
Abstract
This article reports two echinoid taxa viz. Ilarionia sindensis Duncan and Sladen (1884) and Porocidaris schmidelii Münster in Goldfus (1830) from the middle Eocene Sylhet Limestone of Mikir Hills, Assam. P. schmidelii is found from the Lutetian (middle Eocene) to the Priabonian (upper Eocene) in the following regions: NE of Spain, Biarritz (Southwestern France), Angoumé (Southern Aquitaine, France), Carinthia (Southern Austria), Venetian region of Italy Provence, Southern Alps of French, Istria (Croatia), Persian Gulf, Oman and Egypt. I. sindensis was first reported from the Eocene of Sindh province from the Khirthar Series (Pakistan) and Madagascar. It is also recorded from the Bartonian–Priabonian in the following regions: NE Spain, Biarritz in Southwestern France, Angoumé in Southern Aquitaine-France, Carinthia in Southern Austria, Italian region of Veneto, Provence and southern French Alps, Persian Gulf, Oman, Egypt and Istria. They are systematically described to know their stratigraphic, palaeoenvironment and palaeogeographic distribution. The material studied herein represents the first report from the middle Eocene of India, and it significantly expands the geographical extension of Eocene marine echinoids in the northeastern part of India.
Introduction
The study of vulnerable marine ecosystems, such as seamounts and oceanic islands, is critical for the conservation and management of the marine ecosystems (Kvile et al., 2014; Watling & Auster, 2017). Echinoderms are exclusively marine invertebrate organisms and mostly bottom dwellers. So, they are good palaeoenvironmental and ecological indicators. Klein (1734) introduced the Phylum Echinodermata for sea urchins followed by Leuckart and Frey (1847) who recognised them as a distinct group from the radiata. They have well-developed organ systems which are ecologically and geologically important. This phylum contains about 7,000 known living species and more than 20,000 fossil species with 15 classes of extinct species, which has the most extinct classes than any other animal. They are found from poles to the equator and from the intertidal zone to depths of more than 5,000 m. Echinoderms appeared first around 530 Ma in the Cambrian and rapidly diversified into many groups but were not as successful as the crinoids subjugated during the Palaeozoic. Many of the echinoderm groups were either decreased in terms of abundance and diversity or extinct by the early Mesozoic. Rear occurrences are known in the early Jurassic and then diversified and thrived till date (Srivastava, 2014). Nowadays the living representatives of Echinoderms are Echinoidea (sea urchins), Crinoids (Sea lilies & feather stars), Holothuroidea (sea cucumbers) and Asteroidea (starfish) and Ophiuroidea (basket stars, brittle stars) (Tappero, 2021). In India, d’Archiac and Haime (1853) were the first to record the genus Echionolampas from the Eocene of Kachchh. Echinoids have also been reported from other parts of India like Meghalaya (Das & Bora, 1981; Srivastava & Garg, 2014), Mizoram (Tiwari & Jauhri, 2014), Assam (Spengler, 1923), Gujrat (Srivastava & McNamara, 2011), Himachal Pradesh (Reed, 1910), Tamil Nadu (Badve & Aziz, 1983) and Rajasthan (Srivastava & Mathur, 1996). The present study has been carried out in the Dillai Parbat limestone quarry, which lies on the southeastern edge of the Mikir Hills (Figure 1). The sedimentary rocks of limestone, sandstone and shale of the fossiliferous middle Eocene deposits of Sylhet Limestone of the study area have yielded both vertebrate and invertebrate faunas (Biswal et al., 2021, 2022; Venkatachalapathy & Whiso, 2009; Whiso et al., 2009). Based on the larger foraminiferal studies by Venkatachalapathy and Whiso (2009) and Biswal et al. (2021), the age of the Sylhet Limestone indicates an early middle Eocene age (early Lutetian) for the lower part of the succession and a late middle Eocene age (late Lutetian to Bartonian) for the upper part of the succession corresponding to Shallow Benthic Zones SBZ 13 and SBZ 16–18 of Serra-Kiel et al. (1998). The present paper aims to put into record the first-time finding of echinoids from the Sylhet Limestone of Mikir Hills and its implications on the palaeoenvironment and palaeobiogeography during the middle Eocene with a detailed systematic description.
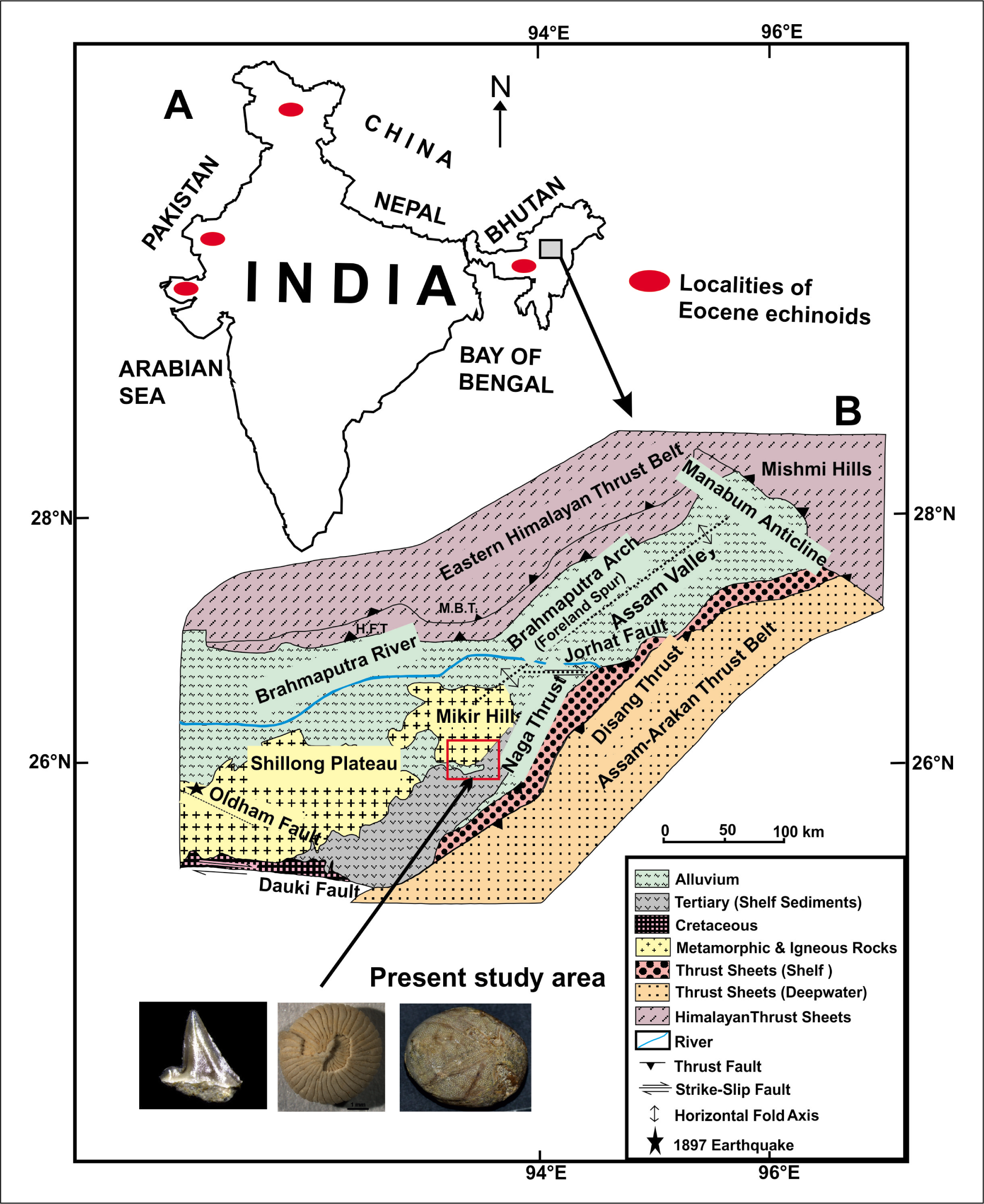
(A) Outline map of India showing the Eocene echinoids localities with a red ball; (B) Geological map of Assam and surrounding areas showing the location of the Mikir Hills and the present study area (after Berger et al., 1983).
Geological Background
The Assam-Arakan Basin is represented by the excellent exposure of Tertiary rocks, differentiated primarily into the following two facies: basin and shelf sediments. This differentiation is mainly marked in the Eocene strata of the succession. The shelf sediments occur in regions of northwest of the Halflong-Disang and Naga thrusts along the southern foothills of Garo, Khasi-Jaintia and Mikir-North Cachar Hills. The basinal facies are exposed in the eastern part of Nagaland-Manipur and Mizo Hills. The Mikir Hills (Assam) and the Shillong Plateau (Meghalaya) are known as the northeastern extension of the Precambrian Shield of the Indian Peninsula (Acharyya et al., 1986; Evans, 1964; Kumar et al., 2017; Nandy, 2017; Sarma & Dey, 1996). The NW-SE trending Kopili fault separates the Shillong Plateau from the Mikir Hills, which lie in the Karbi-Anglong district of Assam. The sediment deposition in this area was basically controlled by tectonic activities along the fault zones and the Indo-Asian plate collision during the late Palaeocene and early Eocene (Jauhri & Agarwal, 2001; Murty, 1983; Tewari et al., 2010), which is mainly reflected in the contrasting lithofacies in the Palaeogene sequence of the Assam shelf, Garo, Khasi, Jaintia and in Mikir Hills.
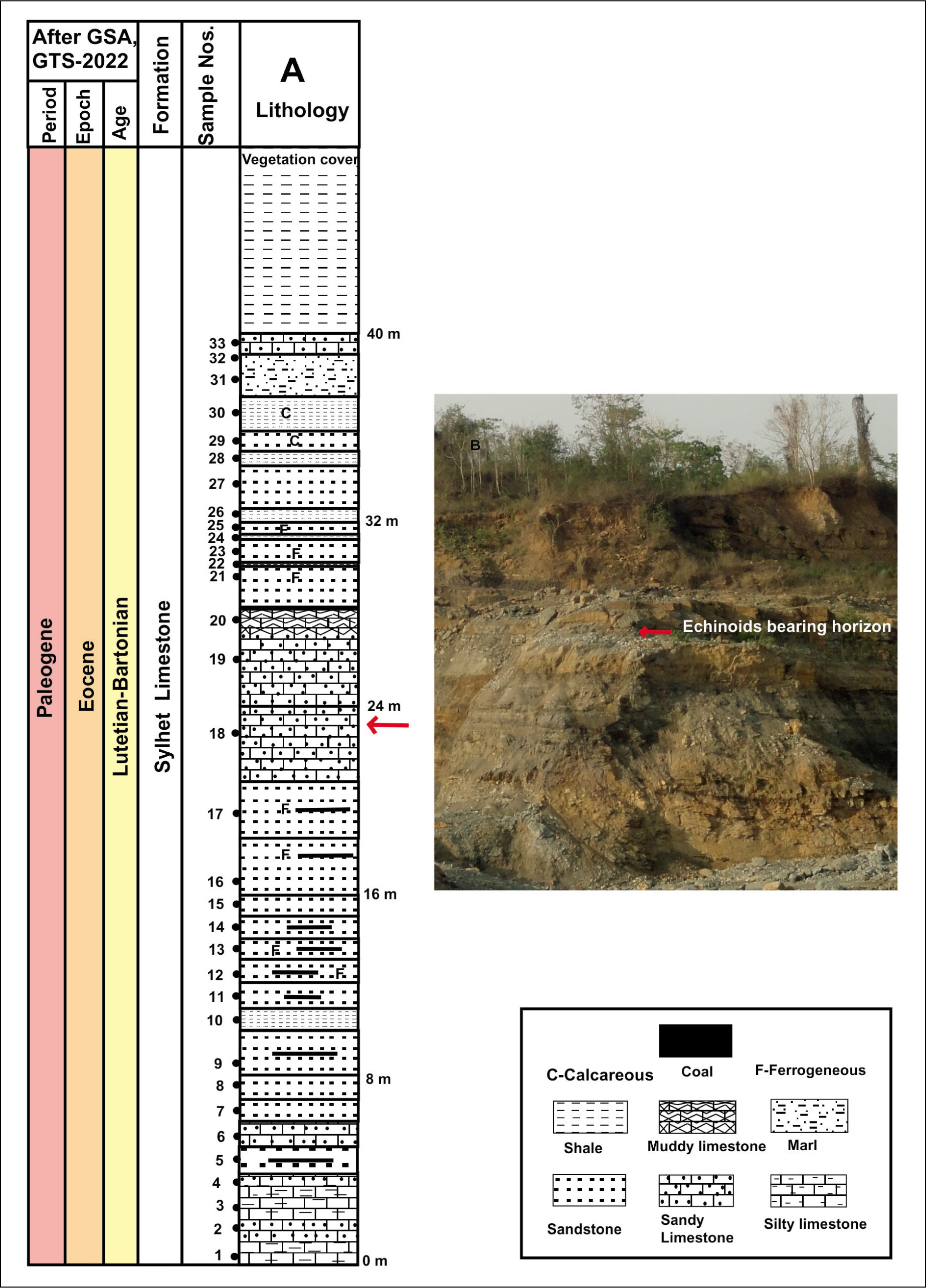
The Sylhet Limestone, which is exposed in the Mikir, Garo, Khasi and Jaintia Hills, was deposited in a shallow, open marine, warm water environment (Bhandari et al., 1973; Dutta, 1982). It consists of alternating limestones and sugary, white sandstones with mineable coal seams. The present study area is confined to the upper part of the Sylhet Limestone on the south-eastern edge of the Mikir Hills, which is of middle Eocene age (Biswal et al., 2021; Venkatachalapathy & Whiso, 2009). Previously, the Sylhet Limestone of Mikir Hills has already yielded invertebrate faunas (Foraminifera) and vertebrate faunal remains like shark, ray, crocodile, conical tooth and echinoderm spines (Biswal et al., 2021, 2022; Whiso et al., 2009). The specimens of Porocidaris schmidelii and Ilarionia sindensis have been recovered from a sandy limestone horizon in the studied section of sample Nos. 18 and 19 respectively (Figures 2A, B). The horizon containing the echinoid faunas (Plates 1 and 2) described here comes from larger benthic foraminiferal Zone SBZ 16–18, which corresponds to late Lutetian to Bartonian of middle Eocene age (Biswal et al., 2021).
(A) Measured litholog of the studied section at Dillai Parbat marking the echinoids yielding horizon; (B) Field photo of the outcrop with the echinoids bearing horizon.
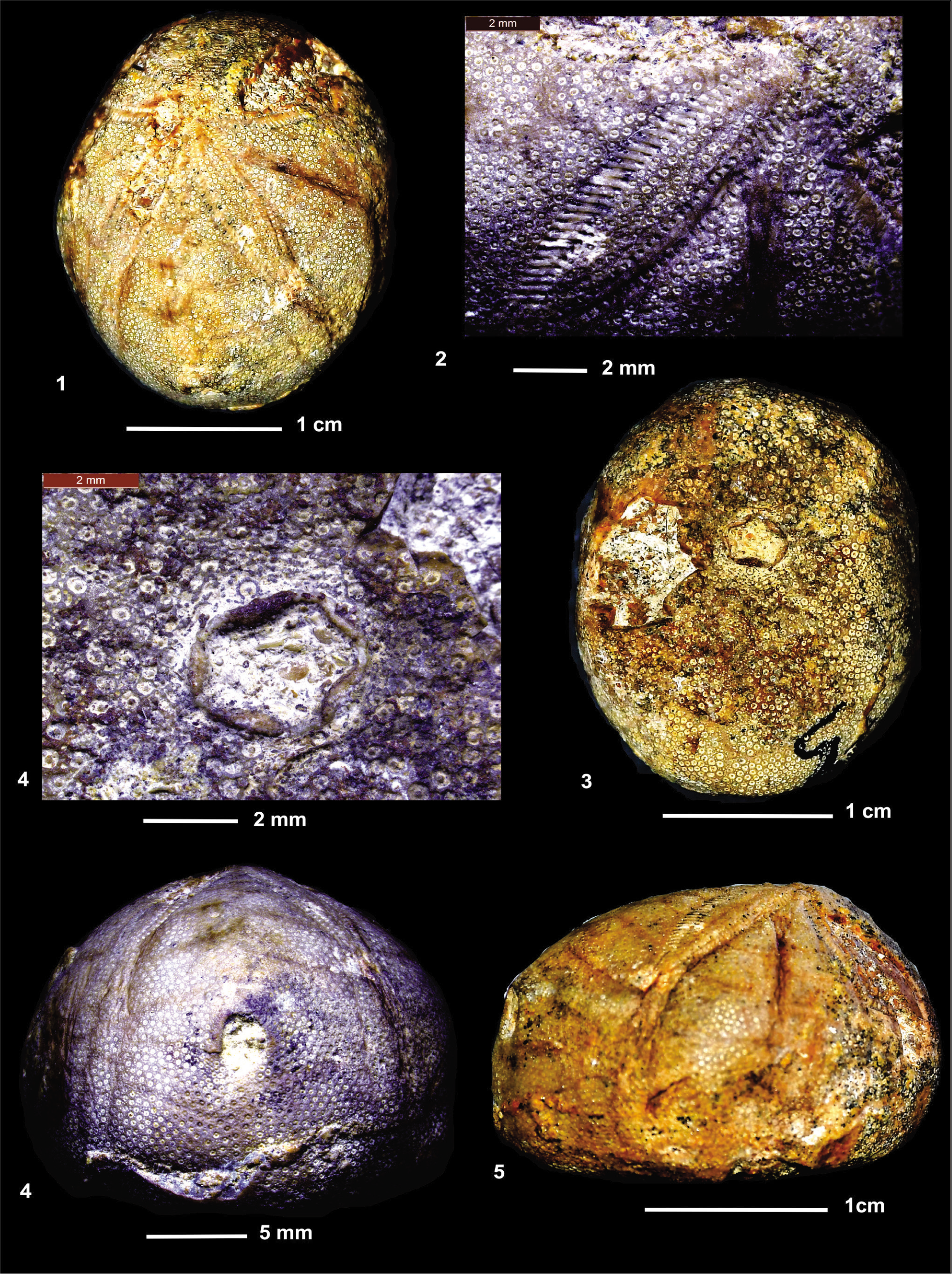
Porocidaris schmidelii. (1) Partial view of an ambulacrum and contiguous interambulacral areas; (2) Detail of interambulacral tubercles; (3) Detail of a primary interambulacral tubercle; (4) Detail of interambulacral surface; (5) Detail of an ambulacrum.

Ilarionia sindensis. (1) Apical view; (2) Petal I; (3) Oral view; (4) Oral area; (5) Posterior view; (6) Lateral view (anterior to right).
Repository and Institutional Abbreviations
Illustrated Echinoides specimens are curated in the WIHG, Dehradun repository bearing nos. WIMF/A4904-4905.
Systematic Palaeontology
The Kroh and Mooi classification (2023) is adopted.
Class ECHINOIDEA Schumacher, 1817
Subclass CIDAROIDEA Smith, 1984
Order CIDAROIDA Claus, 1880
Family RHABDOCIDARIDAE Lambert, 1900
Genus Porocidaris Desor, 1855
? 1830 Cidarites schmidelii; Münster in Goldfus, p. 120, pl. 40, figs. 4a,b [spines]
* 1881 Porocidaris schmidelii Desor (Münster); Loriol, p. 61, pl. I, figs. 1–15
1886 Porocidaris schmidelii Münster; Cotteau in Vidal, L.M., p. 80, Lám. IX.figs.12–14
1892 Porocidaris schmedlii (Münster) (sic); Cotteau, pl. 310, figs. 1–18
1901 Porocidaris schmidelii Münster; Oppenheim, p. 85
1970 Porocidaris schmidelii (Münster); Bodelle, et al., p.30, pl. h t., figs. 1–5
1992 Porocidaris schmideli (Münster, 1826); Roman et al. p. 75
v 2016 Porocidaris schmidelii (Munster, 1830); Llansana et al., p. 152, fig. De la p. 152
2017 Porocidaris schmidelii (Munster, 1830); Ali, p. 51, fig. 6 (9–11)
2018 Porocidaris schmidelii (Munster in Goldfuss 1830); Elattaar, p. 9, (Plate 1, Figs. 8–11°)
v 2021 Porocidaris schmidelii (Munster in Goldfuss 1830); Carrasco, p. 108
Material: Fragment of a specimen with inventory number WIMF/A4904 housed in the museum of Wadia Institute of Himalayan Geology (WIHG), Dehradun.
Remarks
Porocidaris Desor, 1855, was described from disarticulated interambulacral plates that presented a very peculiar characteristic: the areoles were adorned by deep elongated incisions in a radial position. When some isolated spines were found associated with these kinds of plates in the same bed, these spines were attached to the description of a specific species. Subsequently, many species of Porocidaris were described from single spines that exhibited small variations from the type species. This methodology, common in the nineteenth century, has generated many confusions. Therefore, in the ‘Comparison with other species’ and ‘Occurrence’ sections, only the citations based on the study of tests have been taken into account.
Diagnosis
Ambulacra straight with sunken pore zones and inner series of four small tubercles; the two innermost series with the smallest tubercles. Pore pairs conjugate and separated from each other by a wall. The inner pores are circular, and the outer ones are nearly oval. On each interambulacral, two series of primary tubercles with relatively small mamelons. Primary tubercles perforate, crenulate and about 12 indentations at the platform per main tubercle have been counted. Areoles with narrow, long and deep radial grooves. The areoles are tangent to each other, especially on the adoral surface.
Comparison with Other Species
The poor knowledge of the few species of this genus, many described only from plates, and others only from radioles, does not allow setting clear separation criteria. As a notable exception, it should be noted that the species Porocidaris anomala Duncan and Sladen (1884) from the middle Eocene of SE Pakistan, was meticulously described and figured. Porocidaris anomala is distinguished primarily from P. schmidelii by having much shorter areolar grooves on the interambulacral main tubercles.
Occurrence
The findings from the middle Eocene are many: NE Spain (Carrasco, 2021; Gàsser, 2000; Llansana & Romero, 2016), Biarritz in Southwestern France (Castex & Lambert, 1920), Angoumé in Southern Aquitaine-France (Roman & Cahuzac, 1992), Carinthia in Southern Austria (Collignon, 1930), Italian region of Veneto (Oppenheim, 1901), Provence and southern French Alps (Bodelle et al., 1970; Lambert, 1918), Persian Gulf (Roman, 1976), Oman (Roman & Cahuzac, 1992), Egypt (Ali, 2017; De Loriol, 1881; Elattaar, 2018) and from the Bartonnian-Priabonian range of Istria (Mikuž et al., 2014). The material studied herein represents the first record from the middle Eocene of India.
Subclass EUECHINOIDEA Bronn, 1860
Infraclass IRREGULARIA Latreille, 1825
Family GITOLAMPADIDS (nomen provisorium)
Genus Ilarionia, Dames, 1877
Ilarionia sindensis Duncan & Sladen, 1884, Plate 2, figs. 1–6
1884 Ilarionia sindensis Duncan & Sladen, p. 179, pl. XXXII, figs. 9–18
Material - One specimen with inventory number WIMF/A4905 housed in the museum of Wadia Institute of Himalayan Geology (WIHG), Dehradun.
Diagnosis
Small size test, elongated, inflated, rounded contour on the front, slightly narrower at the back, maximum width towards the middle of the test, the maximum height of the test passes through the apical system and this is in a forward position: the ratio between the distance from the apical system to the anterior edge and the length of the test is ca. 1/3. Relatively high test: H/L = 0.68 mm. Aboral surface is nearly flat, slightly sloping backwards and strongly sloping forwards. Petals lanceolate and closed. Posterior petals are 15% longer than the anterior ones. Petals paired (I, I, IV and V) relatively wide: the ratio between their maximum width and their length is ca. 1/3. The anterior petal (III) is a little narrower than the others. The interporous zone is at least twice as wide as the poriferous zone. Pore-pairs conjugate, the internal ones are circular and the external ones are nearly oval. Ambulacral and interambulacral areas are covered by small tubercles of equal size, all with depressed areoles. Tubercles on the adoral surface are slightly larger and a few more widely spaced. Apical system anterior: The ratio between the distance from the peristome to the anterior edge and the length of the test is ca. 0.40. Transverse peristome, pentagonal outline, bordered by a protruding ridge. Periprocte is longitudinally oval; its length is nearly less than 1/5 of the height of the test, located in the centre of the posterior face; the ratio between its width and length is 2/3.
Length (L): 25.565 mm; width (W): 20.155 mm; and height (H): 17.475 mm.
Remarks
Duncan and Sladen (1884) recognised that the species had great variability in the following characteristics: position of the apical system, outline and profile of the test, and the outline of the peristome.
Comparison with Other Species
The closest species is Ilarionia beggiatoi (Laube, 1868) from the middle Eocene of Northern Italy, NE Spain and Croatia (Carrasco, 2016). However, I. sindensis has the apical system relatively more anterior, and I. beggiatoi is almost central. Also, in I. sindensis the posterior petals are relatively shorter than the anterior ones, and in I. beggiatoi they are relatively longer.
Occurrence
The species has been found in the middle Eocene of the Sindh Region (Pakistan) by Duncan and Sladen (1884) and a variety (I. sindensis var. madagascariensis Cottreau, 1935) in the middle Eocene of Madagascar. Herein it has been found in the middle Eocene of Mikir Hills, Assam, India and it represents the first record from India.
Discussion
The present finding brings a new palaeontological data set to the region. As most of the echinoids have robust tests that fossilise well, almost all the characters and environmental constraints which one can determine for extant groups can also be found in fossil forms (Osborn et al., 2016). Due to their ability to fossilise well, they are more common and better preserved than other types of fossils in many deposits and are normally used for biostratigraphy. Echinoderms first appeared in the fossil record in the Cambrian and rapidly diversified into many groups but were not successful in the Ordovician as were dominated by the crinoids. In the Triassic Period, no echinoids were recorded. In the Jurassic Period, rare occurrences were recorded initially, and later with much diversification; they remained successful till date (Srivastava, 2014). The lack of diversity and the time of diversification during the Triassic and Jurassic were explained by the reassembly of Laurasia by Valentine (1970) in Srivastava (2014). According to them, the environmental conditions were not favourable for growth and development of the echinoids as the Indian Plate was situated nearer to the South Pole, away from the equator. The breaking of the Indian plate from the Gondwanal and during the late Jurassic-early Cretaceous and its northward flight towards the equator diversified the echinoids during the Cretaceous in the Indian subcontinent. Moreover, according to Mayr (1954) and Kier and Grant (1965), most echinoids species are geographically restricted to some extent by geographical barriers, oceanic currents, nature of substratum, hydraulic regime, predation, salinity temperature, food availability and depth. Echinoids are reported to have appeared in the United Kingdom (England and Scotland) and in the USA during the Ordovician Period (Mannil, 1962; Smith & Savill, 2001), but in India and Pakistan, the earliest report of echinoids is from the Permian rocks of Karakoram and the Salt Range in Pakistan and shows a European affinity (Srivastava, 2006). In India, Eocene echinoids are recorded from Lairyngao in Meghalaya by Medlicott (1869); the Harudi Formation and Fulra limestone in the southwestern part of Kachchh by Srivastava and McNamara (2011); Jammu of Himalaya by Srivastava et al. (1990). Recently, Iangrai et al. (2022) reported and described late Cretaceous-Early Palaeocene echinoids from the Meghalaya shelf in the vicinity of the K-Pg mass extinction boundary.
Palaeoenvironmental analysis based on the sedimentological observations, foraminifera (Biswal et al., 2021), fossil fish assemblages (Biswal et al., 2022) and mammalian lumbar vertebra (Whiso et al., 2009) from previous studies and echinoids from the present study suggest a warm, shallow marine environment ranging from open marine, inner neritic to more proximal coastal settings. Further, the present finding of Eocene echinoids from Mikir Hills, in addition to its earlier records from Kachchh, Rajasthan and Shillong Plateau, indicates the possible existence of a marine seaway connecting northeastern and western sectors facilitating dispersal of the benthic fauna. Also, it shows evidence that the Neotethys Sea was connected through Biarritz (Southwestern France), Angoumé (Southern Aquitaine, France), NE of Spain, Carinthia (Southern Austria), Venetian region of Italy Provence, Southern Alps of French, Istria (Croatia), Persian Gulf, Oman, Egypt, Sindh province of Pakistan and Madagascar during the middle Eocene. Future studies in this region will be carried out to reveal more new records and to fill in the gap of the species list as well as the geographical distribution of echinoids.
Conclusion
This finding is the first report of middle Eocene echinoids from the Sylhet Limestone of Mikir Hills, Assam.
The two echinoid taxa, that is, P. schmidelii and I. sindensis are also reporting for the first time from India and it brings new data set to the region and imply a warm, shallow marine environment of deposition, also as indicated by foraminifera and remains of sharks and rays in the previous investigation carried out in the study area.
The area of investigation indicates a palaeontological significance.
Present record of echinoids from the Mikir Hills and previous records of echinoids from other parts of India suggest a marine seaway connection between northeastern and western sectors. Also, it shows evidence that the Neotethys Sea was connected through Biarritz (Southwestern France), Angoumé (Southern Aquitaine, France), NE of Spain, Carinthia (Southern Austria), Venetian region of Italy Province, Southern Alps of French, Istria (Croatia), Persian Gulf, Oman, Egypt, Sindh province of Pakistan and Madagascar during the middle Eocene.
Footnotes
Acknowledgements
The authors acknowledge Dr Kalachand Sain, Director of Wadia Institute of Himalayan Geology, Dehradun for providing the facilities and permission to publish this research work (WIHG/Publication no. 0264). Authors sincerely thank the authorities CCI Mines, Bokajan Karbi Anglong District, Assam for the permission to carry out the fieldwork. Mr. Loli Hekha, Kapani Khoziio and Adahrii Adakho are sincerely thanked for the help extended in the field. KL and SB sincerely express thanks to Sanjeev Dabral (Senior Technical Officer), Nain Das (Lab. Asst.), Ravi Lal (Lab. Attendant) of Biostratigraphy Group of WIHG, Dehradun for the various help. Authors also gratefully acknowledge Dr Rajani Panchang, Convener 28th ICMS-2022, SPPU, Pune and Guest Editor, Special Issue JPSI for the invitation to contribute to this article. We are thankful to Dr Sebastián Calzada, Director of the MGSB and Dr Pedro Adserà, Curator of the invertebrate palaeontological collection of MGSB, Spain for reviewing the manuscript, which has helped improve it immensely.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors received no financial support for the research, authorship and/or publication of this article.