanks are extended to the late Kholi Kaihrii, Loli Hekha Mao, for a safe drive, and to Adahrii Adakho and Kapani Khoziio for other valuable help in the field. KL sincerely thanks the authorities of Dillai Parbat Limestone Quarry, CCI Mines, Bokajan, Karbi Anglong District, Assam, for granting permission for fieldwork and for their kind hospitality. Authors also gratefully acknowledge Prof. Ashutosh K. Singh, Convener 29th ICMS-2024, Department of Geology, University of Delhi and Guest Editor, Special Issue JPSI, for the invitation to contribute this article. We are grateful to two anonymous reviewers who have significantly improved the manuscript.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors received no financial support for the research, authorship and/or publication of this article.