
Abstract
The graphoglyptid ichnofossil Paleodictyon (Meneghini, 1850, Osservazioni stratigrafiche e paleontologiche concernenti la geologia della Toscana e dei paesi limitrofi) is documented for the first time from the Early Eocene Flysch of the Andaman Islands. I review all known Indian occurrences of the ichnogenus Paleodictyon, including one from the Indian Ocean. I further argue that the modern Chinese checkerboard-like hexagonal pinholes from the deep-sea of the world’s major oceans, often considered as the extant P. nodosum, differ markedly in ecological and preservational context and therefore represent a distinct ichnogenus and ichnospecies rather than actual ‘living fossils’.
INTRODUCTION
The graphoglyptid Paleodictyon remains an enduring enigma regarding its function, a highly ordered, geometric trace fossil first described from the Eocene flysch of the Apennines, Italy (Meneghini, 1850). Paleodictyon consists of thin tunnels or ridges that typically form hexagonal or polygonal honeycomb-like networks. Both regular and irregular meshes occur throughout its stratigraphic range. However, it is the strikingly regular honeycomb patterns in forms such as P. carpathicum and P. nodosum that make the ichnogenus particularly notable and widely studied. Several ichnospecies of Paleodictyon (Meneghini, 1850) are recognised based on mesh size, regularity, and burrow architecture. The type species, Paleodictyon strozzii (Meneghini, 1850), exhibits a well-developed hexagonal network from the Italian flysch. Paleodictyon nodosum (Seilacher, 1977), the most widely known ichnospecies, has meshes ~1–3 cm. Several specimens previously assigned to Paleodictyon and its various ichnospecies have recently been reevaluated and reassigned to the ichnogenus Megagrapton (Uchman et al., 2022). Graphoglyptid trace fossils display distinct evolutionary trends in diversity, frequency, and structural complexity, reflecting adaptations to changing deep-sea environments. Uchman (2003) emphasised their palaeoenvironmental significance, noting that variations in network geometry and abundance correspond to ecological strategies, benthic conditions, and substrate consistency, offering key insights into ancient marine ecosystems.
In this study, author describes Paleodictyon from the Early Eocene of the Andaman Islands and review previous records of this ichnogenus from India. I also critically evaluate recent reports of Chinese checkerboard-like structures from the deep-sea, which have traditionally been assigned to P. nodosum.
SECTION DETAILS AND PALEODICTYON MORPHOLOGY
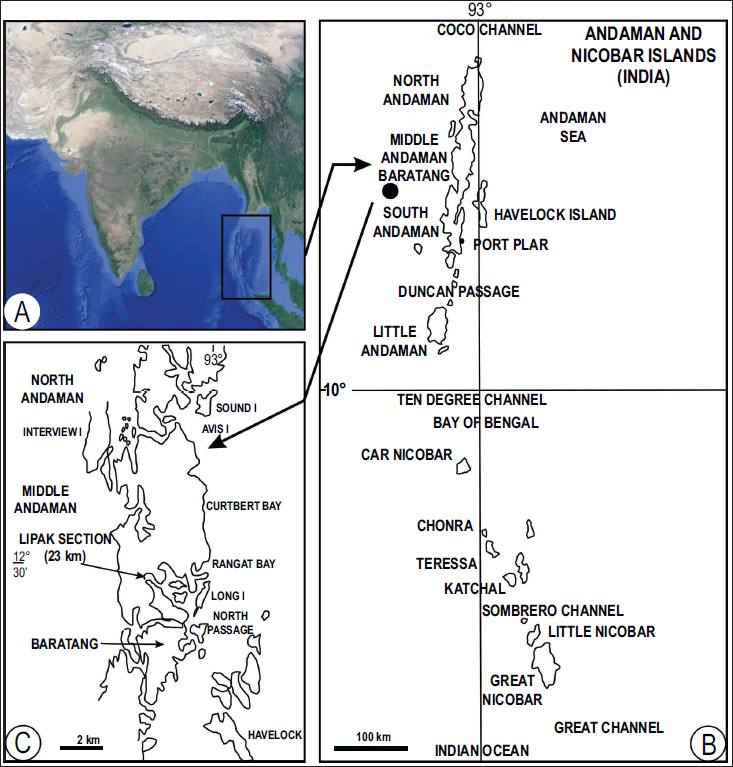
The present specimens of Paleodictyon were collected during a visit to the Andaman and Nicobar Islands in March 1989 by one of the authors (SAJ). These specimens are catalogued as AZ120AG120Ga and AZ120AGb and reposited in the Museum, Geology Department, Panjab University, Chandigarh. Paleodictyon-bearing finely laminated Lipak Black Shale and greywacke sandstone samples were collected from the right side of the road near Camp No 10, Lipak Section, where they are partially preserved as convex hyporelief on the base of greywacke sandstone (Figure 1; Plate 1.1–1.3). The Paleodictyon and associated trace fossils are preserved as convex hyporelief at the sole of the greywacke (Plate 1.4). The preserved portion indicates a regular hexagon measuring ~27 mm, with a total area of ~1900 mm²; string width 2 mm; mesh diameter 4 mm, showing a weaving/overlap pattern (Plate 1.1–1.3). The preserved specimen closely matches P. nodosum in its morphological parameters and is therefore assigned to this ichnospecies. Palynological data suggest an Early Eocene age for Paleodictyon-bearing flysch (Mandal et al., 1994). Although ichnofossils have been reported in recent years from the North and South Andaman Islands (Anju et al., 2016; Bandopadhyay et al., 2009), this constitutes the first record of Paleodictyon from the Andaman region. Typical turbidite sequences from the Andaman Islands, which preserve Paleodictyon, belong to the P. nodosum of a distal fan sequence with a bathymetry of approximately 2000 m.
(A) Map of India showing the location of the Andaman–Nicobar Islands. (B) Map of Middle Andaman and Baratang (asterisk marks study area). (C) Map of Middle Andaman showing the road connecting Kadamtala and Mayabundar. The Paleodictyon-bearing sample was collected from the right side of the road near Camp No. 10, Lipak Section, where it is partially preserved as convex hyporelief on the base of greywacke sandstone.
(A). Paleodictyon specimen fluted by currents coming from bottom left (Arrow). Extrapolation of partially preserved specimen suggests one side of regular hexagon measuring ca. 27 mm with total area of ca. 1,900 mm2 and string size of 2 mm displaying weaving or overlap pattern. Mesh diameter 4 mm, (B). Enlarged view of same specimen as in 2A displaying weaving or overlap pattern, (C). Partially preserved specimen of Paleodictyon (Arrow), (D). Exposure of finely laminated Lipak Black Shale and greywacke sandstone containing Eocene palynofossils with reworked Late Triassic and Late Cretaceous palynofossils, (E). Bottom of greywacke sandstone containing other trace fossils (F) along with Paleodictyon preserved as convex hyporelief.

Paleodictyon is poorly understood from India and is known from only a few reports. Kumar et al. (1978) and Kumar and Srivastava (1989) described poorly preserved specimens assigned to Paleodictyon carpathicum from the Late Cretaceous flysch of the Malla Johar Himalaya. However, close examination of the specimen photographs illustrated by Kumar and Srivastava (1989, Pl. II, 7) and described as Paleodictyon from the Malla Johar area of the Tethyan Himalaya reveals a mesh composed of large hexagonal structures which, based on the scale bar (see Kumar & Srivastava, 1989, Pl. II, 7), is not comparable to Paleodictyon (Meneghini, 1850). Instead, the morphology closely matches Megagrapton (Książkiewicz, 1968) and is therefore herein reassigned to this ichnogenus as Megagrapton aequale (Seilacher, 1977). Other records include P. strozzii from Palaeogene sediments south of Kohima town, Nagaland (Srivastava et al., 2021), and Paleodictyon maximum from the Laisong flysch sediments of Manipur (Khaidem et al., 2015). P. strozzii, described from the Palaeogene sediments south of Kohima town, Nagaland (Srivastava et al., 2021, p. 60, F*igure 4g–4h), exhibits a large mesh size and morphology comparable to Megagrapton (Książkiewicz, 1968), and is herein reassigned to this ichnogenus.
DISCUSSIONS
It is a matter of argument whether the ichnofossil Paleodictyon can be directly related to the present-day species P. nodosum, a form with Chinese Checkerboard-like tiny holes making a regular hexagonal pattern now known from the recent deep-sea of major oceans of the world (Durden et al., 2017; Ekdale, 1980; Gerdes et al., 2021; Míguez-Salas et al., 2023; Rona & Merrill, 1978; Rona et al., 2009). Seilacher (1977), who discovered P. nodosum from the Eocene of the Spanish Flysch, is preserved essentially as a convex hyporelief on the bottom of the sandstone. On the contrary, the recent checkerboard-like hexagonal pinhole patterns in the major oceans of the world are formed at the low-energy sediment-water interface. They should be assigned to a new ichnogenus and ichnospecies. Roma et al. (2009) and others (see Míguez-Salas et al., 2023) appear to have erred in assigning the name P. nodosum to such recent, low-energy, deep-sea forms without providing concrete evidence. Therefore, the taxon does not deserve to be called a ‘living fossil’.
Both fossil and recent Chinese-checkered hexagonal forms form under low-energy conditions. However, fossil forms preserved within the sediment were partially or entirely exposed by high-energy turbidity currents, leading to the preservation of fossil Paleodictyon on the undersurfaces of sandstones. They removed soft sediment until a cohesive level was reached. The traces at that level became cast. Traces made at the surface would not have any chance to be preserved under the onslaught of high-energy turbidity currents. The modern deep-sea hexagonal traces were produced at the sediment-water interface and exhibit behaviour entirely different from that of the fossil occurrences; hence, they should not be named after the fossil P. nodosum and should not be considered a living fossil.
Footnotes
Acknowledgements
The author would like to thank Professors Ashok Sahni and O.N. Bhargava for their encouragement. The dedication of the Team of Doctors at Narayana Hrudayalaya Hospital, Bangalore, and the family’s support for SAJ are deeply appreciated.
Declaration of Conflicting Interests
The author declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Ethical/Patient Consent (If Required) Taken or Not
Not required (NA).
Funding
The author received no financial support for the research, authorship and/or publication of this article.
Statement Regarding Plagiarism or Assisted Content from AI (If Taken)
No help taken from AI.