
Abstract
Pycnodus was among the fish genera that successfully survived the Cretaceous-Palaeogene (K-Pg) mass extinction event. It was the longest existing and most adaptable taxon within the entire group of pycnodonts. The present study highlights the finding of pycnodont remains (prearticular dental plate and isolated teeth) from the Khuiala Formation (Early Eocene, Ypresian) and the Bandah Formation (Middle Eocene, Bartonian), Jaisalmer Basin, Rajasthan. The specimens from the Khuiala Formation are assigned to Pycnodus toliapicus, and those from the Bandah Formation are assigned to Pycnodus sp. The record of Pycnodus toliapicus is particularly significant as it is the only species from India based on a dental plate. It signifies the presence of pycnodonts in the seas east of the Tethys just before they got completely extinct worldwide.
INTRODUCTION
The pycnodonts represent a group of fossil fish that thrived for nearly 200 Ma before going completely extinct. They first appeared around the Ladinian (late Middle Triassic) and survived until the Priabonian (Late Eocene) (e.g., Cawley et al., 2021; Kriwet, 2001; Nursall, 1996; Poyato-Ariza & Wenz, 2002; Stumpf et al., 2017). During this interval, pycnodonts played a significant role in the marine vertebrate communities and distribution worldwide. Pycnodonts thrived mainly in marine and brackish environments although a few occurrences are also known freshwater habitats (Kriwet, 2001, 2005; Longbottom, 1984; Martín Abad & Poyato-Ariza 2013; Stumpf et al., 2017). Many pycnodonts lived in shallow reef environments with swimming adaptations common to reef fish (Nursall, 2010).
The pycnodonts have been well documented from the Indian Late Cretaceous to Palaeogene horizons. Notable Palaeogene pycnodont occurrences include discoveries from Subathu Formation (Kumar & Loyal, 1987) and the Palaeogene of Barmer Basin (Fatehgarh, Akli, and Kapurdi formations) (Rana et al., 2005, 2006). Some pycnodont remains have also been recorded from the Early Eocene Khuiala Formation (Kumar et al., 2007). In the recent year, the Jaisalmer Basin attracted attention after the discovery of diverse vertebrate fossils, including those of early whales from the Bandah Formation of Middle Eocene age (Kumar et al., 2017, 2020). Subsequently, elasmobranchs from the Bandah Formation were also described by Rana et al. (2021).
The present study provides the first detailed description of a dental plate of a pycnodont from the Khuiala Formation along with isolated pycnodont teeth from the Bandah Formation. These newly described specimens are significant as they allow the first ever taxonomic identification of pycnodonts from the Bandah Formation of the Middle Eocene age.
GEOLOGICAL SETTING OF THE STUDY AREA
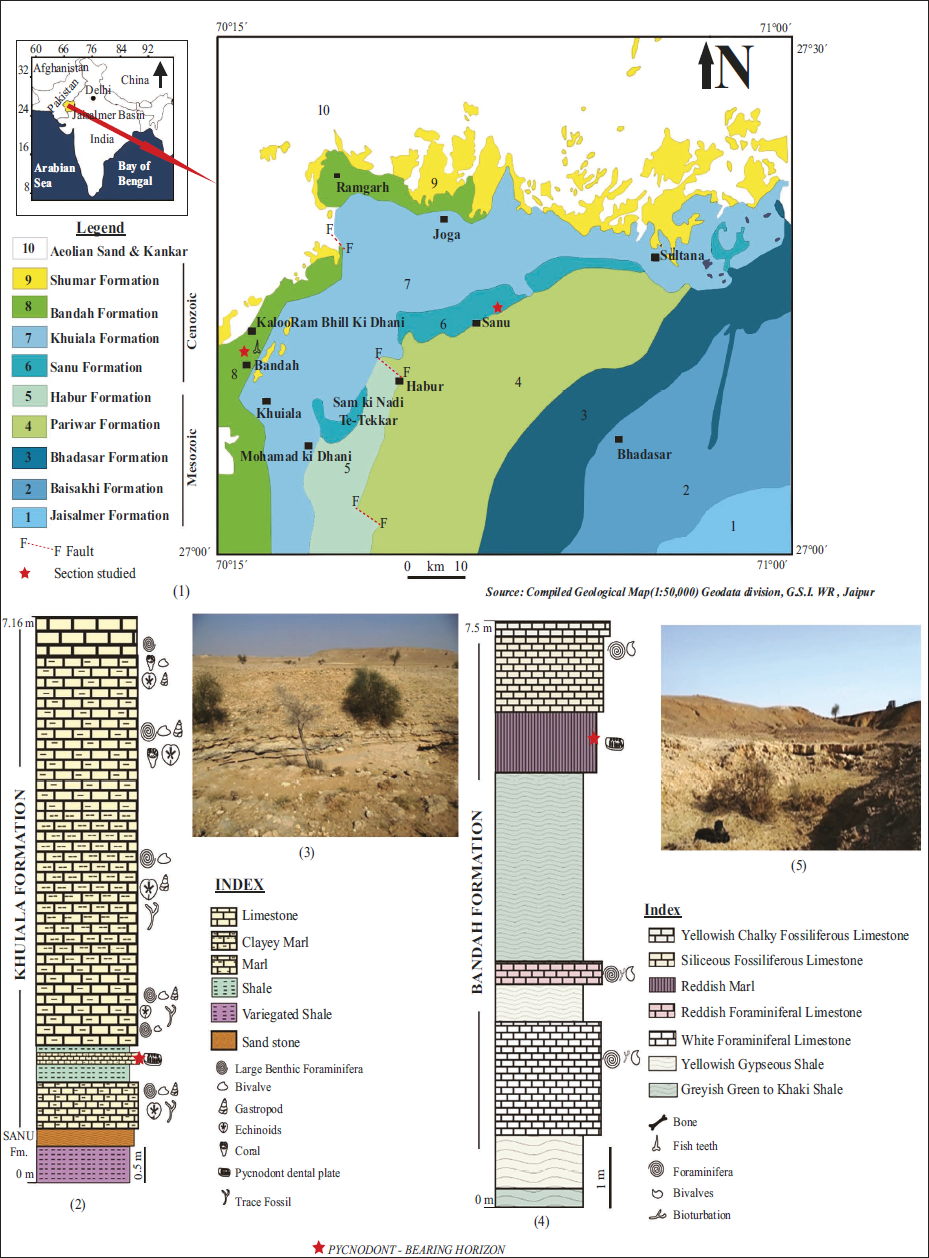
The Jaisalmer Basin is a pericratonic basin situated in the western Part of Rajasthan. It represented the eastern extension of the Indus Basin and forms a significant portion of the “West Rajasthan Shelf” tectonic province, lying west of the Aravali ranges. This area includes several sedimentary basins, namely Barmer, Jaisalmer and Bikaner-Nagaur, spanning approximately 30,000 square kilometres (Singh, 2007). Among all the sedimentary basins, the Jaisalmer Basin preserved almost complete records of the Mesozoic and Cenozoic sedimentary succession. The Cenozoic, particularly, the Palaeogene successions of the Jaisalmer Basin, is richly fossiliferous and deposited under shallow marine environment, with age ranges from Palaeocene to Middle Eocene (Dasgupta, 1974, 1975; Pal et al., 2007; Pareek, 1984; Singh, 1996, 2003). This sedimentary succession of the Jaisalmer Basin is divided into Sanu, Khuiala and Bandah formations in the stratigraphic order. The fossiliferous Khuiala Formation (Early Eocene) has a gradational contact with the underlying Sanu Formation (Palaeocene). The Middle Eocene Bandah Formation, mainly composed of argillaceous and carbonate horizons. The Khuiala and Bandah formations are known to yield various invertebrate fossil assemblages, for example, foraminifer, ostracods, molluscs, echinoderms etc. (Bhandari, 1996; Habibnia & Mannikeri, 1990; Khosla, 1973; Sigal et al. 1971; Singh, 1984, 2007; Srivastava & Kulshreshtha, 2009). Compared to invertebrates, the vertebrate records of these two Eocene horizons are remarkably rare, with comparison to other Palaeogene sedimentary basins of Rajasthan (e.g., Barmer and Bikaner basins). In the Bikaner Basin, fishes were described from the Marh and Palana Formations of Palaeocene-Eocene age (Jolly & Loyal, 1985; Kumar et al., 2005; Paliwal 1999). In the Barmer Basin, vertebrates (mainly fishes) were reported from the Fatehgarh Formation, Akli Formation and Kapurdi Formation of Palaeocene-Eocene age (Barooah, 1950; Rana et al., 2005, 2006; Sahni & Choudhary, 1972, 1975; Tiwari, 1968). However, in the Jaisalmer Basin, Early Eocene fishes were described from the Khuiala Formation by Kumar et al. (2007). Recently fishes, reptiles and mammals have been reported from the Bandah Formation of Middle Eocene age (Kumar et al., 2017, 2020; Rana et al., 2021).
The pycnodont yielding horizons and age of the studied sections
The pycnodont fish remains have been recorded from the two stratigraphic successions of Palaeogene of the Jaisalmer Basin. One is from the Khuiala Formation, and the other is from the Bandah Formation. From the Khuiala Formation, the pycnodont-yielding section is exposed at 3 km NW of Sanu village, situated along Sanu-Ramgarh Road (N 27° 10’ 25.31”; E 70°28 ’26.55”). This section exposes ~7.16 m thick sedimentary sequence of the Khuiala Formation (Figures 1.1–1.3). Lithologically, the Khuiala Formation consists of yellowish to greenish shale and yellowish marl beds and is capped by whitish limestone. Fossils of bivalves, solitary corals, and foraminifers were documented in the Khuiala Formation (Kumar et al., 2007 and reference therein). The pycnodont specimen described in this study was recovered from this section’s lowermost yellow marl band.
The Early Eocene (Ypresian) age of the Khuiala Formation was previously established based on the presence of the index larger foraminifer Nummulites burdigalensis, as documented by Singh (1996, 2003). N. burdigalensis corresponds to Shallow Benthic Zone (SBZ) 10 and Planktic Zone P8, which designate an Early Eocene (Ypresian) age, as delineated by Serra-Kiel et al. (1998). Additionally, the ichthyofauna documented by Kumar et al. (2007), further supports the assignment of Early Eocene (Ypresian) age of the Khuiala Formation.
The second pycnodont-bearing horizon reported here is a part of the Bandah Formation. This nala-cutting section is located approximately 2 km northeast of the Bandah village, along the Asutar-Bandah-Ramagarh Road (N 27°09ʹ0.3”; E 70°28ʹ08.8”; Figures 1, 1.4–1.5). This section exposes a ~7.5 m thick sedimentary sequence of Bandah Formation, beginning with gypseous bentonitic clays (at the base) followed by whitish to yellow marl, fossiliferous limestone, greyish-green colour shale, and siliceous marl. The section is capped by Discocyclina bearing marly siliceous limestone. Several isolated pycnodont teeth have been recovered from the greyish green colour shale and marl unit in the upper part of this section (Figure 1.4).
The Middle Eocene age to the Bandah Formation have been assigned based on age diagnostic benthic foraminifera (e.g., Dictyoconoides cooki, Discocyclina javanavar indica, D. sowerby Nummulites acutus and Assilina spira etc.) and planktonic foraminifera (Globigerina kakugleri, Truncoroloides topilensis, Turborotalia cerroazulensis and Truncorotaloide srohri) by earlier researchers (Raju, 2018; Saraswati et al., 2000; Sigal et al., 1971; Singh, 1984, 1998, 2007). Based on these foraminiferal records, the Bandah Formation belongs to the Orbulinoides beckmanni Zone, which is restricted to the Middle Eocene Zone E 12 (=P 13) of Bartonian age (Premoli Silva et al., 2006; Raju, 2018; Singh, 1998). The calcareous nannofossil records were also assigned a Middle Eocene (Bartonian) age to the Bandah Formation (Rai et al., 2014; Singh, 1998). Based on the diagnostic benthic and planktic foraminifera and calcareous nannofossils, a Middle Eocene (Bartonian) age may be confidently assigned to the pycnodont-yielding horizons of the Bandah Formation.
MATERIALS AND METHODS
A careful examination of the surface outcrop resulted in the recovery of the described specimens under the present study (Figures 1.3 & 1.5). The fossils specimens were prepared by a mechanical method in the laboratory, and photography was conducted using a NIKON D5300 digital camera. All the described specimens in the present study have been reposited in the Vertebrate Palaeontology Division of the Geological Survey of India, Jaipur, Rajasthan. The pycnodont fossil samples are catalogued as RWR numbers and recorded under RWR/Khuiala/Vertebrates/Rajasthan/GSI/WR and RWR/Bandah/Vertebrates/Rajasthan/ GSI/WR.
SYSTEMATIC PALAEONTOLOGY
Class: Osteichthyes
Infraclass: Actinopterygi
Order: Pycnodontiformes
Family: Pycnodontldae (Agassiz, 1833)
Genus: Pycnodus (Agassiz, 1833)
Species: Pycnodus toliapicus, (Agassiz, 1833)
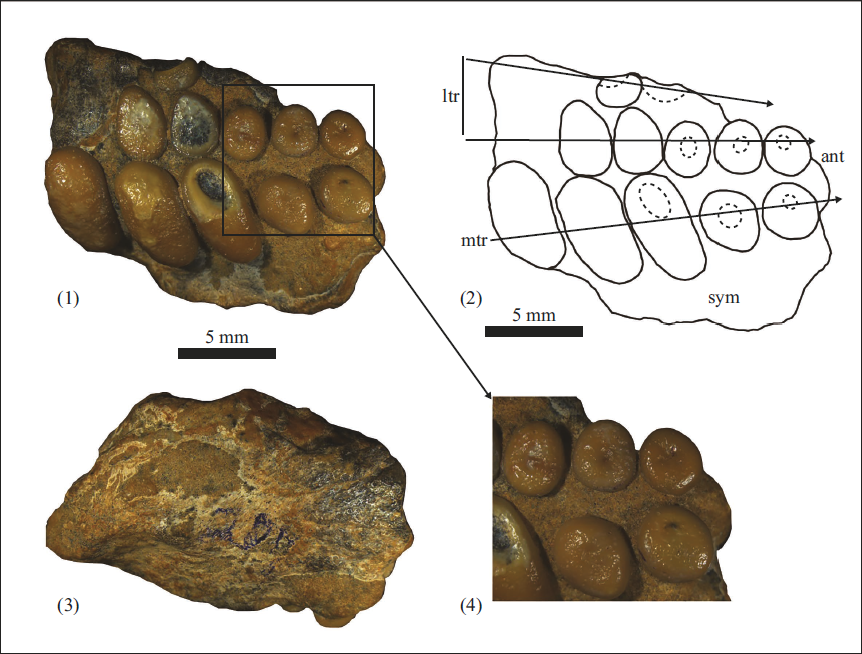
(Plate-1, Figures 1–4)
1–15, isolated molariform teeth of a prearticular or vomer teeth of Pycnodus sp. from Bandah Formation, Jaisalmer (RWR/B/87-RWR/B/93).
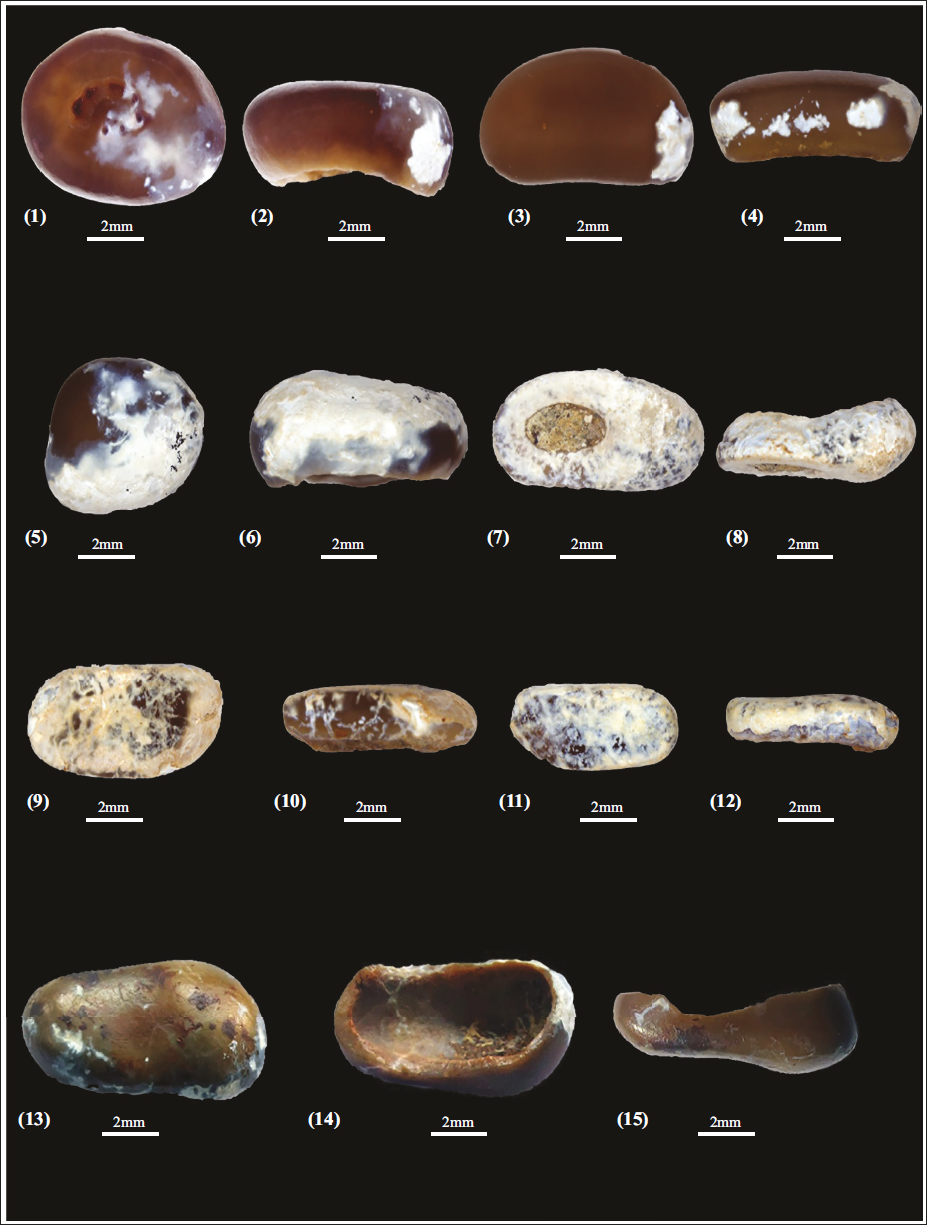
1–12, isolated branchial teeth (RWR/B/94-RWR/B/99); 13–16, isolated incisiform teeth (RWR/B/100-RWR/B/101) teeth of Pycnodus sp. from Bandah Formation, Jaisalmer.

Referred material and Horizon
Single specimen of prearticular tooth plate (RWR/K/86) from yellow marl of the Khuiala Formation, Jaisalmer Basin, Rajasthan.
Description
The present specimen, comprises a single, partial leftward prearticular tooth plate, containing three rows of teeth: A primary row arranged on the symphyseal side, and two lateral prearticular tooth row on the outer side. The second lateral row preserves only one tooth crown and two sockets as a result of breakage. The preserved portion of the main row contains five rows of teeth, with three being elongated and two teeth located anteriorly exhibiting a rounded to sub-rounded shape. The size of the elongated teeth varies from 8 mm to 10 mm. The occlusal surface of the preserved teeth shows a pitted structure on the first and second elongated (bean-shaped) teeth on the posterior side. The third elongated teeth towards the anterior side shows an erosional pit or depression due to erosion. The anterior-most rounded to sub-rounded teeth of the median row show shallow pit marks with wrinkles on their surface.
The size of the primary row teeth reduces their size towards the front (4.9 mm–10.1 mm in length and 4.5 mm long and 5 mm in width) with 0.61 is L/W (length-width). The teeth exhibit elliptical to bean-shaped outlines along both the medial and lateral margins, with the lateral margins being more elongated. The teeth’s front edge (mesial) in the central row is convex, while the rear (distal) edge is straight. These primary row teeth are bigger, while the lateral and medial teeth are small in size. The gaps between the main teeth are very less. On the surface of both, the posterior teeth of the primary row show pitted ornamentation, and it is absent on the third tooth, which may be due to the abrasion. The apical furrow can be seen on the surface of the lateral and medial rows of teeth. Five (05) tooth crowns are present in the first lateral tooth row. The medial tooth is elongated in shape and bigger than the from tooth (anterior), with the average length/width ratio is 0.70. Only a single, incomplete tooth crown and a very small tooth socket are preserved on the second lateral tooth row.
Remarks
This is the first record of a prearticular dental plate of pycnodont from the Early Eocene Khuiala Formation of Jaisalmer Basin, assigned to P. toliapicus. In the eastern Tethys region, including the Asian sea, Pycnodus constitutes the predominant taxon within the Palaeogene pycnodont assemblages (Capasso, 2021) and is generally distributed in shallow marine environments (Cawley et al., 2018). The systematic identification of pycnodont fishes, based exclusively on isolated teeth, is inherently challenging due to considerable variations in the molariform tooth morphology. While isolated dentitions can provide reasonably precise identifications, para-taxonomic issues may arise when taxa are described exclusively from such remains. Nevertheless, distinctive combinations of dental features often enable reliable diagnoses of new taxa (Cooper & Martill, 2020; Kriwet, 2005; Stumpf et al., 2017; Vullo et al., 2017). In certain instances, the autapomorphic characters may also be expressed within the isolated tooth plates. Nonetheless, the taxonomic identifications are most reliably established by examining articulated specimens, which further offer a more complete anatomical context (Kumar et al., 2022).
The Indian record of the Pycnodus is from the Deccan Infra- and Intertrappean beds of peninsular India (Bajpai et al., 1990; Gayet et al., 1984; Mohabey & Udhoji, 1996; Prasad, 1989; Verma et al., 2024); Kakara and Subathu formations (Palaeocene–Eocene) of the sub-Himalayan region (Kumar, 1996; Kumar & Loyal, 1987).
The earliest record P. toliapicus is documented from the Late Palaeocene (Thanetian) deposits of Nigeria, Togo and Niger (Kogbe & Wozny, 1979; Longbottom, 1984; White, 1934). Subsequent records include the Early Eocene (Ypresian) and Middle Eocene (Lutetian) sequences of England, Middle Eocene (Lutetion) deposits of Paris Basin and Belgium (Casier, 1950; Savornin, 1915; Taverne & Nolf, 1978), as well as the Eocene sediments of Qatar in the Persian Gulf region (Casier, 1971). This species (P. toliapicus) was first documented in India in the Himalayan region (Kumar & Loyal, 1987). Additional pycnodont species, including P. subathunensis, P. dattai, P. bicresta, P. lametae were also described from the Subathu Formation of Eocene age (Kumar & Loyal, 1987; Prasad & Singh, 1991; Thappa et al., 1998). Based on the general resemblance in form and dentition arrangement to P. toliapicus described by Longbottom (1984) and isolated molariform teeth described by Kumar and Loyal (1987), the Jaisalmer specimen best fits P. toliapicus. This identification is based on the presence of a circular occlusal outline characterised by a cluster of cusps encircling a central depression. Additionally, the Jaisalmer prearticular specimen exhibits shallow pit marks and surface wrinkles in the main row’s frontside (anterior) teeth. The posterior teeth in the main row feature pitted ornamentation, except for one where abrasion likely obscures these features. In size and dental morphology, P. toliapicus differs from other taxa such as P. dattai, P. subathunensis, and P. bicresta. It is larger and possesses more globular main-row teeth with a convex posterior margin, unlike the transversely wider and straight posterior margins observed in the Jaisalmer teeth. Despite some uncertainty in the ornamentation pattern of the Jaisalmer prearticular teeth, its overall morphology, including five tooth rows, closely corresponds with P. toliapicus. Interestingly, hook-shaped teeth previously identified as Stephanodus lybicus from the Khiuala Formation (Kumar et al., 2007) have been assigned to the branchial teeth of pycnodonts (Kriwet, 1999; Poyato-Ariza & Wenz, 2002, 2005).
Age
Early Eocene (Ypresian age) (Kumar, 2007; Serra-Kiel et al., 1998; Singh, 1996, 2003, 2007).
(Plate-2, Figures, 1–15; Plate-3 Figures, 1–16)
Referred material and Horizon
Seven specimens of isolated molariform teeth of a prearticular or vomer (RWR/B/87-RWR/B/93), six specimens of isolated branchial teeth (RWR/B/94-RWR/B/99), and two specimens of isolated incisiform teeth (RWR/B/100-RWR/B/101) from the greyish green colour shale and marl units, Bandah Formation.
The isolated molariform teeth are medium to large (10 mm–26 mm) and belong to the prearticular or vomer; the teeth are elongated to subcircular in shape, the base is hollow, and minute wrinkles or protuberances mark the occlusal surface. The largest tooth is 23 mm and may belong to the lateral splenial tooth of the main row; the occlusal surface is smooth. The isolated branchial teeth (Plate 3, Figures 1–12) are small in size (6 mm–11 mm); teeth exhibit a flattened, compressed form, featuring a pronounced hook-like principal cusp with a basal shelf-like extension, a subordinate cusp and a well-developed, deep root. The isolated incisiform teeth (Plate 3, Figures 13–16) are characterised by a broader crown (9 mm–14 mm); principal (main) cusp, arched and translucent, displaying a convex outer surface and a concave inner surface, along with a smooth, unornamented cutting edge. A subtle, poorly developed ridge links the secondary cusp to the main cusp.
Remarks
The Bandah pycnodont material includes isolated molariform teeth from a prearticular or vomer, branchial and incisiform teeth. The similar isolated teeth were recorded from the Intertrappean beds of Asifabad and assigned to Pycnodus sp. by Prasad and Sahni (1987). Hook-shaped teeth, previously identified as Stephanodus lybicus, have recently been found within the branchial chambers of pycnodont fishes (Kriwet, 1999; Poyato-Ariza & Wenz, 2002, 2005). Although branchial teeth are also found in other fish groups, for example, Semionotiformes (Prasad et al., 2004; Thies & Mudroch, 1996), their crown morphology distinguishes them from the Eocene forms. Additionally, the frequent associations of these branchial teeth with both incisiform and molariform elements characteristic of pycnodontiforms, in conjunction with the absence of semionotiform remains in the Middle Eocene ichthyofaunal assemblages of the Jaisalmer beds, provide compelling evidence for their attribution to pycnodontiform fishes.
The present collection of pycnodont remains (isolated prearticular or vomer, molariform teeth, branchial teeth and incisiform teeth) from the Bandah Formation cannot be confidently assigned to any previously named species (e.g., P. lametae, P. bicresta, P. toliapicus). Thus, they are designated as Pycnodus sp.
Age
Middle Eocene (Bartonian) (Premoli Silva et al., 2006; Rai et al., 2014; Raju, 2018; Singh, 1998, 2007).
DISCUSSION
This study represents the first documented occurrences of Pycnodus sp. from the Bandah Formation (Middle Eocene), Jaisalmer Basin, Rajasthan. The pycnodonts are extinct non-teleostean actinopterygian fish. The pycnodont fishes first appeared during the Late Triassic period (Norian) and completely disappeared during the end of the Middle Eocene (Lutetian). The pycnodont were distributed worldwide, especially in the western Tethys, with maximum records from Central and South Europe (Martín-Abad & Poyato-Ariza, 2013). The decline of pycnodont fishes began in the latest Cretaceous, culminating in their extinction during the Eocene. The peak of pycnodont in the taxonomic diversity occurred during the early Late Cretaceous (Nursall, 1996; Poyato-Ariza, 2013). All the known Palaeogene pycnodonts to date were restricted to a few closely related lineages within the family Pycnodontidae, which exhibit limited taxonomic and ecomorphological diversity (Poyato-Ariza, 2005; Poyato-Ariza et al., 2013). Among these, Pycnodus stands out as the most prominent genus in the pycnodont assemblages during the Palaeogene Period, with a widespread distribution in shallow marine environments globally. The pycnodonts experienced a significant resurgence in the Tethys Sea during the Palaeocene, marking a notable adaptive radiation in the Lower Eocene. Their fossil records indicate a remarkable capacity to adjust to the new environmental conditions that emerged following the Cretaceous-Paleogene extinction event. A thriving population of pycnodonts persisted in the southern regions, with numerous findings reported from areas immediately south of the Tethys, including Morocco, Algeria (Longbottom, 1984; Savornin, 1915), Tunisia, and Egypt, as well as further south in Niger, Angola, Togo, and Nigeria. Notably, several important discoveries have been made in Egypt, including the most recent Eocene Pycnodont identified (Capasso, 2021) Pycnodus mokattamensis from the Priabonian. Furthermore, the colonies of pycnodonts that expanded into the seas of the eastern Tethys remained well-established throughout the Lower Eocene, with reports from Qatar and Saudi Arabia, and extending to Pakistan and India (Capasso, 2021).
CONCLUSION
The discovery of pycnodonts from Jaisalmer is consistent with their expansion during the Eocene in Tethys just before they became extinct. The record of pyconodonts from Jaisalmer helps to understand the palaeobiogeographic distributions of the Eocene pycnodont fish. The present record of the pycnodont from the Jaisalmer Basin holds the potential to bridge existing gaps in the stratigraphic records of the pycnodont family during the Palaeogene.
Footnotes
Acknowledgement
The authors gratefully acknowledge the anonymous reviewers for their constructive comments and suggestions, which helped improve the manuscript’s quality. Krishna Kumar expresses sincere gratitude to the Director General of the Geological Survey of India for providing an opportunity to publish this research work. The author extends his deepest respect and heartfelt gratitude to Prof. Sunil Bajpai, Indian Institute of Technology, Roorkee, for his meticulous guidance and invaluable support for the successful completion of this study. The authors sincerely thank the Additional Director General & HOD, Western Region, Jaipur, for providing departmental facilities and timely support. The authors also thank Additional Director General & HOD, Eastern Region, for constant guidance and motivation.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: Government of India (The research article is the outcome of the approved Field Season programme of the Geological Survey of India, Proposal ID: M4CRP/NC/WR//2019/28894 for the FS year 2019–2021.).