
Abstract
This study investigates the extinction dynamics of Globigerinoides obliquus and Globigerinoides extremus across multiple deep-sea cores from the Indo-Pacific region. By integrating biostratigraphic records, we document a pronounced diachrony in the last occurrence of G. obliquus, which persisted beyond the mid-Quaternary and survived into the Holocene at several sites. In contrast, the extinction of G. extremus is relatively synchronous across sites. The diachronous extinction pattern of G. obliquus is interpreted to be influenced by local palaeoceanographic factors, notably oligotrophic conditions within the upper-ocean mixed layer. In contrast, the synchronous disappearance of G. extremus likely reflects competitive exclusion and large-scale oceanic reorganisation starting in the Late Pliocene. Shifts in dominance within Ecogroup-1 planktic foraminiferal taxa coinciding with the onset of Northern Hemisphere Glaciation highlight significant changes in mixed-layer hydrography and species interactions. The observed faunal turnover supports the notion that environmental variables and interspecific competition collectively govern species distributions in overlapping niches. These findings challenge the reliability of G. obliquus last occurrence as a universal stratigraphic marker and underscore the need to consider regional oceanographic controls in palaeoceanographic reconstructions.
INTRODUCTION
Biostratigraphic analyses of deep-sea cores from multiple ocean basins indicate that most planktic foraminiferal events, including first (FA) and last appearances (LA), are diachronous across broad geographic regions (Berggren, Hilgen, et al., 1995; Berggren, Kent, et al., 1995; Dowsett, 1988, 1989a, 1989b; Lam et al., 2022; Sinha & Singh, 2008; Srinivasan & Sinha, 1991, 1992). Although some events appear synchronous within limited regions, others display diachronism even between cores separated by only a few kilometres (Singh & Sinha, 2022). The causes of this diachronism have been extensively investigated (Singh et al., 2021; Spencer-Cervato et al., 1994), yet many planktic foraminiferal events remain the subject of debate. In this study, we examine the extinction horizon of Globigerinoides obliquus Bolli across a suite of deep-sea cores recovered from different water masses from the Indo-Pacific region.
As unicellular, microscopic protists, planktic foraminiferal species are widely used to establish biostratigraphic zonation and correlation of deep-sea cores from the world’s oceans. However, the diachronism observed in many planktic foraminiferal events has made zonal correlation difficult, and the concept of datum planes (Hornibrook, 1966) has gained importance instead. Still, there are quite a number of planktic foraminiferal events that have been used as chronostratigraphic boundary markers; for example, Globorotalia tumida (last occurrence) is often used to demarcate the Miocene/Pliocene boundary in deep-sea cores across tropical latitudes (Singh & Sinha, 2022). Along with this, many of the planktic foraminiferal datums are widely used for biostratigraphic zonation of deep-sea sediment cores (Berggren, Hilgen, et al., 1995; Berggren, Kent, et al., 1995; Kaushik et al., 2020; Singh & Sinha, 2022; Sinha & Singh, 2008; Singh et al., 2021; Wade et al., 2011). Advances in the study of deep-sea cores from the world’s oceans have led to revisions in the stratigraphic ranges of several planktic foraminiferal species. In this article, we examine the revisions and diachronism associated with the last occurrence of G. obliquus Bolli, which, until recently, served as an important biostratigraphic marker. Bolli (1957) originally described Globigerinoides obliqua (basionym), later corrected to G. obliquus, from the Miocene of Trinidad. Subsequently, Bolli and Bermúdez (1965) established G. obliquus extremus n. subsp. (basionym) from the Miocene of Venezuela, and Perconig (1968) described G. obliquus amplus n. subsp. from the Miocene and Pliocene of Spain. Following Kennett and Srinivasan (1983), we employ the names G. obliquus and Globigerinoides extremus throughout this study. Both taxa have been reported by several authors not only from the Miocene, but also from the Pliocene and even the Pleistocene (Akers, 1972; Blow, 1979; Jenkins & Orr, 1972).
The genus Globigerinoides
Cushman (1927) established the genus Globigerinoides for planktonic foraminiferal species characterised by multiple and supplementary apertures on the spiral side, designating Globigerina rubra (basionym) d’Orbigny as the type species. In his original description of Globigerinoides, Cushman observed the presence of spine bases on the test surface. Based on surface ultrastructure, such forms were later placed in Group A by Kennett and Srinivasan (1983) and considered to have originated from Globigerina (Globigerina). However, Cushman’s designation of Globigerina rubra (d’Orbigny, 1839) as the type species conflicted with this view, since Kennett and Srinivasan (1983) subsequently assigned it to Globigerinoides Group B, characterised by a cancellate surface. Notably, in his original description of Globigerina rubra, d’Orbigny (1839) did not comment on the nature of the test surface.
The genus Globigerinoides is distinguished from Globigerina by having more than one aperture. In planktonic foraminifera, surface ultrastructure is widely regarded as a conservative morphological trait, exhibiting limited change over evolutionary time and thus providing a dependable basis for taxonomy and phylogenetic interpretation. Loeblich and Tappan (1988) highlighted that wall textures and ultrastructural elements—such as spines, spine bases, and cancellate patterns—exhibit far greater stability than more plastic features like chamber arrangement or aperture position. This view is reinforced by both earlier and later studies that employed ultrastructural characteristics to differentiate lineages and establish higher-level classifications (e.g., Bé, 1965; Hemleben et al., 1987; Kennett & Srinivasan, 1983). Because these features are less influenced by ecological variation or developmental flexibility, they are thought to capture deeper evolutionary signals. Kennett and Srinivasan (1983) divided Globigerinoides into two broad groups based on surface ultrastructure as a conservative morphological feature and their inferred origins from different Globigerina lineages. Group A includes species derived from Globigerina (Globigerina)—such as G. primordius, G. altiapertura, G. obliquus, G. extremus, G. conglobatus, and G. bulloideus, which retain spines and spine bases. Group B comprises species derived from Globigerina (Zeaglobigerina = Globoturborotalita), including G. trilobus, G. sicanus, G. immaturus, G. quadrilobatus, G. sacculifer, G. fistulosus, G. subquadratus, G. diminutus, G. mitra, G. ruber, G. seigliei, and G. tenellus, which possess a cancellate surface with only weak evidence of former spinosity. Takayanagi and Saito (1962) classified species of Globigerinoides into two groups based on the position of the primary aperture. In Group 1, the aperture lies above the two sutures connecting the three earlier chambers and includes species such as G. bolli, G. conglobatus, G. immaturus, G. sacculifer, and G. trilobus. In Group 2, the aperture is positioned above the suture between the penultimate and antepenultimate chambers, comprising species such as G. elongatus, G. obliquus, G. ruber, and G. ruber cyclostomus. However, this was fully based on a morphological character without any phylogenetic or cladogenetic consideration. Spezzaferri et al. (2015) showed with evidence from molecular phylogenies that the trilobus group is not the sister taxon of the ruber group, though they retained the ruber group aligned to Globoturborotalita type ultrastructure. Keller (1981) recognised three evolutionary lineages leading to a Globigerinoides-like morphology: one derived from Globigerina praebulloides, another from Globigerina woodi, and a third from Globigerina connecta, with the latter two species now placed in the genus Globoturborotalita. Spezzaferri (1994) likewise identified three groups of Globigerinoides: the first arising from Globigerina with a bulloides-type wall texture (as defined by Hemleben et al., 1989), and the second showing a cancellate, honeycomb-like wall texture, further divided into two subgroups with ruber- and sacculifer-type wall textures (Hemleben et al., 1989). Together, these studies consistently highlight two distinct surface ultrastructures as the basis for separating the two main groups of Globigerinoides. The fossil record shows that the earliest Globigerinoides in the late Oligocene gave rise to the trilobus group, while the ruber group emerged separately at the base of the Miocene with forms distinct from the trilobus lineage. Keeping this in view, Spezzaferri et al. (2015) argued that the trilobus group is assigned to a separate genus and erected a new genus Trilobatus for the trilobus group (type species Globigerina triloba Reuss), giving logic that two separate evolutionary lineages are recognised: one comprising Globigerinoides trilobus and its descendants, while the other formed by the G. ruber clade with its extant members and their ancestors. Globigerinoides has important stratigraphic significance. Bolli and Saunders (1985) proposed using the sudden first appearance of G. trilobus, G. immaturus, and G. sacculifer to mark the Oligocene–Miocene boundary. The different levels at which Globigerinoides first appears may be related to the rise in ocean temperatures that occurred after the cooler conditions in the Oligocene (Jenkins, 1971). This fact seems to be quite acceptable, as the members of the Globigerinoides group still prefer warm ocean waters. The number of secondary apertures differs from species to species. The number and shape of the chambers in the final whorl also show a wide range of variation.
Globigerinoides obliquus – Globigerinoides extremus
Here, we discuss the taxonomic features of G. obliquus and G. extremus documented from various deep-sea cores from the Indo-Pacific region. In addition, we examined the extinction events of G. obliquus and G. extremus. The variation in the relative abundance of both species over the last 6 million years has also been studied.
Globigerinoides obliquus (Figures 1–3); Surface ultrastructure of Globigerinoides obliquus (Figures 4 and 5). Globigerinoides extremus (Figures 6–8); Surface ultrastructure of Globigerinoides extremus (Figures 9 and 10). Globigerinoides ruber (Figures 11–13); Surface ultrastructure of Globigerinoides ruber (Figure 14).


In this study, G. obliquus is recognised by its obliquely compressed final chambers and a high-arched primary aperture (Plate 1, Figures 1–5). It differs from G. ruber in these features and from G. extremus by having a less compressed last chamber. According to Kennett and Srinivasan (1983), G. obliquus originated from G. altiapertura in Zone N5 (Early Miocene) through the development of later chambers with oblique lateral compression.
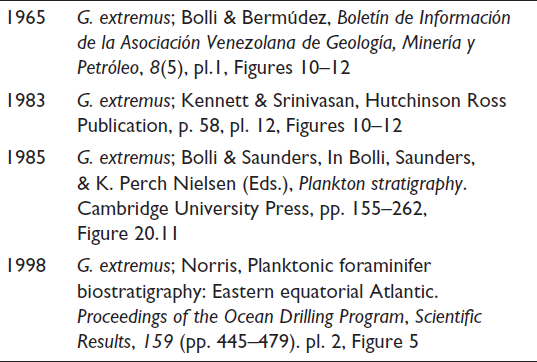
Phylogenetic considerations: G. extremus evolved from G. obliquus through progressive chamber compression in the final whorl and the development of a distinctly flattened terminal chamber (Kennett & Srinivasan, 1983).

RESULTS AND DISCUSSION
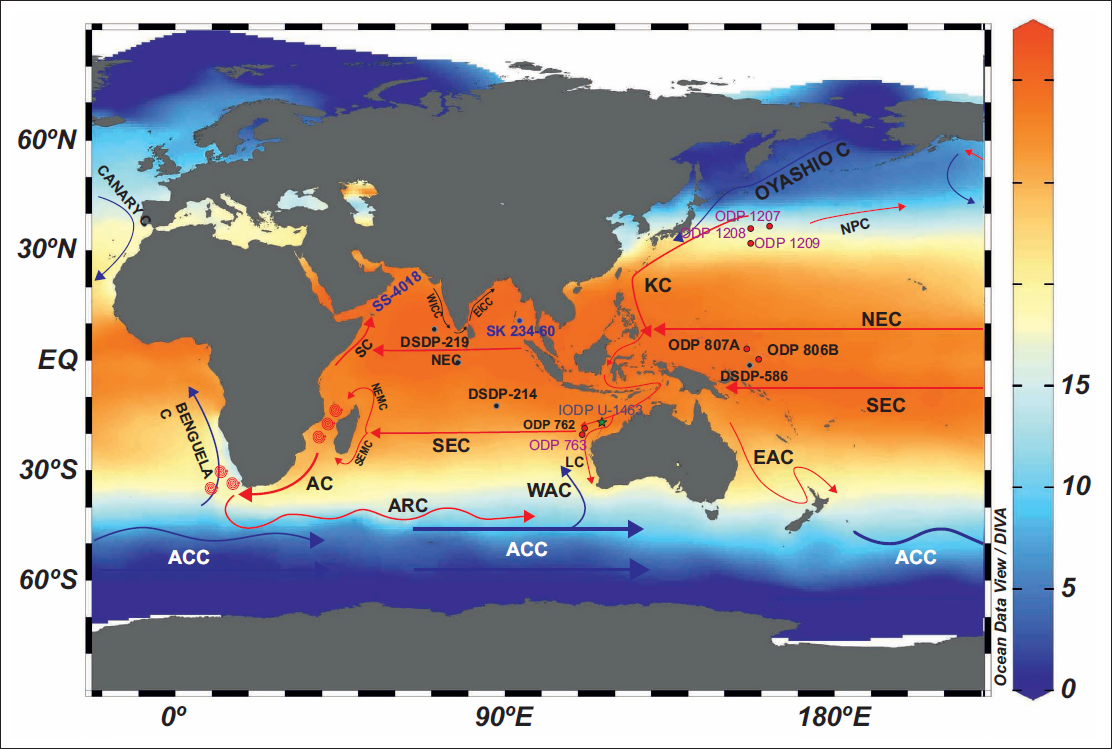
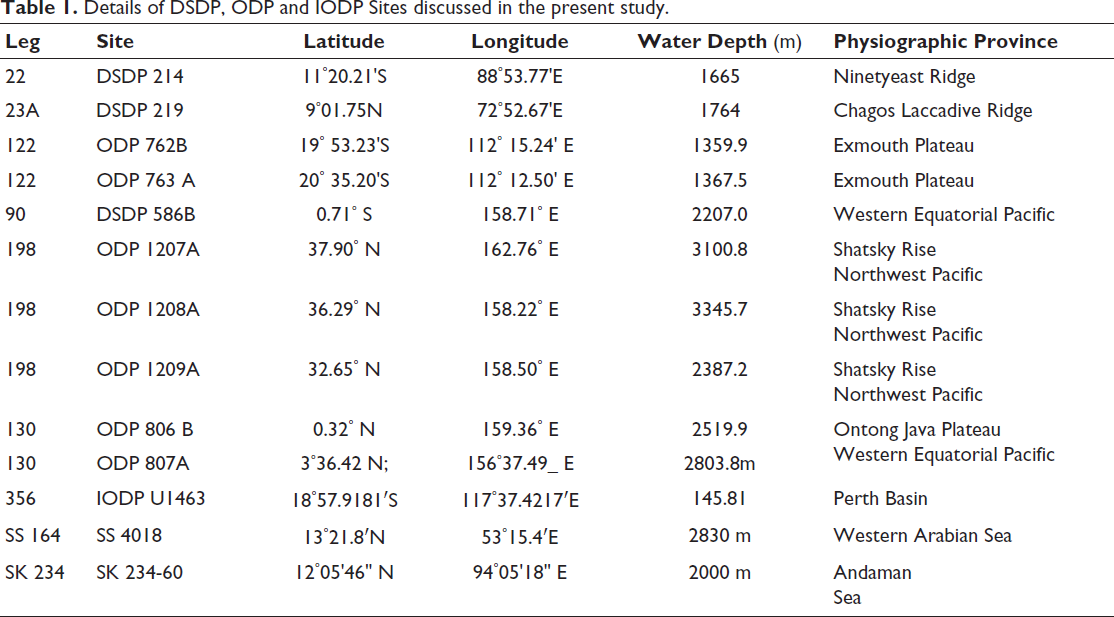
A range of sites was examined, including DSDP Sites 219 and 214 (Northern Indian Ocean); ODP Holes 763A and 762B (Eastern Indian Ocean); Site SS 4018 (Western Arabian Sea); Site SS 234-61 (Andaman Sea); and ODP Hole 807A (Figure 1, Table 1). Additional data on the extinction of G. obliquus and G. extremus were obtained from ODP Holes 1207A, 1208A, and 1209A in the Northwest Pacific (Lam & Leckie, 2020), IODP Site 1463 in the Indonesian Throughflow region (Groeneveld et al., 2021), and a compilation of biochronological records for the Atlantic and Pacific (Berggren, Hilgen, et al., 1995; Berggren, Kent, et al., 1995; Wade et al., 2011). The results show that G. obliquus displays pronounced diachronism in the stratigraphic level of its last occurrence within the Indo-Pacific, whereas the last appearance of G. extremus is nearly synchronous (Table 2).
Location map of the DSDP, ODP and IODP Sites discussed in the present study. Data from the World Ocean Atlas 2023 (Reagan et al., 2024), visualised using Ocean Data View (Schlitzer, 2023).
Details of DSDP, ODP and IODP Sites discussed in the present study.
Numerical age estimates of planktic foraminiferal events at ODP Hole 762B and comparison with published ages from various parts of the Indo-Pacific region. 1. Singh and Sinha (This work); 2. Singh and Sinha (This work); 3. Singh et al. (2021) Composite age – Northern and Eastern Indian Ocean (Calibrated to Gradstein et al., 2012); 4. Sinha and Singh (2008), Eastern Indian Ocean (Calibrated to Berggren, Hilgen, et al., 1995) (Recalibrated-Gradstein et al., 2012); 5. Singh and Sinha (2022) Eastern Indian Ocean (Calibrated to Berggren, Hilgen, et al., 1995) (Calibrated to Ogg, 2020); 6. Groeneveld et al. (2021) (orbitally-tuned age model for IODP Site U1463); 7. Srinivasan and Sinha (1992) (I = Indian Ocean, P = Southwest Pacific) (Recalibrated-Gradstein et al., 2012); 8. Kaushik et al. (2020) Western Pacific (Calibrated to Gradstein et al., 2012); 9. Lam and Leckie (2020) – ODP 1207A, Northwest Pacific (Calibrated to Ogg et al., 2016); 10. Lam and Leckie (2020) – ODP 1208A, Northwest Pacific (Calibrated to Ogg et al., 2016); 11. Lam and Leckie (2020) – ODP 1209A, Northwest Pacific (Calibrated to Ogg et al., 2016); 12. Berggren, Hilgen, et al. (1995); Berggren, Kent, et al. (1995) (Calibrated from Cande & Kent, 1995) (Recalibrated-Gradstein et al., 2012); 13. Wade et al. (2011) (Calibrated to Cande & Kent, 1995) (Recalibrated-Gradstein et al., 2012).
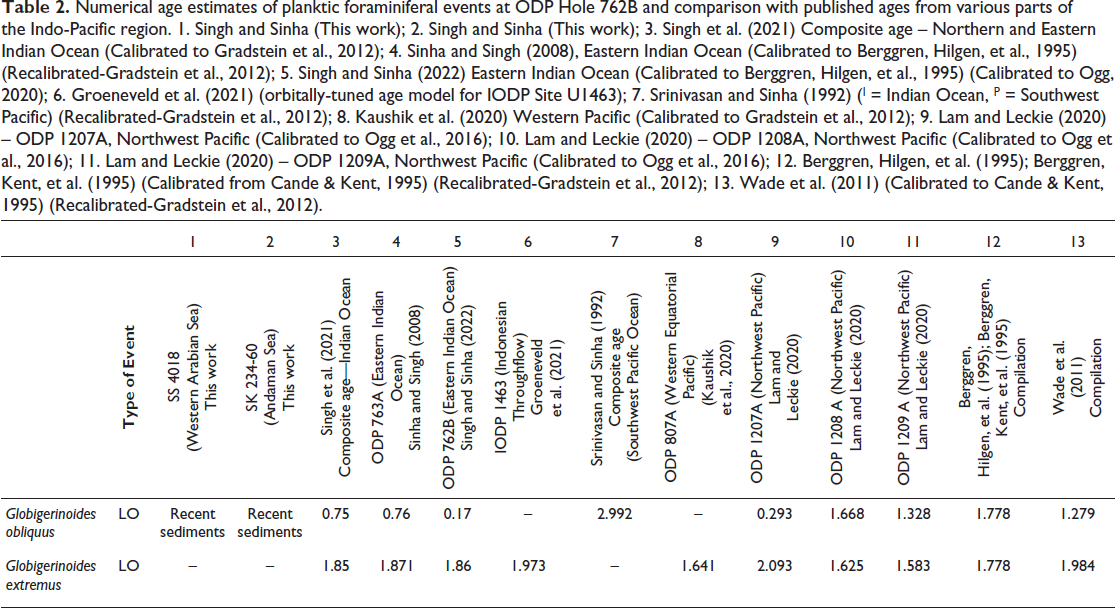
The earliest reported last occurrence (LO) of G. obliquus is from DSDP sites in the Southwest Pacific at ~ 2.9 Ma (Srinivasan & Sinha, 1992). In contrast, Boltovskoy and Watanabe (1975), examining material from the Foraminifera Laboratory collection of the Museum ‘B. Rivadavia’, documented the species in recent sediments of the South Pacific. They identified G. obliquus in Sample 882 (40°S, 109°W, 3,240 m water depth) and Sample 884 (31°S, 109°W, 3,140 m water depth). Abundant Globorotalia truncatulinoides and an absence of Globorotalia tosaensis characterised the associated planktonic assemblage. Given that the last appearance (LA) of G. tosaensis is ~0.54 Ma (Singh & Sinha, 2022), the sediments containing G. obliquus must postdate this interval. Notably, the observation of G. obliquus in recent South Pacific sediments by Boltovskoy and Watanabe (1975) went largely unrecognised in subsequent literature, and the LO of the species continued to be cited near 1.5 Ma, well within the Pleistocene.
Our analysis of several deep-sea core sequences from Site SS 4018 (western Arabian Sea), Site SS 234-61 (Andaman Sea), yielded numerous specimens of G. obliquus from the uppermost samples. These findings support the interpretation of Boltovskoy and Watanabe (1975) that G. obliquus persists in recent sediments. Furthermore, Singh and Sinha (2022) reported the youngest LO of the species at 0.17 million years ago (Ma) from ODP Site 762B in the eastern Indian Ocean.
Numerical age estimates for the LO events of G. obliquus and G. extremus at various DSDP and ODP sites are presented in Table 2 and Figure 2. These data indicate that the LO of G. extremus is broadly synchronous across sites, whereas the LO of G. obliquus is distinctly diachronous (Figure 2).
Diachronism in Globigerinoides obliquus and Globigerinoides extremus, observed at several DSDP, ODP, and IODP sites in the Indo-Pacific region, is shown against Magnetic Polarity Stratigraphy. For the explanation of numbers in the pink circle, refer to the caption of Table 2.
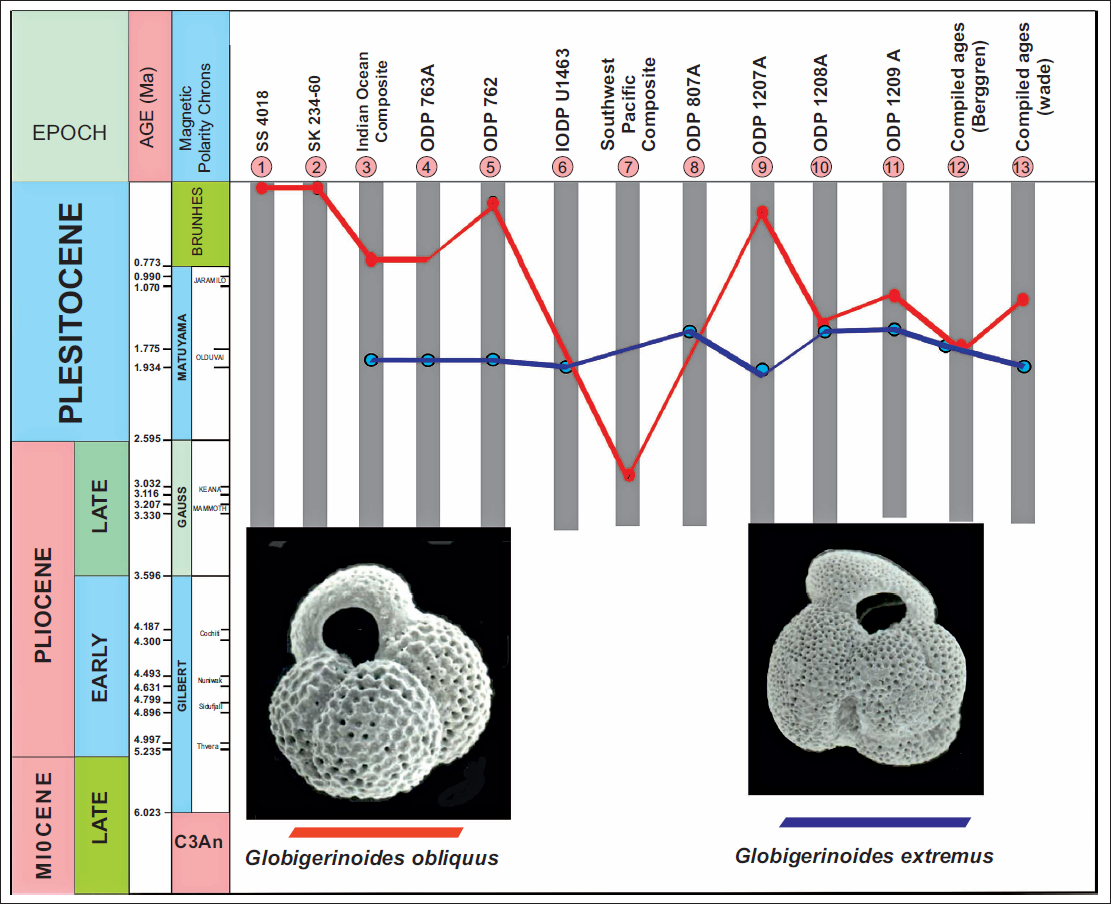
G. obliquus is an open-ocean mixed-layer tropical/subtropical species, with symbionts (Aze et al., 2011; Keller, 1985; Zou et al., 2022), characterised by very heavy δ13C and relatively light δ18O. Vincent (1977) stated that the Last Appearance of G. obliquus and Globigerinoides fistulosus occurs at the same level close to the Pliocene/Pleistocene boundary. Srinivasan and Chaturvedi (1992) at site 214 (Northern Indian Ocean) recorded the first appearance of G. obliquus below the Gr. truncatulinoides FAD. Srinivasan and Sinha (1992) estimated ages of 1.8 Ma for G. obliquus LAD in the Northern Indian Ocean and 2.88 Ma in the Southwest Pacific using the graphic correlation method. The species makes its last appearance at site 762B in the Core 1H, section 2 at 2.31 mbsf. (Singh & Sinha, 2022) and site 763A, the LO of G. obliquus occurs in Core 2H, section 7 at 14.61 mbsf (Singh et al., 2021). Among all ODP and DSDP sites, the LO of this species is youngest at site 762B, with an estimated age of 0.17 Ma. This very young age of G. obliquus LO at this site is intriguing and questions the validity of this event for approximating the Pliocene/Pleistocene boundary (Thunell, 1979). As the earlier definition of the Pliocene-Pleistocene boundary was based on the Top of Olduvai Event ~ 1.6 Ma, many authors, finding G. obliquus and G. extremus LO events close to 1.5 Ma, considered these events important for approximating the boundary. Bolli and Saunders (1985) mentioned a similar-looking species, Globigerinoides elongatus (d’Orbigny, 1826), which occurs even in the Holocene, and cautioned that confusion may arise in distinguishing it from G. obliquus extremus, from which the former is distinguished by a symmetrical primary aperture above the suture between the two earlier chambers.
Singh et al. (2021) and Singh and Sinha (2022) reported the youngest occurrence (LO) of G. obliquus at 0.17 Ma at ODP Hole 762B, while at the nearby ODP Site 763A, it disappears earlier, at 0.75 Ma. Examination of Table 2 and Figure 2 suggests that the LO of G. obliquus is not a reliable biostratigraphic marker, as it is highly diachronous and has even been sporadically recorded in recent sediments of the Indian and South Pacific Oceans. In contrast, the LO of G. extremus appears to be a relatively synchronous event across the Indo-Pacific region.
AN EXAMPLE OF BUDDING CLADOGENESIS (ASYMMETRIC SPECIATION)
The evolution of G. extremus from its ancestor G. obliquus provides a clear example of asymmetric speciation (Stadler et al., 2018), specifically budding cladogenesis. Stratigraphic evidence and previous observations indicate that G. extremus originated from G. obliquus, with both species coexisting for a time before the daughter lineage, G. extremus, went extinct while the ancestor persisted. This evolutionary event occurred in the open-ocean mixed layer. Comparable cases are also documented in deeper-dwelling planktonic foraminifera. For instance, Globorotalia tosaensis (unkeeled) diverged from Globorotalia crassaformis in the Late Pliocene and later gave rise to Globorotalia truncatulinoides (keeled). While the ancestor (G. crassaformis) and the terminal descendant (G. truncatulinoides) survived and remain extant, the intermediate offshoot G. tosaensis became extinct around 0.54 Ma (Singh & Sinha, 2022). Such cases exemplify budding speciation, where a daughter morphospecies branches from an ancestral lineage but eventually disappears, while the ancestor continues as the main lineage.
Palaeoceanographic implications of G. obliquus and G. extremus LO events
Following the Huttonian principle and informed by observations from modern marine sediments, it is inferred that the ecological preferences of Globigerinoides species have remained stable through geological time. Aze et al. (2011) classified both G. obliquus and G. extremus within Ecogroup-1, which comprises symbiont-bearing planktonic foraminiferal taxa typical of open-ocean mixed-layer habitats. In addition, isotopic data support the inclusion of G. ruber in this group as well. To interrogate extinction dynamics within the G. obliquus–G. extremus lineage, we generated census counts of planktonic foraminifera from ODP Hole 763A (Eastern Indian Ocean) and compared the relative abundances of these three key Ecogroup-1 taxa (Figure 3).
The Figure shows that since the Late Pliocene, the ecological niche represented by the open-ocean mixed layer in tropical to subtropical regions characterised by symbiont-bearing taxa with strongly enriched δ13C and relatively depleted δ18O values, classified as Ecogroup 1 (Aze et al., 2011), has been primarily occupied by Globigerinoides ruber. In contrast, Globigerinoides obliquus shows a pronounced decline in abundance, while its descendant Globigerinoides extremus gradually decreases and ultimately goes extinct. This major turnover, marked by the dominance of G. ruber and the decline or disappearance of G. obliquus and G. extremus, is coeval with the onset of Northern Hemisphere Glaciation (NHG) and likely reflects a fundamental reorganisation of upper-ocean structure. The Figure also illustrates competition between G. ruber and G. obliquus + G. extremus for dominance of the Ecogroup-1 niche (open-ocean mixed layer in tropical to subtropical waters). It supports Ricklefs’ (1979) hypothesis that species abundance is determined not only by interactions with the physical environment (habitat) but also by competition with other species in overlapping niches. During the Late Miocene–Early Pliocene, competition between G. obliquus and G. extremus for the Ecogroup-1 niche is evident in their opposite trends in relative abundance, represented by blue shaded bars.
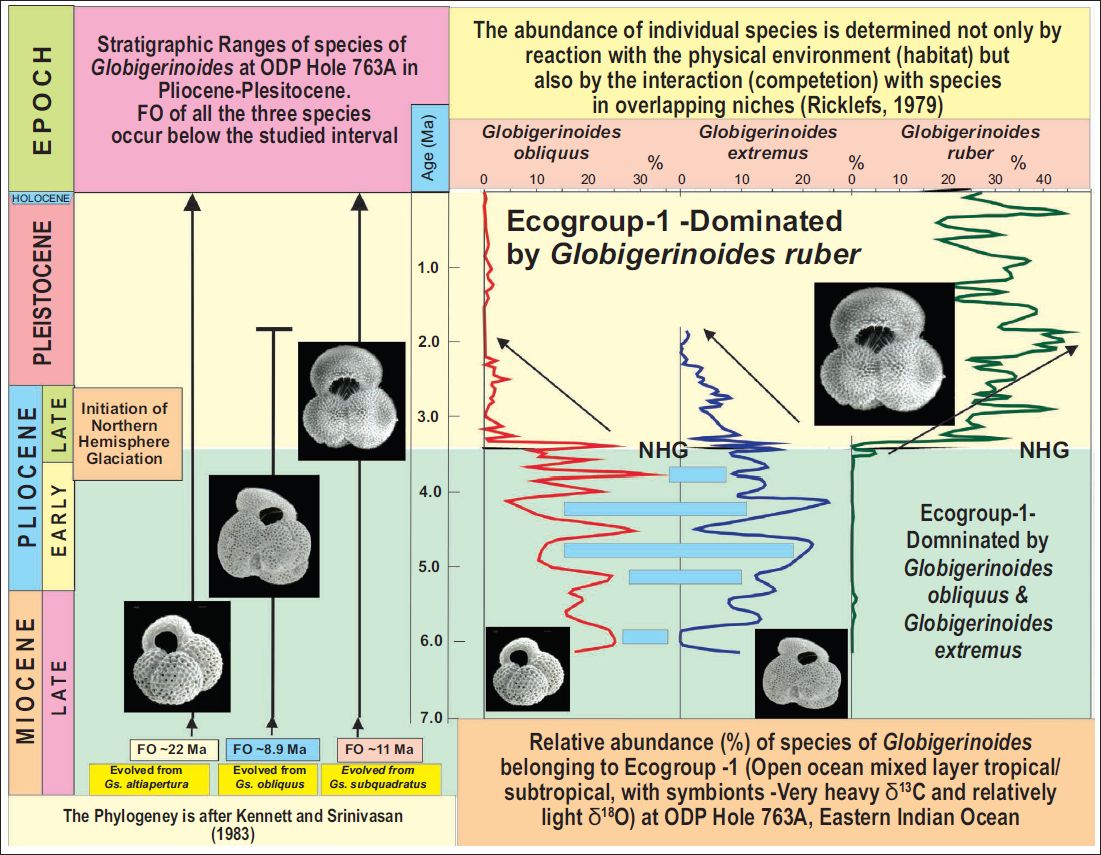
Our analysis reveals a pronounced faunal turnover. In the Late Miocene to Early Pliocene interval, G. obliquus and G. extremus were predominant, while G. ruber was comparatively rare. However, beginning just prior to 3 Ma, coincident with the inception of Northern Hemisphere Glaciation (NHG), a striking transition is evident as both G. obliquus and G. extremus exhibit rapid population declines, culminating in the extinction of G. extremus around 1.8 Ma. Concurrently, G. ruber rises sharply in abundance, achieving numerical dominance. This significant restructuring of the foraminiferal assemblage likely reflects a major reorganisation of the upper-ocean mixed layer in response to NHG.
Figure 3 further illustrates competitive interactions among Ecogroup-1 taxa. During the Late Miocene to Early Pliocene, inverse abundance trajectories for G. obliquus and G. extremus are consistent with direct competition within a shared ecological niche. Following the onset of NHG, G. ruber appears to have competitively displaced both taxa, aligning with theoretical models (Ricklefs, 1979) suggesting that both environmental factors and interspecific competition within overlapping niches influence species abundances. Although census data from additional sites described in this study are lacking, it is reasonable to suggest that the observed diachroneity of the last occurrence (LO) of G. obliquus could be modulated by local palaeoceanographic variability in the upper water column. Sporadic occurrences may also give the impression of pseudo-extinction, particularly when recovery frequencies are very low. In contrast, the extinction of G. extremus, unable to persist in competition with other Ecogroup-1 taxa, appears broadly synchronous across broad geographic areas and likely reflects global restructuring of upper-ocean stratification.
In addition to its classification as an Ecogroup-1 taxon, G. obliquus prefers oligotrophic surface waters. Gupta and Srinivasan (1992) documented high surface-water productivity around 2.4 million years ago (Ma) in the South Equatorial Indian Ocean near the Ninetyeast Ridge, based on the abundance of Uvigerina. The relatively early disappearance of G. obliquus, at approximately 2.3 Ma in the Northern Indian Ocean, may be attributed to enhanced productivity or eutrophic conditions. Similarly, at DSDP Hole 219 (Northern Indian Ocean), Gupta and Thomas (1999) reported highly productive, oxygen-poor environments between 2.4 and 1.8 Ma.
Singh et al. (2021) proposed an explanation for the later survival of G. obliquus (to 0.17 Ma) at ODP Site 762B in the Eastern Indian Ocean. The survival of G. obliquus to later intervals at ODP 762B may correspond to localised favourable conditions, akin to those experienced by G. ruber. Notably, along the Western Australian margin (ODP Holes 763A and 762B), persistent northward winds do not induce upwelling due to the mitigating influence of the Leeuwin Current (Sinha et al., 2006). Here, episodic upwelling has been documented (Sinha & Singh, 2007; Sinha et al., 2006). The delayed extinction (LO) of G. obliquus in the eastern Indian Ocean (Table 2; Sinha & Singh, 2008; Singh & Sinha, 2022) is therefore attributable to the persistence of oligotrophic conditions and a thick, warm mixed layer.
In summary, the present findings indicate that previous interpretations of synchronous extinction at the G. obliquus LO in the Indo-Pacific are contradicted. Instead, the diachrony of extinction across this region is most plausibly explained by regional oligotrophic or trophic state variations controlling the stratigraphic distribution of this taxon.
CONCLUSIONS
In the present study, multiple deep-sea cores from the Indo-Oceanic Region (IOR) were examined, with a particular focus on G. obliquus. We report the highest stratigraphic occurrence of the last appearance datum (LAD) of G. obliquus in the IOR. The taxonomic identity of the specimens is confidently confirmed, excluding any confusion with G. elongatus, which is characterised by a cancellate surface and less distinct spinosity. Based on our results, we propose that, contrary to previous reports suggesting that G. obliquus went extinct in the mid-Quaternary, this species actually persisted through the Holocene, exhibiting significant diachrony in its last occurrence across various Indo-Pacific sites.
This study further highlights a pronounced diachronous extinction pattern of G. obliquus in the ocean, likely linked to palaeoceanographic parameters such as mixed-layer oligotrophic conditions. Consequently, the stratigraphic placement of the last occurrence datum of G. obliquus in the IOR requires revision. Given its diachronic disappearance, the use of G. obliquus LO as a reliable stratigraphic marker is called into question.
In contrast, the relatively synchronous extinction of G. extremus likely reflects its competitive exclusion by other Ecogroup-1 Globigerinoides species, in addition to large-scale reorganisation of the oceanic water column structure from the Late Pliocene onward.
The shifting dominance patterns within Ecogroup-1 before and after the onset of NHG suggest a significant alteration in mixed-layer hydrography. G. ruber emerged as the dominant member of Ecogroup-1 following NHG initiation, while the abundances of G. obliquus and G. extremus gradually declined, with G. extremus becoming extinct around 1.8 Ma. These observations support Ricklefs’ (1979) conceptual framework, in which species sharing a common ecological niche not only respond to environmental variation but also engage in competitive interactions within overlapping niches.
Footnotes
Acknowledgements
AKS and DKS thank the Department of Geology, University of Delhi, for their infrastructural support. AKS conceived the research problem and wrote the manuscript. DKS helped draft the manuscript and offered constructive suggestions. AKS and DKS also thank the Ministry of Earth Sciences’ Palaeoclimate Programme, Government of India, for their financial support under the Palaeoclimate Programme (Sanction No. MoES/CCR/Paleo-4/2019).
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This research was supported by the Ministry of Earth Science Palaeoclimate Programme, Government of India (Sanction No. MoES/CCR/Paleo-4/2019).