
Abstract
The early Eocene period marks a critical juncture in India’s geological and biological history, as the subcontinent drifted northwards as a physically isolated landmass before colliding with Asia, influencing global evolutionary patterns. The past two decades have seen significant advances in the documentation of India’s Eocene mammals and our understanding of their evolutionary and palaeobiogeographic significance. This article presents an overview of recent advances in our knowledge of early Eocene terrestrial mammals of India and their role in addressing questions of global importance, particularly in the context of India’s biogeographic relations and the early evolutionary history of perissodactyls, primates, artiodactyls, and bats. We contend that taxa with the hitherto underemphasized Asian affinities form a significant component of the Indian early Eocene mammal fauna, constraining the timing of India-Asia contact to ~55 Ma or earlier. On the other hand, the consistently favoured idea of India-Europe biotic interchange during this interval is weakly supported, since the purported affinities of several Indian early Eocene mammalian taxa exclusively with the European forms are now questionable.
INTRODUCTION
During the early Paleogene Period, the Indian subcontinent, having drifted northward from its Gondwanan origins, stood at a crossroads between physical isolation and subsequent collision with Asia. India’s fossil biota from this period represents a melting pot of ancient lineages and emerging fauna, revealing a dynamic palaeobiogeographic story preserved in sites like the Vastan lignite mine of Gujarat (see Chatterjee et al., 2017). This article deals with India’s biogeographic relationships with other landmasses during the early Eocene. It examines its role as a raft of Gondwanan relics, a cradle for new species, and a bridge for possible faunal exchanges with Asia, Europe, and Africa.
The past 25 years or so have seen an explosion of research on the Eocene mammals of India from the lignite mines in the Surat region of Gujarat, western India. Initial investigations of these sections yielded fish remains (Bajpai & Kapur, 2004; Samant & Bajpai, 2001) but sustained field work conducted at Vastan (Figure 1) over subsequent years by SB along with his students (VV Kapur, DP Das, N Saravanan, and R Sharma) resulted in a considerable amount of new data on early Eocene (~55 Ma) mammalian faunas from the lignite mine at Vastan, district Surat (Bajpai et al., 2005a, 2005b, 2005c; Bajpai et al., 2006; Bajpai et al., 2007a, 2007b; Bajpai et al., 2008, 2009; Das et al., 2022; Kapur & Bajpai, 2015; Kapur et al., 2017a; Rautela & Bajpai, 2023; see Kapur et al., 2022 for a summary). Many of these important early discoveries were reported in a series of publications in the Journal of the Palaeontological Society of India. Parallel efforts made independently by another group of workers also resulted in significant data on Eocene mammals from the same section (e.g., Rana et al., 2005; Rose et al., 2006; Rose et al., 2009a; Rose et al., 2013, 2014, 2020). Together, investigations carried out by these two groups have provided a wealth of globally significant information on the early Eocene terrestrial mammal faunas of the Indian subcontinent, addressing a significant gap in our knowledge of vertebrate evolution during a critical juncture in India’s geological history. At least 10 eutherian mammal orders are represented in the Vastan assemblage, which is found in a coastal marsh-bay complex of mangrove swamps and lagoonal muds (Prasad et al., 2013). The Vastan fauna has garnered international interest because of its clear implications for several modern land mammal orders’ early evolution, biogeographic origins, and dispersal patterns. It is now becoming increasingly clear that a number of mammalian taxa from the Indian early Eocene are the most plesiomorphic and, in some cases, stratigraphically oldest records in the Cenozoic of South Asia.

Multiple approaches have been used to constrain the age of the mammal-bearing horizon at Vastan: dinoflagellate biostratigraphy, 87Sr/86Sr isotope ratios, and carbon isotope stratigraphy (e.g., Clementz et al., 2011; Garg et al., 2008). The main mammal-yielding horizon at the Vastan mine lies 2–3 m above the basal coal seam (Seam 2) and at least 20 m stratigraphically below the Carbon Isotope Excursion (CIE) correlated to the second Eocene Thermal Maximum or ETM-2 (Clementz et al., 2011). The age of this mammal level was, until recently, considered to be at least 54.5 Ma (see Kapur et al., 2022). However, as shown by recent high-resolution investigations (Westerhold et al., 2018), the age of the ETM-2 now stands revised to 54.05 Ma from the previous estimate of 53.7 Ma. Thus, the age of the main mammal-yielding layer at Vastan is now estimated at ~55 Ma (Rautela & Bajpai, 2023).
Outside of India, a significant early Eocene terrestrial mammal-yielding horizon is the Ghazij Formation in the coal mines near the city of Quetta in Baluchistan (Pakistan), which is known to have produced a diversified assemblage of mammals (e.g., Gingerich et al., 1997, 2001). The middle Ghazij Formation, which contains the oldest known mammal-bearing interval in the Cenozoic of Pakistan, cannot be older than the P6 zone or younger than the C24R-C24N transition; that is, is approximately between 54.5 and 54 Ma in age (Missiaen & Gingerich, 2014). Therefore, the mammal-bearing interval of the Cambay Shale at Vastan mine, which is ~55 Ma (early Ypresian) in age, appears to predate the middle Ghazij mammal-yielding interval of Baluchistan, and is chronologically equivalent (in part) to the marine lower Ghazij Formation. The suggested correlation (Missiaen & Gingerich, 2012; Missiaen et al., 2011) of the middle Ghazij Formation with the Wasatchian North American Land Mammal Age (NALMA) and that of the upper Ghazij with the late Wasatchian to early Bridgerian NALMA, or the middle to late Bumbanian Asian Land Mammal Age (ALMA), is consistent with the estimated age of ~55 Ma for the main mammal-yielding horizon (i.e., the one slightly above Seam 2) at Vastan.
Additionally, the Vastan mammal faunas assume significance because they broadly coincide in age with the withdrawal of the Neotethys and the initiation of India–Asia collision (see Clementz et al., 2011), as well as with the intense warming intervals at the Paleocene-Eocene boundary or P/E boundary (i.e., Paleocene-Eocene Thermal Maximum or PETM), at 56 Ma, and the ETM-2, at 54.05 Ma. The Asian affinities of some of the mammalian taxa from Vastan, as discussed below, are important in constraining the timing of the initial India–Asia collision. This review focuses on the biogeographic affinities of early Eocene mammal faunas of India, particularly about their purported European and/or Asian connections (Table 1).
Updated list of early Eocene (~55 Ma) mammal faunas of Cambay Shale, India, and their biogeographic affinities.
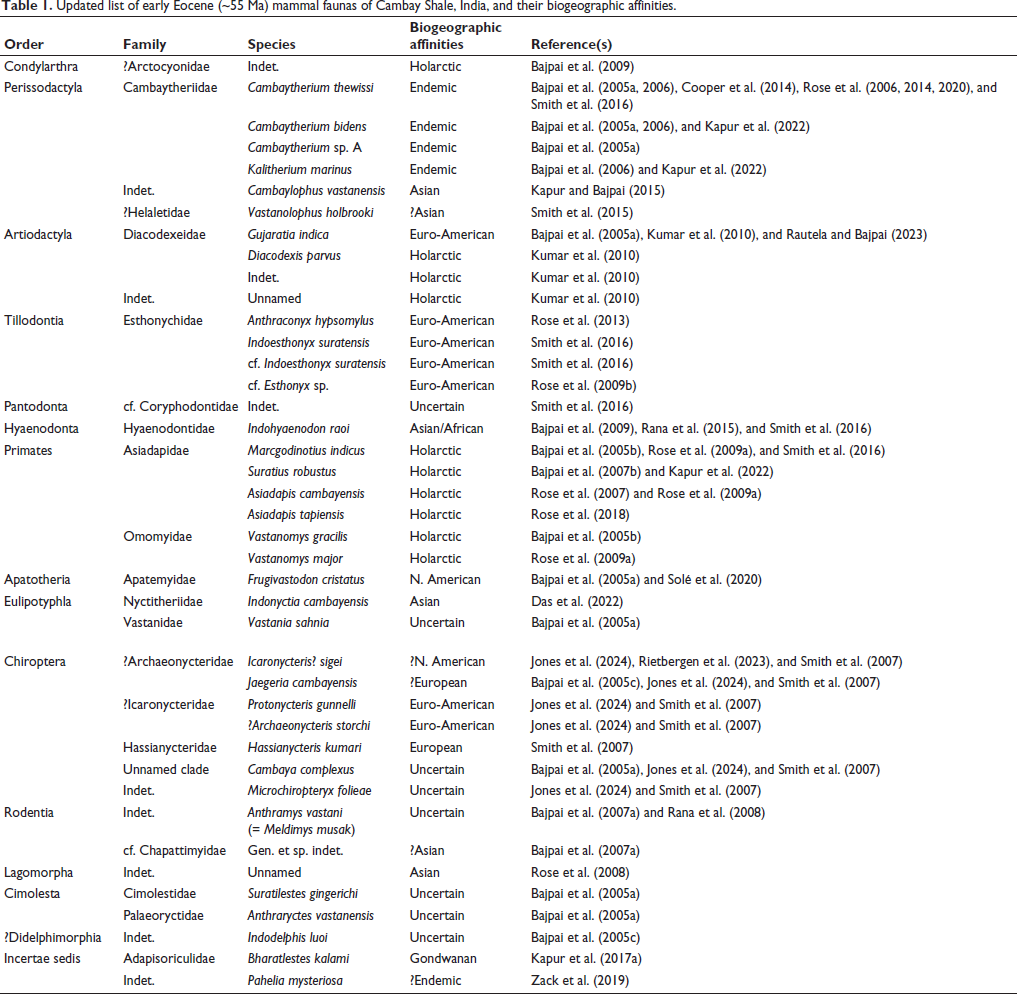
DISCUSSION
The issue of European connection
The supposed European ties during the early Eocene are primarily based on four main mammal taxa from the Cambay Shale: adapoid primates, bats, artiodactyls, and ailuravine rodents. Here we address this issue by re-evaluating the affinities of each of these groups.
Primates
Two early-diverging clades of euprimates, the lemur-like Adapoidea and the tarsier- or galago-like Omomyidae, have been described from the early Eocene Cambay Shale (~55 Ma) of Vastan lignite mine. While contemporary adapoids and omomyoids are well documented from North America and Europe, there are few records of plesiomorphic euprimates from Africa and Asia. The Cambay Shale primates from Vastan are represented by adapoids (Marcgodinotius indicus Bajpai et al., 2005b; Suratius robustus Bajpai et al., 2007b; Asiadapis cambayensis Rose et al., 2007) and omomyoids (Vastanomys gracilis Bajpai et al., 2005b; V. major Rose et al., 2009a). Among the adapoids, Marcgodinotius indicus is the most common and plesiomorphic primate at Vastan. In contrast, Suratius robustus, described initially as an omomyid, is now considered a relatively derived adapoid based on the absence of a p1. The Indian adapoids from Vastan were initially referred to the primitive European family Notharctidae, and this was interpreted as evidence of India-Europe biogeographic connection during the early Eocene (Rose et al., 2009a). However, the Vastan adapoids were later classified in their own family, Asiadapidae, with Asiadapis as its type genus (Fleagle, 2013; Rose et al., 2009a).
The sole omomyoid known from the early Eocene of India is Vastanomys (type species V. gracilis), which may also be an early-diverging genus of the omomyoid radiation (Bajpai et al., 2008).
In addition to adapoids and omomyoids, a purported eosimiid (Anthrasimias gujaratensis) was also described from the early Eocene Vastan section (Bajpai et al., 2008). About the size of a mouse (~75 g), it consumed a mixed diet of fruits and insects. However, A. gujaratensis was later considered to be an adapoid, congeneric with Marcgodinotius (e.g., Rose et al., 2009a). Additional specimens are required to definitively confirm or deny the existence of eosimiids at Vastan. In any case, the co-occurrence of Adapoidea, Omomyoidea, and ?Anthropoidea in the Vastan fauna suggests that the Indian early Eocene primates were fairly diversified.
Chiropterans
Bats form the most diverse component of India’s early Eocene Vastan mammal faunas. Bats from Vastan were first described by Rana et al. (2005) based on two upper molars, referred to two genera, namely cf. Icaronycteris and cf. Archaeonycteris. Subsequently, Smith et al. (2007) reported seven genera of bats from the same locality – Icaronycteris, Protonycteris, Archaeonycteris?, Hassianycteris, Cambaya, Microchiropteryx, and Jaegeria, all based on lower jaws and/or teeth. These were included in four families: Icaronycteridae, Archaeonycteridae, Hassianycteridae, and Palaeochiropterygidae.
Smith et al. (2007) claimed that the bat fauna from the Indian Eocene strongly resembles that from the European Eocene. In particular, three groups of Vastan bats have been cited as evidence of Europe-India biotic interchanges: Icaronycteridae, Hassianycteridae, and Archaeonycteridae (Smith et al., 2016). However, Smith et al. (2012) stated that the genus Icaronycteris might not be monophyletic (Smith et al., 2012, p. 33). Recent investigations by Rietbergen et al. (2023) showed the genus Icaronycteris to be closely related to the North American Onychonycteris, noting that Icaronycteris sigei from Vastan is not a member of the genus Icaronycteris or the family Icaronycteridae. According to these authors, the definitive geographic distribution of Icaronycteris and Icaronycteridae is restricted to North America. Another recently published, comprehensive phylogenetic analysis (Jones et al., 2024) places Icaronycteris sigei closely with the North American Palaeochiropteryx sambuceus instead.
Archaeonycterids, a family previously limited to Europe (Smith et al., 2012), were identified in Vastan by Smith et al. (2007), who named two new species: Protonycteris gunnelli and Archaeonycteris? storchi. According to a phylogenetic analysis by Jones et al. (2024), the two Vastan species form a group with Archaeonycteris brailloni, three species of Icaronycteris, and Xylonycteris stenodon, with this group being closest to onychonycterids, which also include the oldest known bat, ?Archaeonycteris praecursor, from the earliest Eocene of Europe (Tabuce et al., 2009). This entire group includes most of the early bats from Europe and North America, raising the possibility of early Eocene faunal exchanges between North America, Europe, and India.
Smith et al. (2007) also described a purported hassianycterid from Vastan (Hassianycteris kumari), which remains poorly known. Another bat, initially described as a nyctitheriid insectivore (Bajpai et al., 2005a), is Cambaya complexus, which was later referred to Hassiancyteridae by Smith et al. (2007, 2012). However, Jones et al. (2024) recently showed Cambaya as the oldest early-diverging bat, not a hassianycterid, but closer to Asian Protorhinolophus and Afro-European necromantids.
Smith et al. (2007) described Microchiropteryx as a palaeochiropterygid, a family known from Europe, and possibly Asia and North America (Jones et al., 2019; Smith et al., 2012). Jones et al. (2019) described another palaeochiropterygid from Turkey and suggested the dispersal of volant mammals between Europe and India via the Pontide terrane, northern Turkey. However, the idea of such early Eocene exchanges between Europe and India in the case of non-volant terrestrial mammals was not considered tenable by Métais et al. (2018). Interestingly, a recent comprehensive phylogenetic study by Jones et al. (2024) showed that Microchiropteryx is the oldest crown bat, which is not closely related to stem bat families, including Palaeochiropterygidae. In fact, according to Jones et al.’s (2024) analysis, palaeochiropterygids form a sister taxon with hassianycterids and the Asian palaeochiropterygid Lapichiropteryx, which suggests a faunal dispersal between Asia and Europe during the middle Eocene.
Jaegeria cambayensis, first described as a herpetotheriine marsupial (Bajpai et al., 2005c), was later identified as a bat (Smith et al., 2007). However, its relationships within Chiroptera are still poorly understood. Smith et al. (2007) suggested that Jaegeria is distinct from Icaronycteridae, Archaeonycteridae, and Hassianycteridae. More recently, Jones et al. (2024) showed it to be a sister taxon to Archaeonycteris pollex and A. trigonodon from the middle Eocene of Messel, Germany. This group is the closest to the clade including Icaronycteris sigei from Vastan and Palaeochiropteryx sambuceus from the early middle of the Eocene of North America, suggesting dispersal from India to Europe/North America during the late early Eocene.
Overall, the Vastan bats represent a diversified assemblage of mixed affinities, primarily Holarctic (European and North American), and are of considerable importance in understanding the origins, diversification, and dispersal of stem and, probably, crown bat lineages. However, since the presence of Icaronycteridae in the Vastan fauna is now discarded, and the presence of the families Archaeonycteridae, Hassianycteridae, and Palaeochiropterygidae is brought into question, major revisions are required at the suprageneric level. Nevertheless, a close relationship between some of the Vastan bats and early Eocene European and North American bats is consistently supported. However, this relationship may reflect the paucity of early Eocene bats in Central Asia, as pointed out by Smith et al. (2007). Relationship to Asian bats is open to discussion and must await the discovery of more complete forms from the early Eocene of Asia. If the origin of bats is Asian (Yu et al., 2014), as is also evident from the discovery of Altaynycteris from the early Eocene of China (Jones et al., 2021), then the relationships between Indian and Asian bats must be evaluated, similar to other small insectivorous mammals, such as nyctitheriids (Das et al., 2022). The allocation of upper dentition and other fossil remains to the Vastan bats may prove useful in deducing their relationships accurately.
In any case, bats may not be the most suitable taxa for inferring land connections based on palaeobiogeographic relationships, since chiropterans are known to have developed powered flight early on in their evolution (Simmons et al., 2008). Furthermore, chiropterans had a cosmopolitan distribution by the early Eocene, being capable of traveling considerable distances over water (Beck, 2017; Smith et al., 2012). Interestingly, Beck (2017) also noted that Australonycteris, one of the globally oldest bats (Hand et al., 1994), may have been fully volant, which, if true, pushes back the origin of flight in chiropterans by nearly two million years, to 54.6 Ma.
Artiodactyls
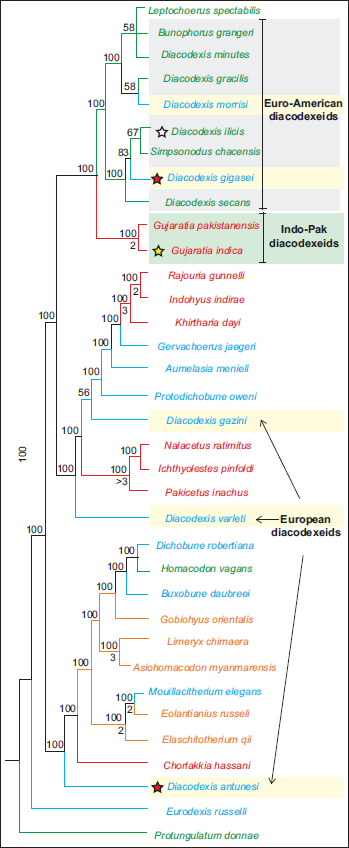
The earliest artiodactyl from the Indian subcontinent is Gujaratia indica from the early Eocene Cambay Shale of Vastan (Bajpai et al., 2005a; Rautela & Bajpai, 2023). Gujaratia is currently known from two species, G. indica and G. pakistanensis, the latter being the type species of this genus known from the Eocene of Pakistan (Thewissen et al., 1983). Some workers considered Gujaratia to be congeneric with the Euro-American genus Diacodexis (Boivin et al., 2018; Kumar et al., 2010). According to earlier interpretations, Gujaratia has European connections and is closely related to the European Diacodexis, particularly D. antunesi. However, the recently described new dental material and phylogenetic results do not support this interpretation (Rautela & Bajpai, 2023). This latter study shows that Gujaratia (including G. pakistanensis and G. indica) is monophyletic. In contrast, both Diacodexis and diacodexeids are strongly polyphyletic (Figure 2). Gujaratia appears to be related to diacodexeids from North America and D. gigasei and D. morrisi from Europe, which form a paraphyletic group. The European Diacodexis represents an unnatural grouping. Consequently, the affinities of Gujaratia are unclear and cannot be used to deduce biogeographic connections. Interestingly, G. pakistanensis shows more plesiomorphic traits than Diacodexis secans (Thewissen & Hussain, 1990). It is also to be noted that the early Eocene artiodactyls from the Indian subcontinent are distinct from coeval Asian forms. According to some workers (Bai et al., 2023; Orliac & Ducrocq, 2012), faunal exchanges between India and Asia became possible during the early middle Eocene, as suggested by the shared taxa (e.g., Haqueina and Dulcidon) in the Mongolian Plateau, southeastern China, and the Indian subcontinent.
50% majority-rule consensus tree showing phylogenetic relationships of early artiodactyls (modified after Rautela & Bajpai, 2023). Taxa are mentioned in colours indicating their place of origin: red, Indian subcontinent; orange, Asia; blue, Europe; green, North America. Colored stars before taxa indicate the earliest records of artiodactyls: yellow, India; red, Europe; white, North America. Note that Gujaratia is closer to the Euro-American diacodexeids but not to the European Diacodexis antunesi.
Rodents
The presence of isolated rodent teeth from Vastan, referred to the ailuravine genus Meldimys, has often been cited as evidence of European affinities and intercontinental migrations between Europe and India during the early Eocene (Rana et al., 2008). Kapur et al. (2022) doubted the purported record of ailuravine rodents in Vastan based on the fact that this European group is entirely absent in the nearly contemporary or younger rodent assemblages from the Subathu Formation of NW Himalaya, India, and from the Mami Khel Clay of Barbara Banda (Kohat), and Kuldana Formation, Ganda Kas (Punjab), both in Pakistan, despite extensive sampling of these deposits (De Bruijn et al., 1982; Gupta & Kumar, 2015; Hussain et al., 1978; Kumar, et al., 1997a, b). A more recent work (Vianey-Liaud et al., 2021) has shown that the material from Vastan clearly does not pertain to the genus Meldimys. The Vastan rodent fauna also includes a possible chapattimyid that is still unnamed (Bajpai et al., 2007a).
The issue of Asian connection
The Asian stamp of India’s early Eocene mammal faunas is documented mainly by tapiroids (Perissodactyla), lagomorphs, and eulipotyphlans.
Tapiroids (Perissodactyla)
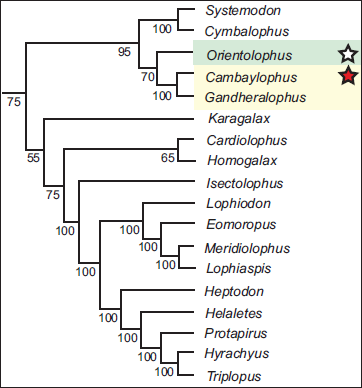
Kapur and Bajpai (2015) named the first early Eocene tapiromorph, Cambaylophus, from the Cambay Shale of Vastan lignite mine. Phylogenetically, Cambaylophus, along with Gandheralophus from the Ghazij Formation (Missiaen & Gingerich, 2012), was found to be closest to Orientolophus from the earliest Eocene (Bumbanian ALMA) Lingcha Formation of China (Figure 3). Another tapiroid from Vastan is Vastanolophus, a ?helaletid (Smith et al., 2015). Both these taxa at Vastan suggest direct faunal connections with Asia.
50% majority-rule consensus tree showing phylogenetic relationships of early tapiromorph perissodactyls (modified after Kapur & Bajpai, 2015). The Vastan tapiroid Cambaylophus (indicated by a red star), along with Gandheralophus from Pakistan, shows close (sister-taxon) relationships with the Chinese Orientolophus (indicated by a white star) from the early Eocene (early Bumbanian ALMA).
Lagomorphs
Rose et al. (2008) described a few ankle bones of an unnamed lagomorph (rabbits, hares, and pikas) from the Vastan mine. They noted their strong similarities to those of modern lagomorphs and the tarsal bones known from the Eocene of Asia. Recently, Lopatin and Averianov (2021) described additional dental remains of the oldest known lagomorph (sensu lato) from the base of the Bumban Member of Naran Bulak Formation (earliest Eocene) of Mongolia and created a new clade, Eulagomorpha, uniting the Eocene stem lagomorphs and the crown group lagomorphs. According to these authors, this new clade appeared in Asia shortly before the P/E boundary. Thus, the Vastan lagomorphs offer compelling evidence for faunal exchanges and, consequently, the collision between India and Asia, at or slightly before the P/E boundary. The Vastan lagomorphs are most likely Asian immigrants, considering the existence of Paleocene mimotonids and Eocene eulagomorph Arnebolagus in Asia (Lopatin & Averianov, 2021).
Eulipotyphlans
Indonyctia represents the sole nyctitheriid from the Indian subcontinent and shows close similarities to Voltaia from the late Palaeocene (Gashatan ALMA) of Kazakhstan, Central Asia (Das et al., 2022). The presence of a nyctitheriid in the early Eocene of India suggests a wider palaeobiogeographic distribution and complex dispersal of this Holarctic family. Das et al. (2022) suggested that a nyctitheriid related to Voltaia may have migrated directly from Asia to India, possibly before the P/E boundary, and then evolved in isolation in India.
The issue of endemic evolution on the drifting Indian plate
The early Eocene fossil record from India provides compelling evidence in support of an Indian origin (Out-of-India hypothesis) for some of the major terrestrial groups, particularly Perissodactyla, whose antiquity can now be traced to the iconic early Eocene taxon Cambaytherium Bajpai et al., 2005a. Cambaytherium was identified as a perissodactyl from the Cambay Shale of Vastan mine two decades ago and, together with another genus Kalitherium, was classified in its own new family, Cambaytheriidae. Cambaytherium is the most abundant and amongst the largest mammals known from the Cambay Shale (Bajpai et al., 2005a, 2006; Rose et al., 2020). Cambaytheriids are closely related to anthracobunids from the middle Eocene Indo-Pak localities, and together these stem perissodactyl groups are sister taxa to crown Perissodactyla (Cooper et al., 2014; Rose et al., 2020). Both cambaytheriids and anthracobunids are endemic to the Indian subcontinent and provide strong evidence that the perissodactyls originated in the Indian subcontinent.
Other endemic taxa that evolved in India include the monospecific erinaceomorph family Vastanidae (type species Vastania sahnia Bajpai et al., 2005a), diagnosed by a high protoconid on p4 with a straight and lingual cristid obliqua and lacking a talonid basin. Ongoing work is likely to reveal its relationships more clearly.
Gondwanan relics
Kapur et al. (2017a, b) described an early Eocene adapisoriculid Bharatlestes kalami from the Cambay Shale of Vastan, based on well-preserved lower dentition. This new adapisoriculid represents a family previously unknown from the Eocene of the Indian subcontinent. Phylogenetic analysis (Kapur et al., 2017a) suggested that Bharatlestes is derived relative to the well-known Late Cretaceous Deccan intertrappean genus Deccanolestes (Prasad et al., 2021) and the Afro-European Paleocene genus Afrodon. Bharatlestes provides strong evidence for the continued survival of a Gondwanan mammal lineage in the Indian subcontinent after the Deccan volcanic activity (Cretaceous–Paleogene transition).
Other mammalian taxa
An Eocene apatemyid (Frugivastodon cristatus) was identified for the first time from the Indian subcontinent from the Vastan Cambay Shale (Bajpai et al., 2005a). Apatotheria is a group known from North America, Europe, and recently, Asia (Lopatin & Averianov, 2022). According to Solé et al. (2020), the Indian apatemyid has affinities to the genus Aethomylos of North America, with which it may form a clade. The dispersal pattern of apatemyids is poorly understood.
A carnivorous mammal in the early Eocene Vastan fauna is the hyaenodontid Indohyaenodon raoi (Bajpai et al., 2009; Rana et al., 2015). Hyaenodontids are commonly recorded from Africa and the Holarctic continents, but their first appearance dates to the middle Paleocene of Africa. I. raoi exhibits a combination of primitive and derived characters and is closely allied to other South Asian hyaenodontids. The biogeographic affinities of I. raoi are yet to be conclusively established, but the case of an Asian origin cannot be ruled out (Kapur et al., 2022).
Bajpai et al. (2009) described an unnamed large, bunodont, tritubercular upper molar of a condylarth, reminiscent of the family Arctocyonidae, from the early Eocene Cambay Shale of Vastan. This enigmatic taxon may represent a transitional form between the North American triisodontids of Paleocene age and the Holarctic mesonychians (Tabuce et al., 2011).
Three or four tillodonts are known from Cambay Shale, including Anthraconyx and Indoesthonyx with affinities to Euro-American esthonychids (Rose et al., 2013; Smith et al., 2016).
Zack et al. (2019) described an indeterminate ungulate-like herbivorous mammal, Pahelia mysteriosa, from the early Eocene Cambay Shale of Tadkeshwar. Its affinities are still uncertain, but according to the authors, Pahelia may be endemic to India, like the Ghazij quettacyonids of Pakistan, similar to cambaytheres.
Other eutherian mammals of uncertain affinities include the cimolestan Suratilestes, palaeoryctid Anthraryctes, and an unnamed pantodont (Bajpai et al., 2005a; Smith et al., 2016).
Metatherians are extremely rare in the Cenozoic of the Indian subcontinent and Asia and, compared to those from other continents, do not contribute significantly to the global fossil record. Nevertheless, a single tooth of a possible metatherian (Indodelphis) is currently known from the early Eocene of Vastan (Bajpai et al., 2005c). Additional material is needed for a definitive assignment. Ongoing work will likely shed more light on the presence of metatherians at Vastan.
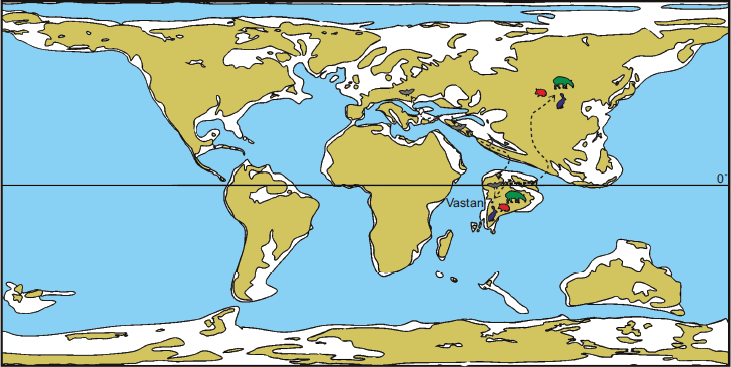
India–Asia collision
The timing of the India–Asia collision remains a hotly debated topic and is a dynamic area of research with multiple lines of evidence (Hu et al., 2016). Uncertainties continue to persist, but most studies converge on the initiation of collision around 55–60 Ma, based on evidence including the cessation of marine sedimentation, sediment provenance changes, palaeomagnetic data, and high-pressure Himalayan metamorphic rocks (e.g., Zhang et al., 2024; Zhou et al., 2024). As stated above, the age of the Vastan mammals is ~55 Ma, slightly younger than the P/E boundary. Some of the land mammals from Vastan, such as tapiroid perissodactyls, lagomorphs, nyctitheriids, and possibly even hyaenodontids, suggest Asian affinities (Figure 4). A subaerial contact or proximity between two landmasses is required for terrestrial faunal exchanges. Therefore, the close similarity of the mammalian taxa from India with those from Asia suggests that the collision between these landmasses began as early as the P/E boundary (56 Ma) or perhaps even earlier (Clementz et al., 2011). The discovery of late Paleocene mammals in India will allow a rigorous test of this hypothesis, but the age and close relationships of some of the Vastan taxa, particularly the tapiroid Cambaylophus with the Chinese Orientolophus (Bai et al., 2018; Kapur & Bajpai, 2015), suggests that a direct land connection between India and Asia via intervening crustal blocks (including the Kohistan-Ladakh Island Arc) may have been in place at 55 Ma or earlier. The medium size of these mammals makes such terrestrial connections especially likely (see Prasad & Sahni, 1999).
Recent fossil and molecular evidence from a few other faunal and floral groups, in addition to mammals, points to biotic exchanges between India and Asia as early as late Paleocene or even Late Cretaceous. These include fossil palms (Sabalites) from the late Paleocene of Tibet (Gao et al., 2025) and the nearly cosmopolitan natatanuran frogs (Yuan et al., 2019). Interestingly, the evidence from fossil palms (Gao et al., 2025) lends credence to the idea that biotic exchanges between India to Asia were made possible by the Kohistan-Ladakh Island Arc, as discussed in Chatterjee et al. (2017).
CONCLUSIONS
The oldest terrestrial mammal fauna from the Cenozoic of the Indian subcontinent is known from the lignite mines near Surat, particularly Vastan, in the Gujarat state of western India. The mammal-yielding horizon is ~55 Ma in age and has yielded a diverse fauna representing at least 10 eutherian orders. The assemblage is largely Holarctic (e.g., primates, artiodactyls, nyctitheriids, and tillodonts) but also includes certain endemic groups (e.g., cambaytheriid perissodactyls, vastanid eulipotyphlans) and Gondwanan relics (e.g., adapisoriculids). Whereas previous interpretations focused on European affinities of the Vastan mammal fauna, it is emphasised here that this fauna also has a distinct component with Asian affinities, which needs to be studied in more detail because of its direct implications for the timing of India–Asia collision at, or before, the P/E boundary.
Footnotes
Acknowledgements
SB acknowledges with gratitude the constant encouragement he received from the Late Prof A. K. Jauhri, former editor of the Journal of the Palaeontological Society of India, for publishing many of the early discoveries of Vastan mammal faunas in this long-standing journal. SB also thanks the present editor, Prof Mukund Sharma, for inviting him to contribute an article for this special volume of the journal marking the platinum jubilee of the Palaeontological Society of India. We thank the two anonymous reviewers for their suggestions which helped improve the manuscript.
Author Contributions
SB conceptualized the article and wrote it along with AR.
Consent to participate
Not required.
Consent for publication
Not applicable.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Ethical considerations
Ethical approval is not required.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: SB acknowledges funding from the Department of Science & Technology, Government of India. AR acknowledges financial support under the Shyama Prasad Mukherjee Fellowship, CSIR-HRDG, New Delhi, Government of India (SPM09/0143(16119)/2022-EMR-I).