
Abstract
Benthic foraminifera assemblages from two fjords of Svalbard—Kongsfjorden and Krossfjorden in the High Arctic—have been studied. The foraminifera assemblages show three characteristic assemblages from the two fjords’ glacial proximal, middle and distal parts. The Cassidulina reniforme–Elphidium clavatum, Textularia spp. assemblage dominates the foraminifera fauna near the glacier proximal that indicates increased sedimentation and meltwater runoff; a Nonionellina labradorica–Islandiella helenae–Lobatula lobatula assemblage relates to well-oxygenated and high energy conditions and mainly occupies the middle part; and the Nonionellina labradorica–Islandiella helenae assemblages dominate in the distal region that indicates high surface primary productivity. Krossfjorden contains an oxygen deficiency zone around the middle part characterised by Stainforthia loeblichi. The abundance of Islandiella helenae and Nonionellina labradorica indicates that the high surface primary productivity of Krossfjorden is more than that of Kongsfjorden.
Introduction
The Arctic Ocean plays a major role in global ocean circulation and has undergone significant climatic changes in recent decades (Racine et al., 2018). While notable investigations related to climatic changes in different parts of the Arctic region have been carried out (Mangerud et al., 1998), further studies will enhance the knowledge of the characteristics of the past environment. Studies based on the present distribution of benthic foraminifera provide a better understanding of the region’s environment (Lloyd et al., 2006). Benthic foraminifera assemblages in the Arctic region have been significantly used as a valuable proxy for estimating Quaternary palaeoclimatic changes (Bergsten et al., 1998; Chauhan et al., 2014; Feyling-Hanssen & Ulleberg, 1984; Hovland 2014; Kubischta et al., 2010; Lycke et al., 1992; Miller et al., 1989). Due to their greater preservation potential, abundance and diversity in marine waters, shelf areas and fjords (Jernas et al., 2018), they are helpful, particularly when studying long records of climate changes along with other records, that is, sedimentology and chronology and accessibility of certain areas around the Arctic region is challenging (Husum et al., 2015). Studying the ecology and habitat of modern benthic foraminifera assemblages denotes the characteristics of the associated marine environment and, therefore, can be used as an analogue for interpreting past environments (Hald & Steinsund, 1992; Jennings & Helgadottir, 1994).
Much of the relevant modern benthic foraminiferal investigations have been concentrated in various parts of the Arctic region, adjacent to the Arctic region and Barents Sea (Hald and Korsun 1997; Hald & Steinsund, 1992; Husum et al., 2015; Korsun & Hald 1998, 2000; Korsun et al., 1995; Lagoe 1977; Osterman et al., 1999; Polyak et al., 2002; Racine et al. 2018; Saher et al., 2009, 2012; Schrӧder Adams et al., 1990; Wollenberg & Mackensen, 1998). Previous studies on Kongsfjorden constitute annual environmental changes based on benthic foraminifera populations (Jernas et al., 2018), the morphology of benthic foraminifera (Saraswat et al., 2018) and palaeogeochemical analysis and determination of benthic environments (Loubere et al., 2011). Previous studies on Krossfjorden constitute works by Hald and Korsun (1997) and Saraswat et al. (2018).
This article presents the investigations of foraminiferal populations collected from the surface sediments of two fjords, namely Kongsfjorden and Krossfjorden, in the north-western part of the Svalbard Archipelago, High Arctic. The modern environment, that is, the interaction between water discharged from the glaciers as well as sea ice and the intrusion of warm Atlantic waters (AW) in these two fjords, may approximate Quaternary glaciomarine settings (Hald & Korsun, 1997). The main objective of our study is to represent detailed information on recent Arctic foraminifera distribution and the surrounding environment from the inner to the outer ends of two fjords, Kongsfjorden and Krossfjorden. The study reveals baseline data that can be used as an analogue for fossil assemblages of the Quaternary and to reconstruct past environments of the Kongsfjorden area.
Study Area
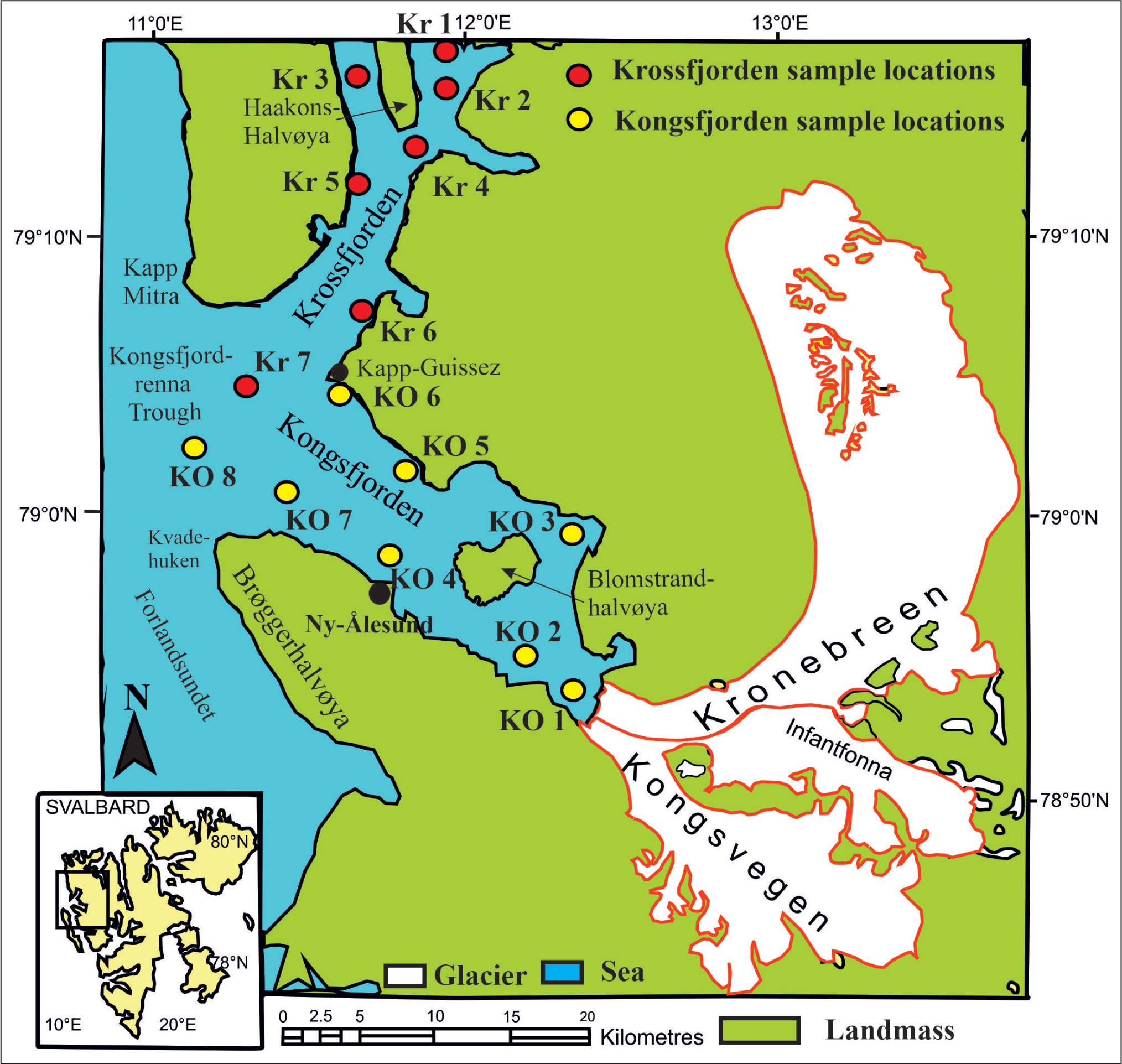
The Kongsfjorden and Krossfjorden are two fjord systems (Figure 1) situated between 78°40′ and 77°30′N and 11°3′ and 13°6′E on the north-western coast of Svalbard. The fjords comprise two submarine trenches, which converge to the deep glacial trough Kongsfjordrenna, covering an area of 3,074 km2 (Howe et al., 2003). Kongsfjorden is 20 km long with a varying width from 4 to 10 km, expanded up to the end between Kvadehuken and Kapp Guissez, with an estimated total volume of 29.4 km³ (Husum et al., 2019). Krossfjorden is about 30 km long, 3–6 km wide, divided into two minor fjords with 100 m depth by Haakonshalvøya, with an estimated 25 km³ (Howe et al., 2003). The area that embraces Kongsfjorden and Krossfjorden is situated at the tectonic boundary between the Tertiary fold belt and thrust sheet in the south-west and is composed of metamorphic rocks older than the Tertiary rocks in the north-east (Howe et al., 2003). The northern part of Kongsfjorden consists of medium-grade metamorphic rock of the Proterozoic age as the basement rock. Marvels from the Devonian age occur on the nearby Brømstrandvøya island. East of Kongsfjorden comprises ‘Old Red Sandstone’ of Devonian age overlies the Northern Basement Province (Husum et al., 2019). Palaeozoic conglomerates, sandstones and Proterozoic metamorphic phyllite of the Kongsvegen group lie to the south of the Kongsfjorden area. Near Ny-Ålesund, a Tertiary fold and thrust belt fragment exists as the Ny-Ålesund group, containing Palaeogene coal and conglomerates (Husum et al., 2019).
Oceanographic conditions in the Kongsfjorden–Krossfjorden have been strongly related to the features of the currents flowing around the Svalbard archipelago (Svendsen et al., 2002). The fjords are characterised by two dominant water masses—Arctic Waters (ArW) and warm AW. AW follows northward as the West Spitsbergen Current (WSC), occupying the upper ca. 600 m of that current (Svendsen et al., 2002). When entering the Kongsfjorden, AW mixes with Arctic coastal waters and is designated as Transformed Atlantic Water (TAW; Howe et al., 2003). Polar, fresh and cold ArW from the Barents Sea flows parallel to the WSC. When the front between warm Spitsbergen and cold Arctic Waters (ArW) weaken, Atlantic Waters (AW) enters the Kongsfjorden fjord (Husum et al., 2019). During summer, AW enters Kongsfjorden and stays for a few months. During winter, the generation of the polar front between Atlantic and ArW restricts the inflow of AW in the fjord. The sea ice production at the innermost part of the fjord and the release of brine water cause dense upper water mass formation (Cottier et al., 2010), resulting in vertical exchange and reviving water masses in the fjord (Husum et al., 2019).
Location map of Kongsfjorden and Krossfjorden (After Howe et al., 2003 and Husum et al., 2019).
Materials and Methods
Sampling
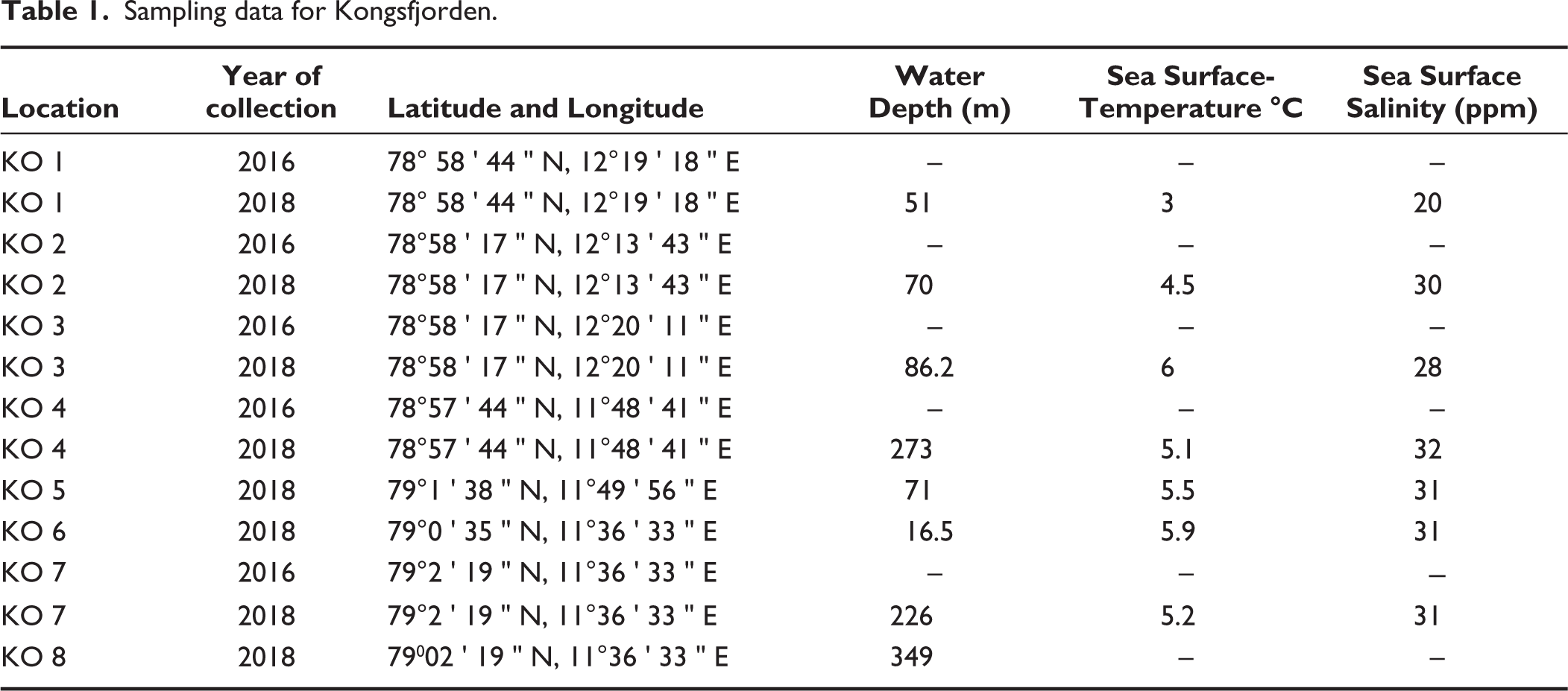
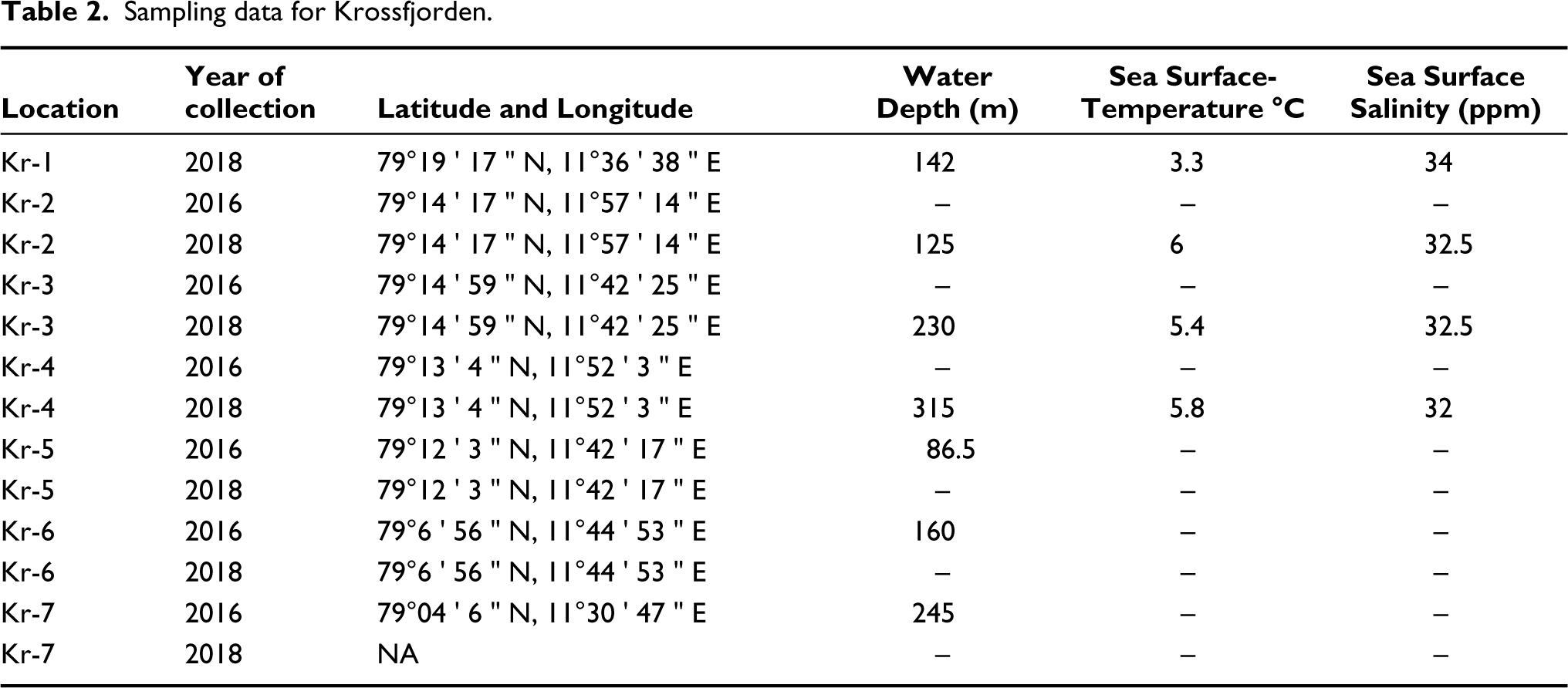
Eight surface sediment samples were collected from Kongsfjorden and seven sediment samples from Krossfjorden (Figure 1) along a transect from the proximal to the distal part of both fjords during the summer months of August–September in 2016 and 2018. During sampling, physical oceanographic parameters such as temperature, salinity and depth were calculated with a Conductivity–Temperature–Depth (CTD) profiler at each sampling station in Kongsfjorden and Krossfjorden (Tables 1 and 2). The Van Veen Grab sampler has retrieved the samples. The top sediment (0–2 cm) was transferred into a 250 ml plastic bottle and stained with Rose Bengal solution (2 g of rose Bengal powder mixed homogeneously in 1 l ethanol) to identify the living (stained) fauna. The stained samples were brought back to India and processed in the micro-palaeontological laboratory of Jadavpur University.
Sampling data for Kongsfjorden.
Sampling data for Krossfjorden.
Comparative study of Kongsfjorden and Krossfjorden.

Laboratory Analyses
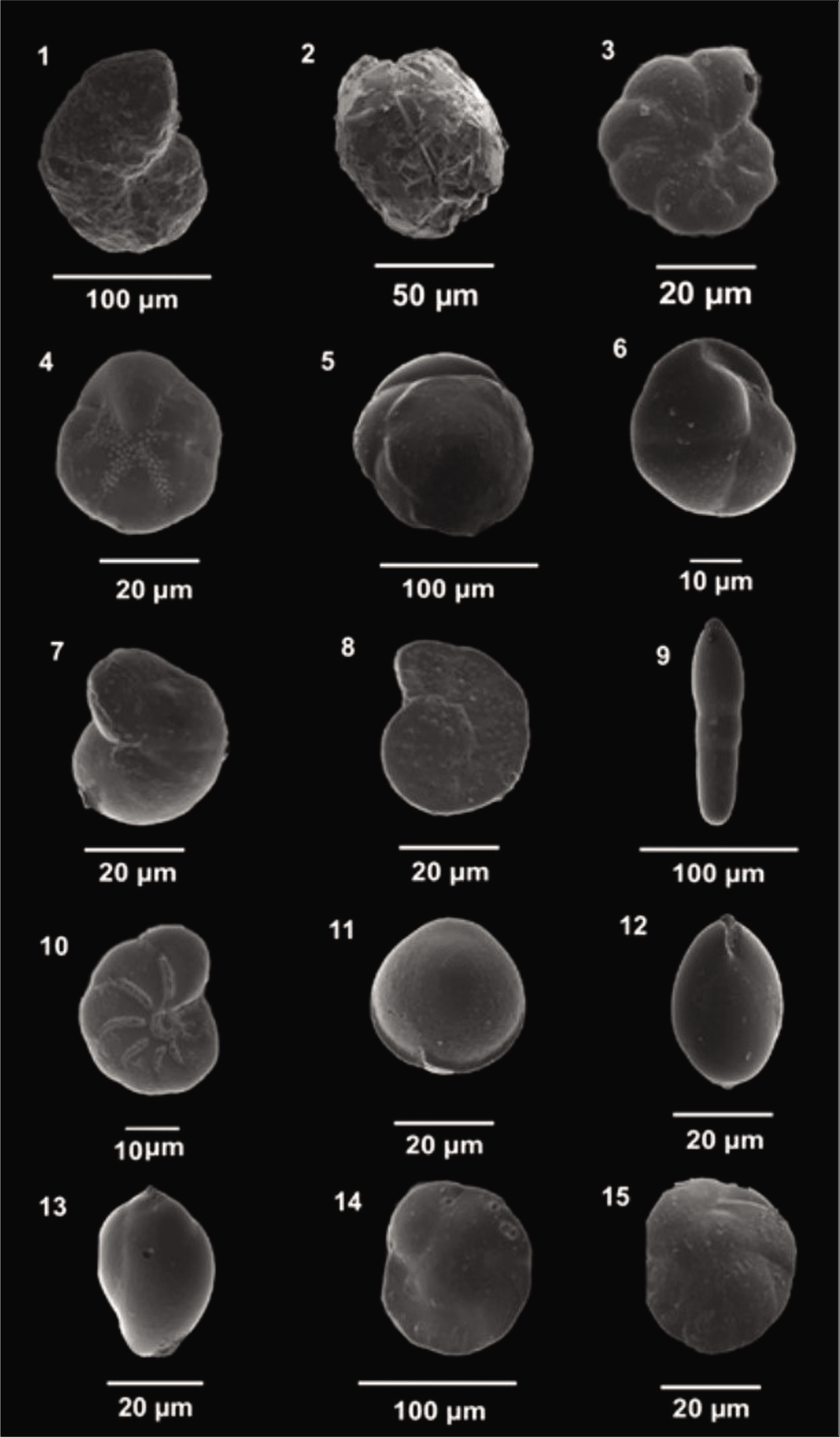
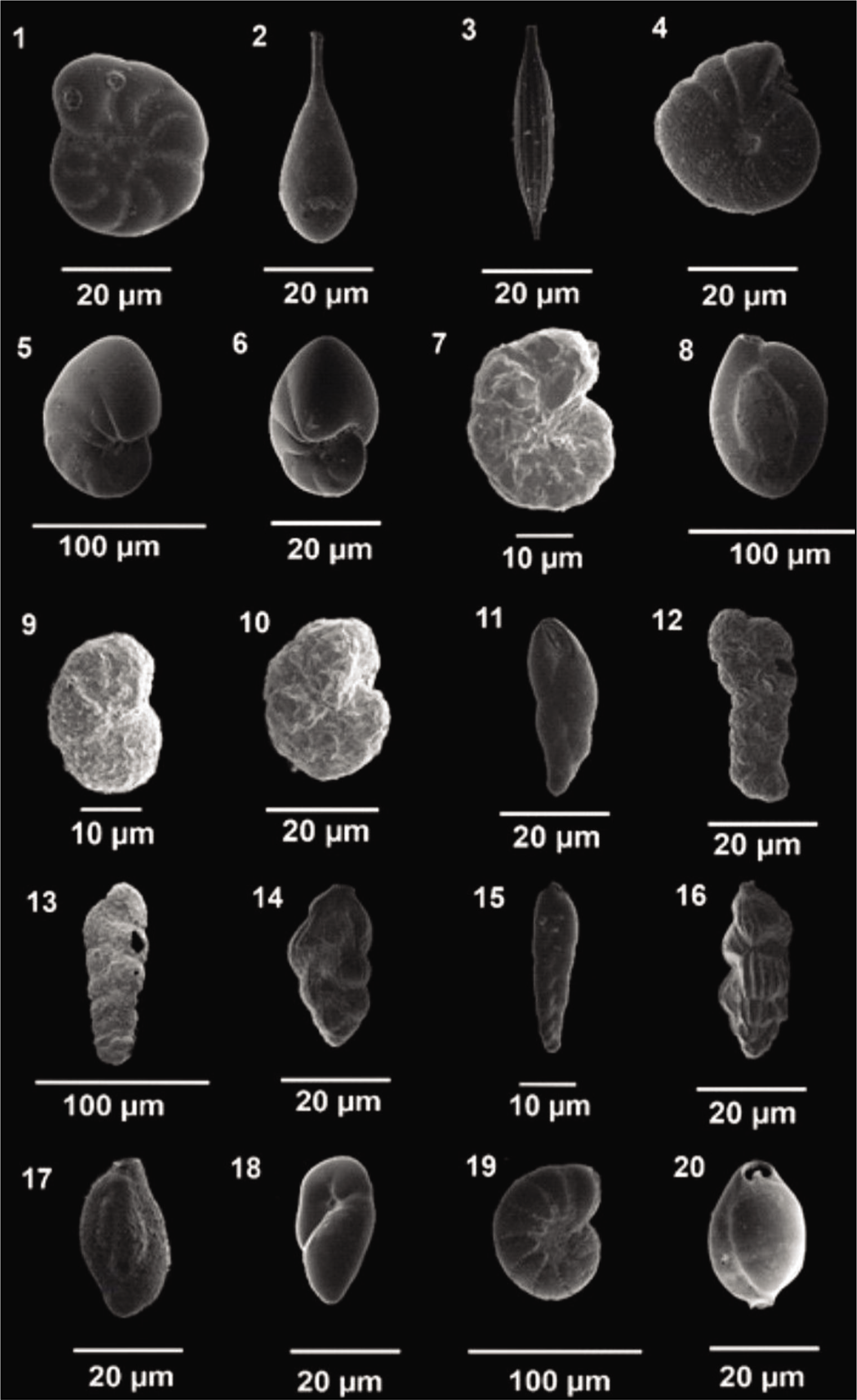
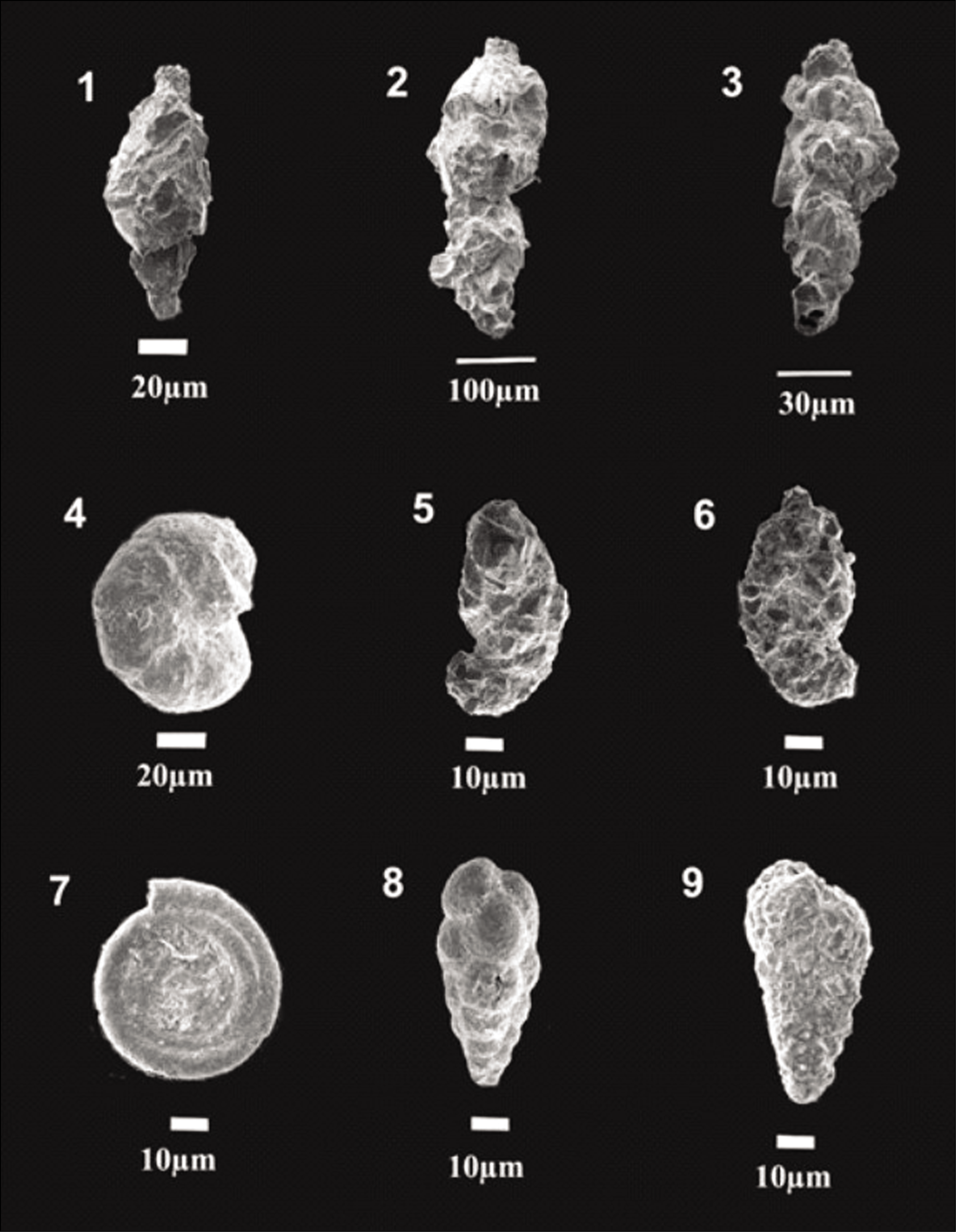
The collected samples were washed using a 63 µm sieve to remove the clay and silt fractions. The ≥63 µm fraction was oven-dried at 50 °C till the sample dried completely. The dried sediment was put in a micro-splitter, and 1 gm of each weighted sample was observed under a stereo zoom microscope (Nikon SMZ 1000). Tests of both stained and non-stained benthic foraminifera together were picked directly from 1 gm of the dry sediment using a thin, damp brush and placed into 24 chambers of micro-palaeontological slides. Foraminifera tests were counted (living + dead) as Total foraminifera number (TFN), while living foraminifera (stained tests) were counted separately. The details of the foraminifera analysis of both fjords are shown in Table S1. Later, they were identified up to the species level with the help of the works of Loeblich and Tappan (1987), Feyling-Hanssen (1964), Korsun et al. (1995) and Hald and Korsun (1997). The best-preserved tests were selected and photographed using a scanning electron microscope (ZEISS EVO 18; Plates 1–3).
Scanning electron microscope images of Arctic Foraminifera . 1. Labrospira crassimargo; 2. Adercotryma glomeratum; 3. Astrononion hamadaense; 4. Buccella frigida (U); 5. Buccella frigida (Sp); 6. Cassidulina reniforme; 7. Lobatula lobatula (U); 8. Lobatula lobatula (Sp); 9. Dentalina sp.; 10. Elphidium clavatum; 11. Fissurina sp.; 12,13. Globobulimina sp. (S); 14. Islandiella norcrossi (Sp); 15. Islandiella helenae (Sp); U, umbilical view; Ap, apertural view; Sp, spiral view; S, side view.
Scanning electron microscope images of Arctic Foraminifera . 1. Haynesina orbicularis, (Si); 2,3. Lagena sp., (Si); 4. Melonis affinis, (Si); 5,6. Nonionellina labradorica (Si, Ap); 7. Portatrochammina sp. (Si); 8. Quinqueloculina sp. (Si); 9,10. Recurvoides turbinatus, (Si); 11. Stainforthia loeblichi (Ap); 12. Spiroplectammina biformis (Si); 13. Textularia earlandi, (Si); 14. Trifarina fluens, (Si); 15. Bolivinellina sp., (Si); 16. Uvigerina sp.; 17. Miliammina fusca; 18. Robertina sp.; 19. Elphidium bartletti, (Si); 20. Pyrgo sp.
Scanning electron microscope images of Arctic Foraminifera . 1–3. Reophax sp.; 4. Portatrochammina bipolaris; 5, 6. Ammotium sp.; 7. Ammodiscus sp.; 8. Eggerella sp.; 9. Textularia torquata.
Results
Kongsfjorden
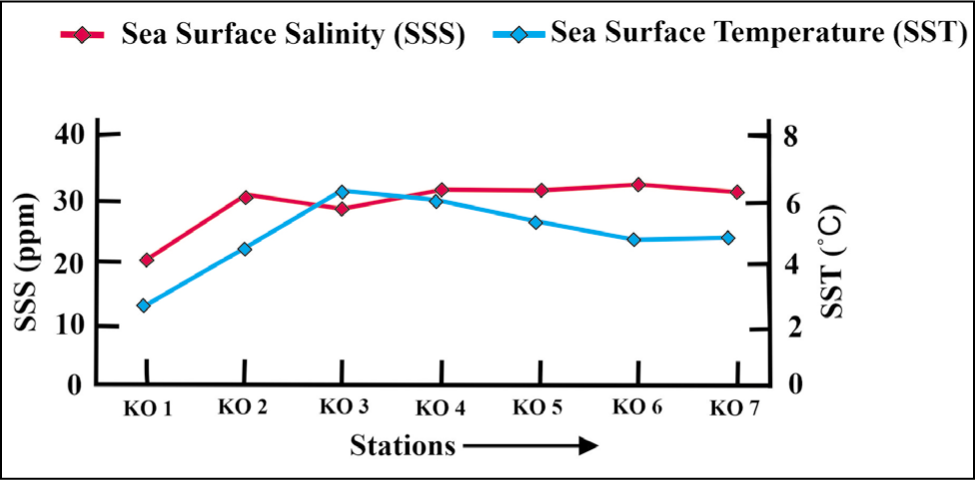
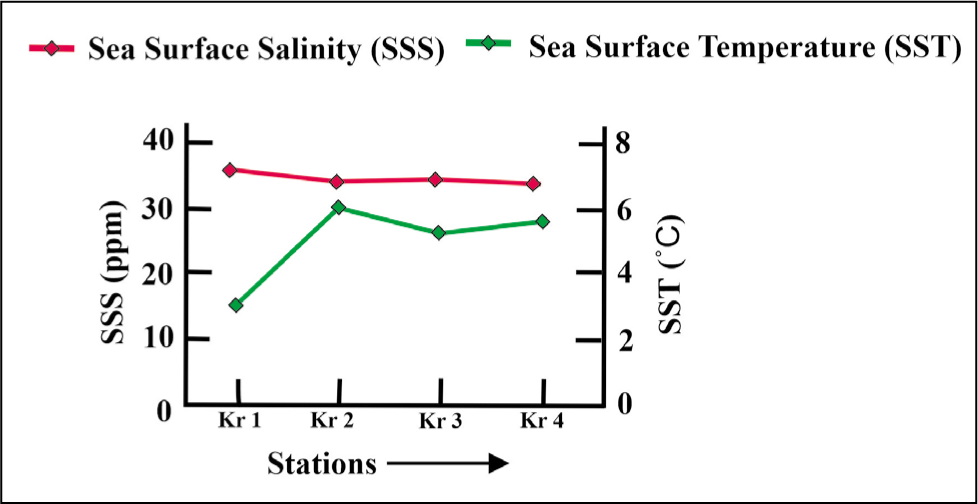
The sea surface temperature of Kongsfjorden varies from 3 °C to 6 °C, and salinity ranges from 20 to 32 ppm from the inner to outer fjord from August to September. Due to strong wind flow and sea turbulence, temperature and salinity were not measured for the distal station KO 8 (Figure 2).
Sea surface salinity and sea surface temperature plot for Kongsfjorden.
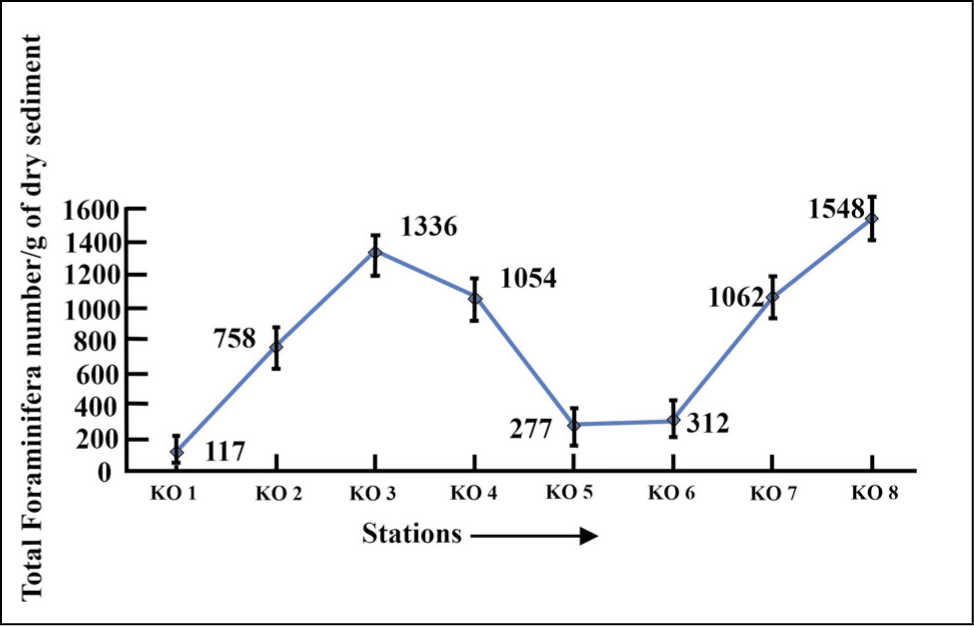
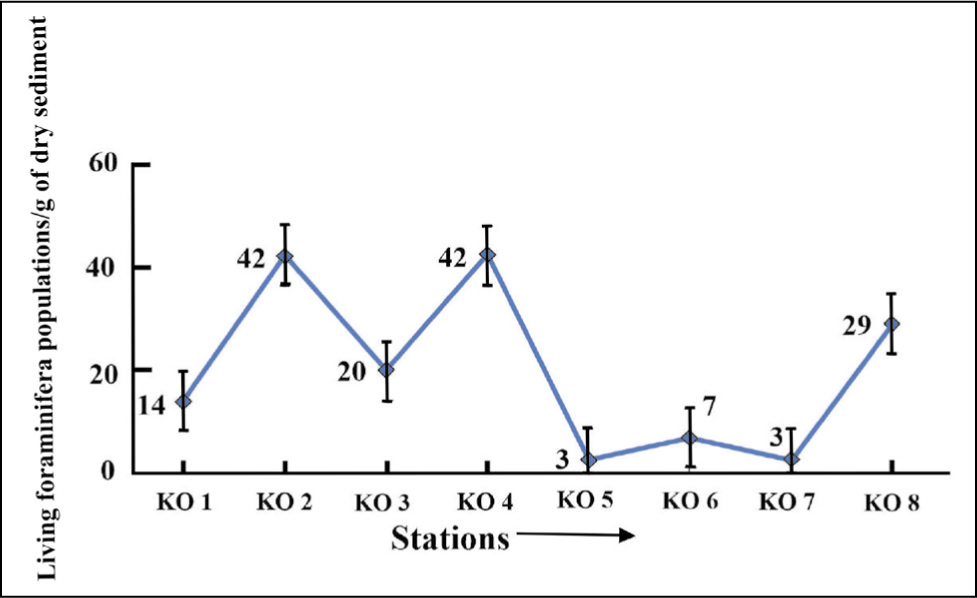
A total of eight samples from the glacial proximal (namely KO 1, KO 2 and KO 3), middle part (namely KO 4 and KO 5) and glacial distal (namely KO 6, KO 7 and KO 8) have been investigated. The TFN increases from the proximal part (TFN-117 in KO 1) to the distal part (TFN-1548 in KO 8) (Figure 3), and a moderate abundance of living foraminifera has been observed in the glacial proximal, middle and distal part (Figure 4).
Total foraminifera number (TFN) plot for Kongsfjorden.
Living foraminifera plot for Kongsfjorden.
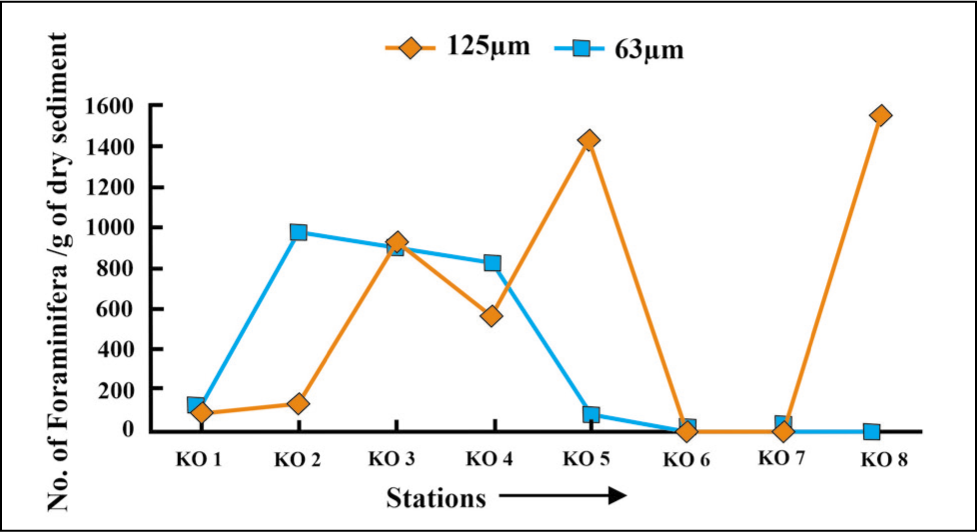
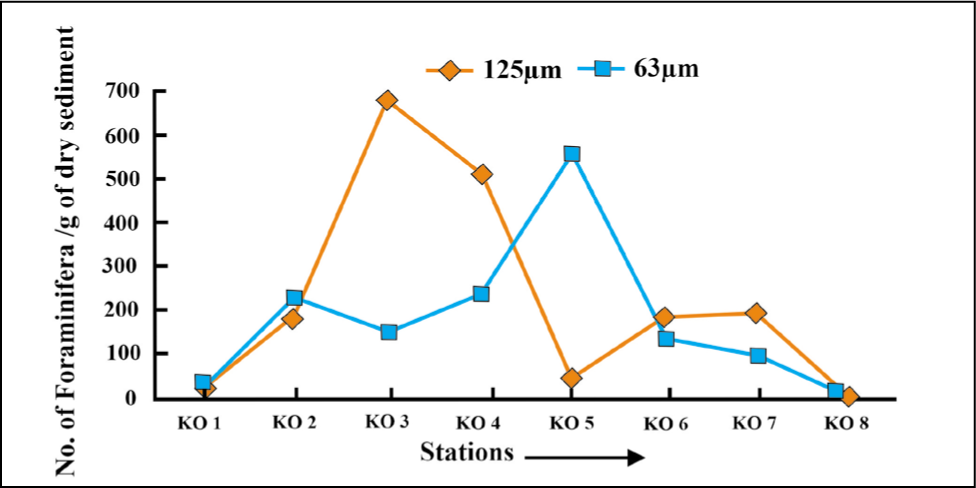
At the distal part of the fjord, foraminifera specimens >125 µm are more abundant compared to those >63 µm, which are high in proximal to the middle part (Figures 5 and 6).
Benthic foraminifera size variation in Kongsfjorden 2016.
Benthic foraminifera size variation in Kongsfjorden 2018.
Dominant benthic foraminifera abundance percentage in Kongsfjorden.
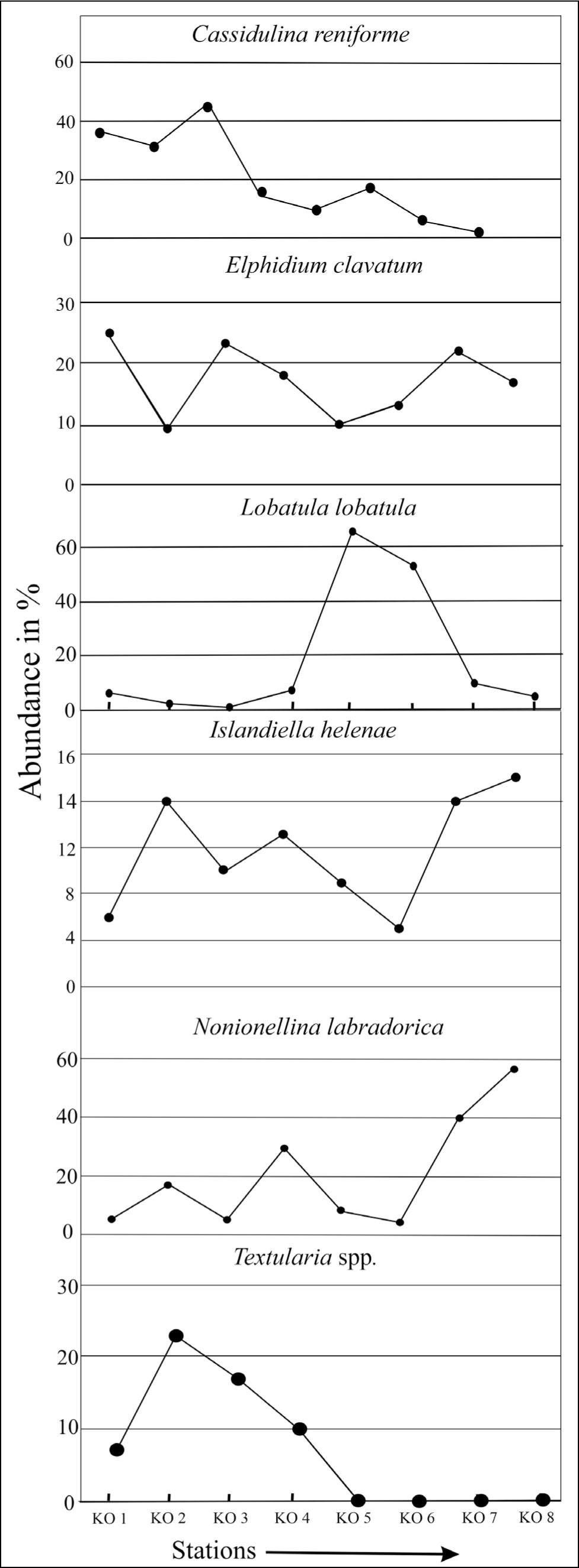
The fjord is characterised by calcareous hyaline forms, Cassidulina reniforme (42%–57%) and Elphidium clavatum (22%–25%), assemblages in the proximal part. The middle and distal fjords contain foraminifera assemblages that constitute Nonionellina labradorica (25%–58%), Islandiella helenae (12%–16%) and Lobatula lobatula (20%–60%), with the specimens of Nonionellina labradorica comprising the bulk of the foraminiferal fauna (58% of TFN in sample KO 8). Among the agglutinated specimens, that is, Textularia spp. (10%–23%) and Spiroplectammina biformis (8%) widely occur near the glacier to the middle of the fjord. Labrospira crassimargo and Adercotryma glomeratum are the other two characteristic agglutinated species in the middle of the fjord.
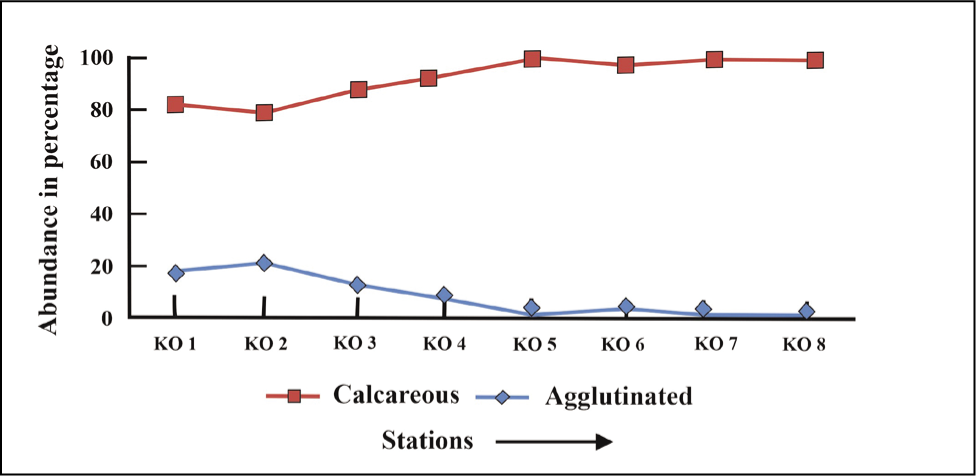
Agglutinated foraminifera are characteristic in the proximal and middle regions, while calcareous forms increase from the glacial middle to the distal part (Figure 8).
Calcareous and agglutinated foraminifera plot for Kongsfjorden.
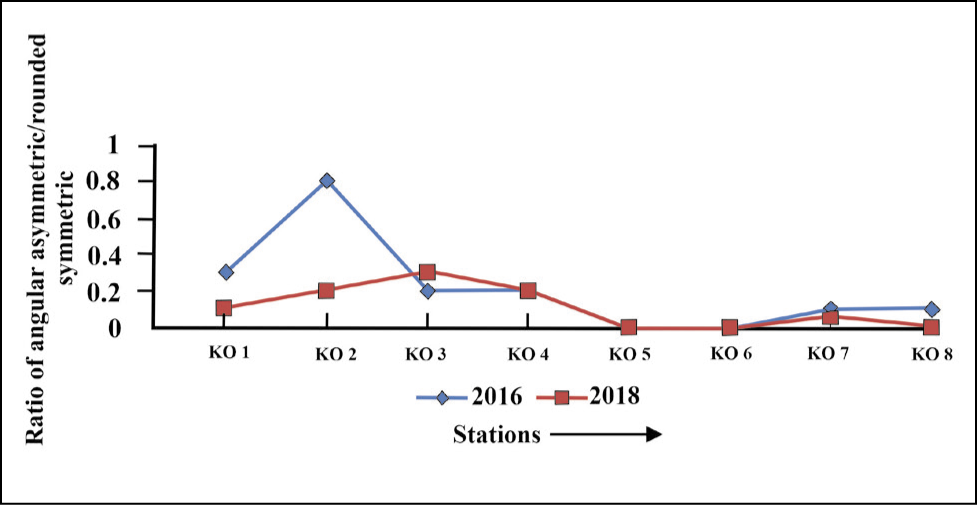
The ratio of angular asymmetric to rounded symmetric forms shows a high value (0.3) in the proximal part (Figure 9) and gradually diminishes towards the distal part (0.1).
Ratio of angular asymmetric to rounded symmetric plot for Kongsfjorden.
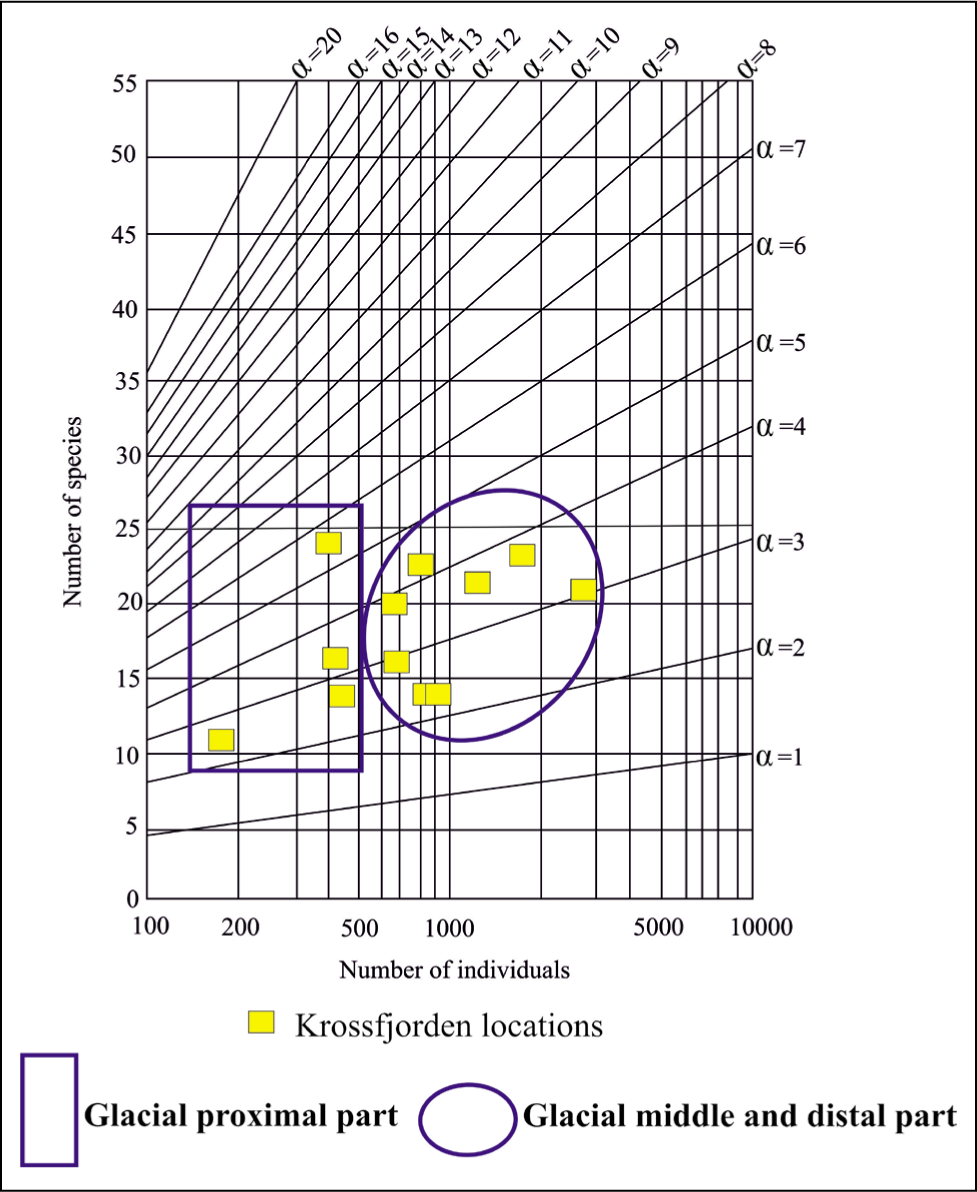
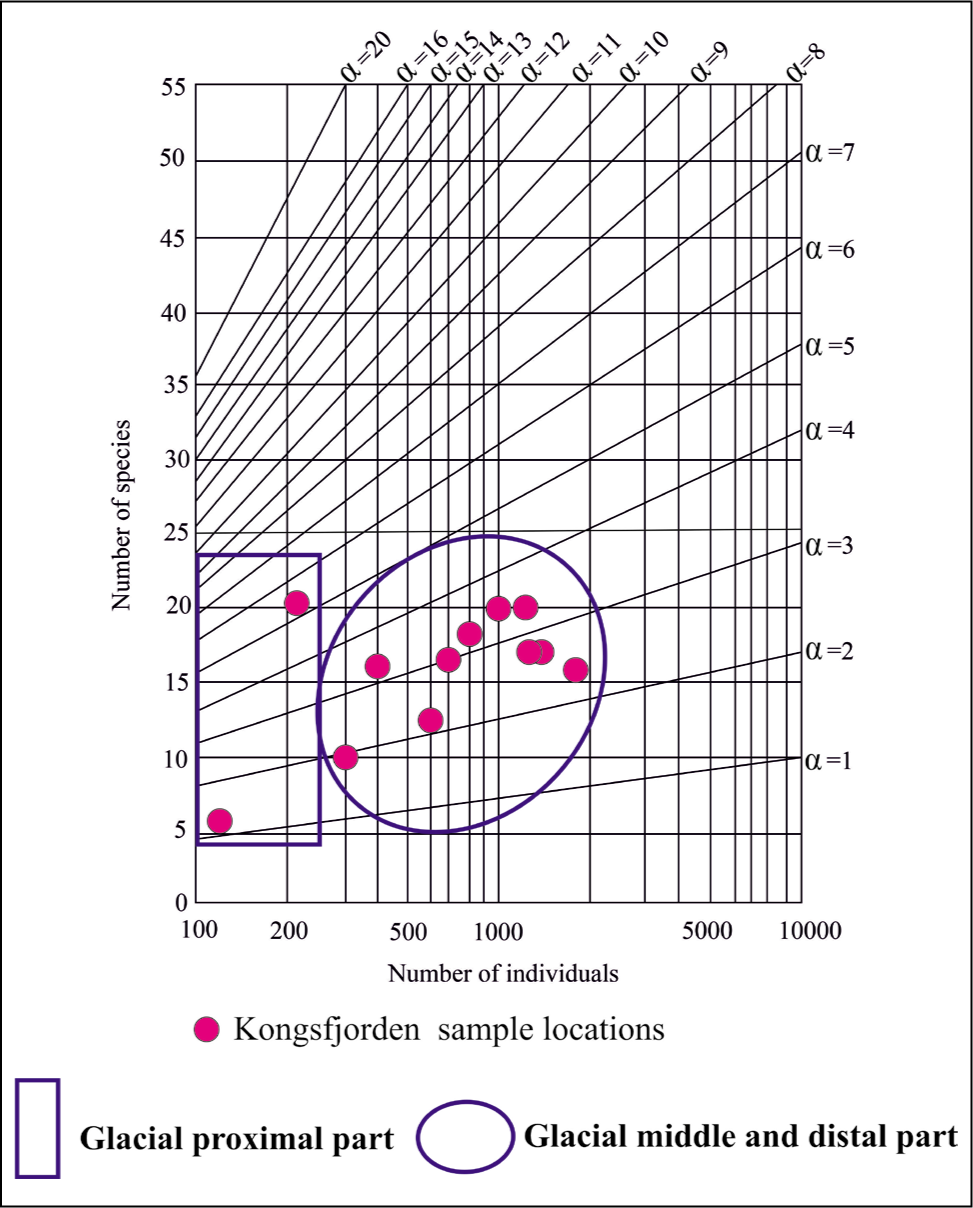
The Fisher alpha diversity index of foraminifera in Kongsfjorden varies from 2 in the proximal part to 4 in the middle and distal parts. The value indicates that foraminifera from the distal to middle fjords are highly diverse compared to the proximal part (Figure 10).
The fishers alpha diversity index for Kongsfjorden.
Krossfjorden
The surface temperature of Krossfjorden, recorded in the immediate vicinity of the glacier, is 3 °C and reaches 6 °C towards the distal part. Surface salinity starts at 34 ppm near the glacier and 32 ppm in the middle and distal parts (Figure 11). Data for temperature and salinity have been recorded at only four stations and not at others due to the heavy wind and turbulence of the sea on the sampling day.
Sea surface salinity and sea surface temperature plot for Krossfjorden.
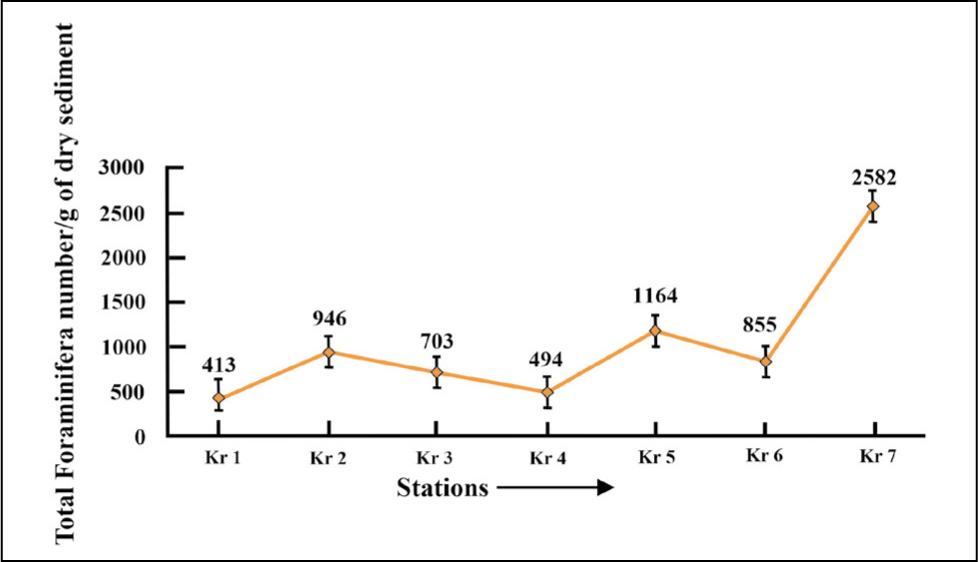
A total of seven samples from the proximal (namely Kr 1, Kr 2 and Kr 3), middle parts (namely Kr 4 and Kr 5) and distal parts (namely Kr 6 and Kr 7) of Krossfjorden have been investigated. The TFN increases from the proximal (TFN = 413 in Kr 1) to the distal part (TFN = 2582 in Kr 7) (Figure 12).
Total foraminifera number (TFN) plot for Krossfjorden.
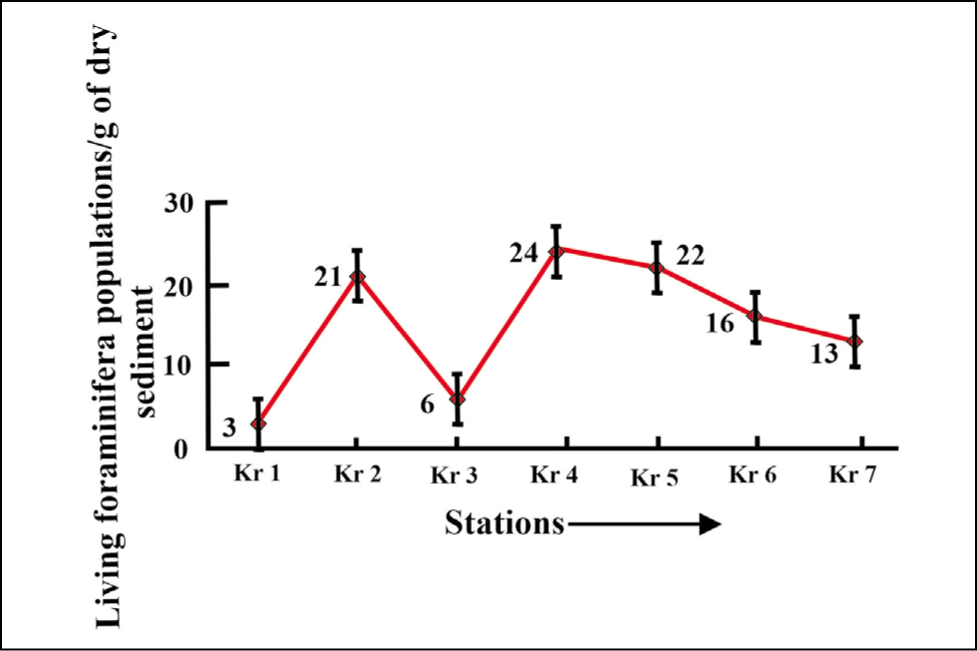
Living foraminifera populations are considerably lower throughout (Figure 13) (Kr 1 = 3 to Kr 4 = 24) the fjord.
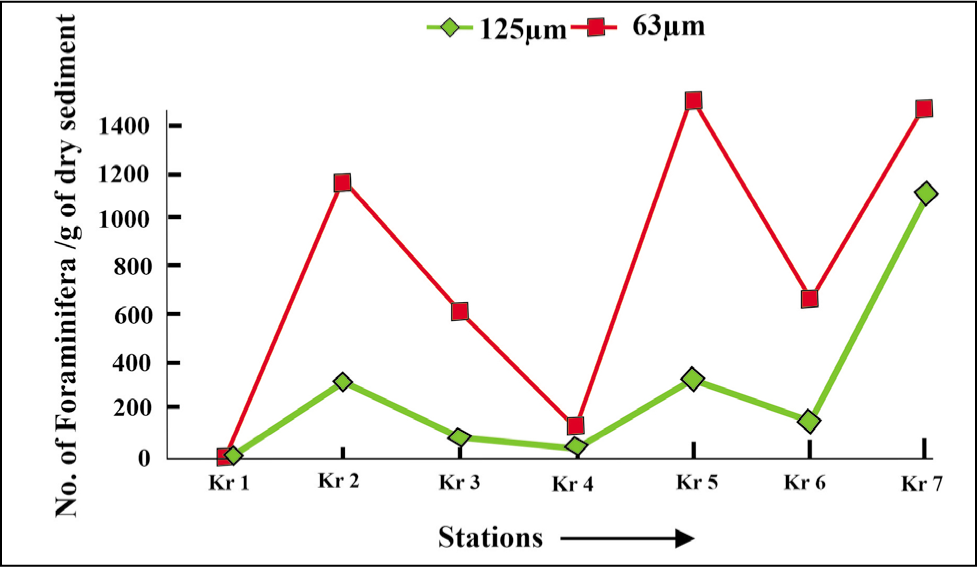
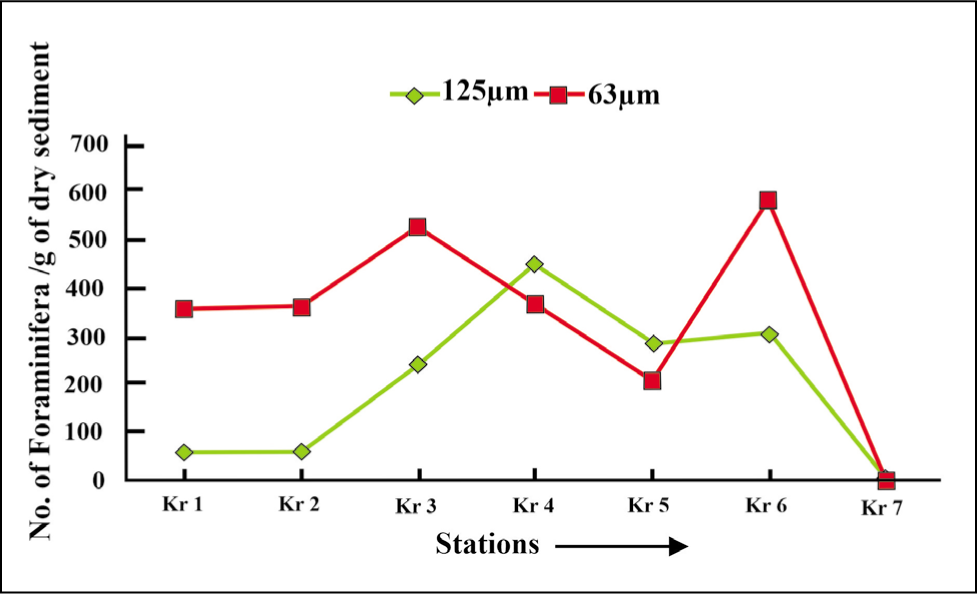
Based on the benthic foraminiferal size distribution (Figures 14 and 15), the smaller forms (>63 µm) are in good abundance as compared to the larger forms (>125 µm).
Living foraminifera plot for Krossfjorden.
Benthic foraminifera size variation in Krossfjorden (2016).
Benthic foraminifera size variation in Krossfjorden (2018).
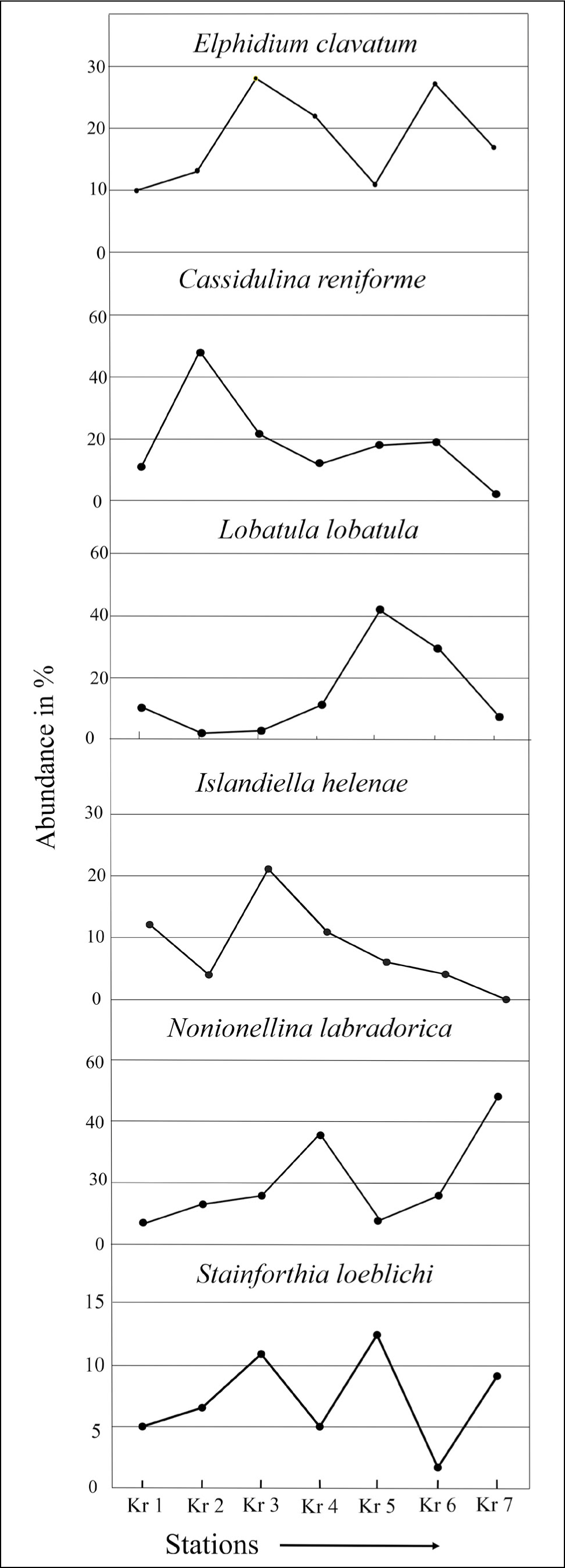
Within calcareous hyaline, Cassidulina reniforme (20%–45%) and Elphidium clavatum (27%–30%) are found to be the most dominant calcareous benthic foraminifera in the proximal part (Figure 16), Nonionellina labradorica (23%–48%) and Lobatula lobatula (15%–40%) occur significantly in the middle and distal parts (Kr 4, Kr 5, Kr 6 and Kr 7) (Figure 16), while Islandiella helenae (10%–20%) occurs in the proximal and middle (Kr 1, Kr 3 and Kr 4) parts of the fjord. Stainforthia loeblichi occurs (10%–12%) in the middle and distal parts. Among agglutinated species, Adercotryma glomeratum (10%) and Textularia spp. (12%) are the two most characteristic species in the proximal part, with a minor presence of Recurvoides sp. (1%) and Reophax sp. (1%).
Dominant benthic foraminifera abundance in Krossfjorden.
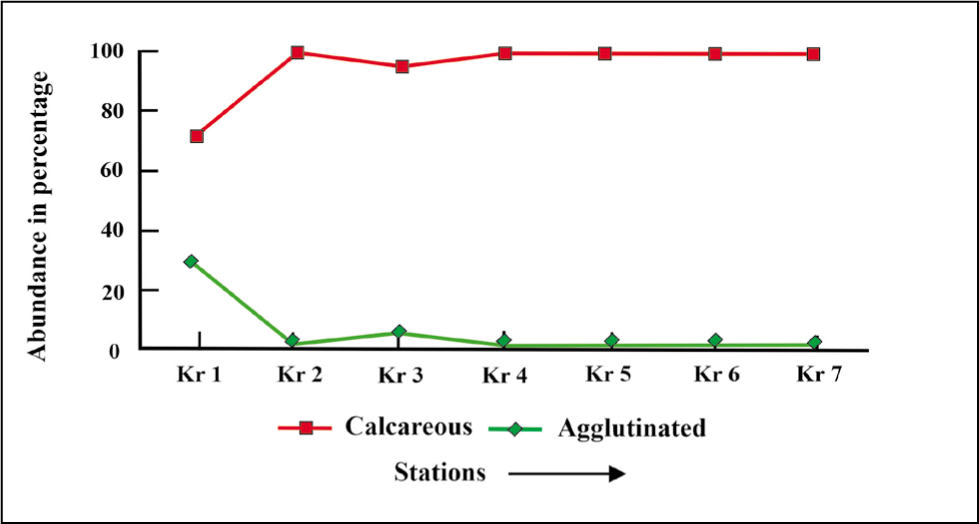
The calcareous forms are more abundant than the agglutinated ones in all the stations except at the extreme proximal station Kr 1, where they are less abundant compared to the agglutinated ones (Figure 17).
Calcareous and agglutinated foraminifera plot for Krossfjorden.
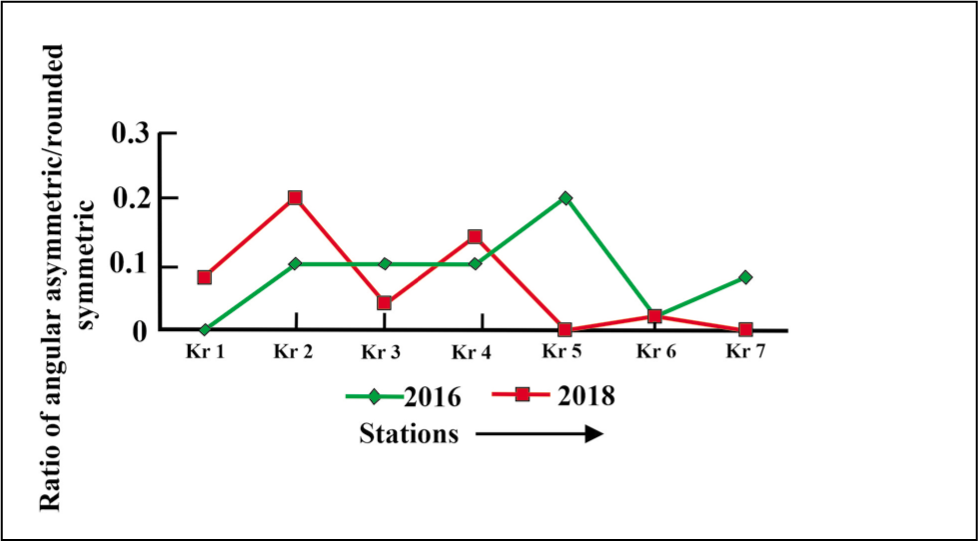
The fjord shows an increase in the ratio of angular asymmetric to rounded symmetric forms in the proximal and middle parts (0.1) and reduces to (0.03) in the distal part of the fjord (Figure 18).
Angular asymmetric to rounded symmetric plot for Krossfjorden.
The Fisher alpha diversity index of foraminifera in Krossfjorden varies from 2 to 5 from the proximal to middle and distal parts. The value indicates that foraminifera from the middle to distal fjord are highly diverse compared to the proximal part (Figure 19).
Fisher’s alpha diversity plot for Krossfjorden.
Discussion
Kongsfjorden
The present study represents foraminifera distribution in the Kongsfjorden area. Calcareous benthic foraminifera species that have been found in Kongsfjorden are Cassidulina reniforme, Elphidium clavatum, Lobatula lobatula, Islandiella helenae, Stainforthia loeblichi and Nonionellina labradorica. Accessory species are Melonis affinis, Haynesina orbicularis, Astrononion hamadaense, Buccella frigida, Globobulimina sp., Islandiella norcrossi, Quinqueloculina sp., Elphidium bartletti, Bolivinellina sp., Pyrgo sp., Dentalina sp., Uvigerina sp., Lagena sp., Fissurina sp., Robertina sp., Ammodiscus sp. and Trifarina fluens.
Agglutinated benthic foraminifera species that have been found in Kongsfjorden are Labrospira crassimargo, Adercotryma glomeratum, Textularia torquata, Textularia earlandi, Spiroplectammina biformis, Recurvoides turbinatus, Reophax sp., Portatrochammina bipolaris, Ammotium sp., Miliammina fusca and Eggerella sp.
The samples were collected twice in 2016 and 2018 and averaged for evaluation of foraminifera composition and environmental interpretation. The result shows that the total abundance of benthic foraminifera specimens increases from the glacial proximal through the glacial distal (Figure 3), indicating high surface water productivity in the distal glacial part of Kongsfjorden (Osterman et al., 1999). Foraminifera specimens >125 µm occur in more significant proportions towards the distal fjord (Figures 5 and 6), indicating the availability of nutrients, salinity and temperature sufficient for the growth of the foraminifera. The fjord is characterised by lesser-living foraminifera (Figure 4). The density of dead foraminifera has been attributed to either remobilisation and transportation by active currents from shallow to deeper settings or the dissolution of living foraminifera (Racine et al., 2018).
It has been found that the proximal part of the fjord is characterised by Elphidium clavatum and Cassidulina reniforme assemblage (Figure 7). In contrast, the middle and distal fjords are characterised by Nonionellina labradorica, Islandiella helenae and Lobatula lobatula assemblages. Nonionellina labradorica occupies significant proportions in the distal ocean sediment.
Glacial Proximal Part:
The present study shows a greater abundance of stress-tolerant species, that is, E. clavatum (23%–25%) and Cassidulina reniforme (36%–45%), in the glacier’s proximal part (stations KO 1, KO 2 and KO 3). This co-dominance of C. reniforme and E. clavatum typically characterises glacial proximal foraminifera assemblages in Svalbard fjords with low faunal diversity (Hansen & Knudsen 1995; Korsun et al. 1995). Spitsbergen fjord foraminifera assemblages dominated by C. reniforme have been linked to winter-cooled waters in the inner parts (Bergsten et al., 1998). The species prefer temperatures below 2 °C and stable salinity (>30 ppm) (Polyak et al., 2002). The present distribution of C. reniforme in this part of Kongsfjorden goes well with temperature (~3 °C), salinity (~30 ppm) and depths (51–86 m). The assemblage is typical in Late Pleistocene records of the Barents Sea and Norwegian shelf, indicating glacial proximity (Korsun et al., 1995). The occurrence of smaller foraminiferal tests (<63 µm) in the proximal part is high compared to larger tests (125 µm) (Figures 4 and 5). The reason may be due to meltwater discharge and high sedimentation rate, which affect primary production in the plankton community and thereby diminish foraminifera test sizes (Korsun et al., 1995). This result goes well with the study that has shown that the calving rate of the Kongsbreen glacier has increased in addition to increasing meltwater discharge during the melt season of 2016 (Schild et al., 2018). The abundance of agglutinated foraminifera in the proximal part has been attributed to meltwater flux and sediment load (Saraswat et al., 2018). Spiroplectammina biformis and Textularia spp. (23%) (Figure 7) are the two characteristic species that dominate the agglutinated forms in this part of the fjord. Textularia spp. and S. biformis have been identified in areas characterised by relatively cold, low-salinity ArW and often in glaciomarine sediments (Lloyd et al., 2006). This result matches well with cold (3 °C) and less saline (20–30 ppm) environments in the proximal part of Kongsfjorden. The increased ratio of overall angular asymmetric to rounded symmetric forms (0.8) and the presence of agglutinated forms (10%–20%) (Figures 8 and 9) have been attributed to the presence of cold water (>0 °C), low salinity, low organic carbon (Jennings & Helgadottir, 1994), rapid change in sea-ice cover (Rytter et al., 2002), increased sedimentation (Osterman et al., 1999) and oxygen deficiency (Saraswat et al., 2018) in the vicinity of glaciers.
Glacial Middle Part:
The middle part of the fjord characterises Nonionellina labradorica (31%–46%), Islandiella helenae (12%–14%) and Lobatula lobatula (45%–60%) assemblages. A significant amount of rounded calcareous foraminifera has been found in the middle (stations KO 4 and KO 5) parts of the Kongsfjorden. This high amount of calcareous, rounded benthic foraminifera in the middle indicates elevated oxygen concentration in the water, a low sedimentation rate, a greater distance from glaciers and an increase in organic carbon (Saraswat et al., 2018). The presence of rounded agglutinated foraminifera, that is, Adercotryma sp., only in the middle part relates to the presence of TAW (Hald & Korsun, 1997) at a greater distance from glaciers. This result indicates that Atlantic-sourced waters influence the middle part of Kongsfjorden (Lloyd et al., 2006). However, the increase in calcareous angular foraminifera species Stainforthia loeblichi in the middle part (5%–6%) indicates local oxygen-reduced conditions (Saraswat et al., 2018). Lobatula lobatula in coarse sediments (station KO 5) in the mid-fjord, close to the coast, indicates higher bottom currents in that region.
Glacial Distal Part:
Islandiella helenae in the distal part of the fjord (KO 6, KO 7 and KO 8) relates to a high stable salinity and summer ice-age productivity. At the same time, the presence of Nonionellina labradorica indicates increased food availability (Polyak et al., 2002). The high diversity and abundance of well-rounded calcareous foraminifera towards the distal part of the fjord represent the presence of well-oxygenated water (Saraswat et al., 2018). The wide occurrence of N. labradorica (Figure 7) in the sediment is ascribed due to the connection between the outer basin and shelf at a deeper level that causes a significant exchange of water masses and intrusion of AW in Kongsfjorden. This interaction enhances the availability of nutrients and food concentrations that helps to proliferate N. labradorica, which is not frequent in other fjords (Jernas et al., 2018). The result indicates elevated food concentration at least seasonally in the sediment in this part of Kongsfjorden (Polyak et al., 2002).
This preference to proliferate towards the distal glacier of N. labradorica significantly differs from its ubiquitous distribution in Kongsfjorden, as observed by Jernas et al. (2018). Based on the cumulative distribution of the most dominant benthic foraminifera, they concluded that from 2005 to 2008, the fjord was characterised by a single benthic foraminifera, N. labradorica, that occupies significant proportions and is strikingly different from other northern and southern fjords of Svalbard. In the context of morpho groups, the rounded calcareous symmetrical groups are more widespread in the distal Kongsfjorden than in the study by Saraswat et al. (2018), which showed calcareous rounded symmetrical forms are absent in the distal Kongsfjorden, while they are maximum at the confluence of two fjords and the inner part of the Kongsfjorden. In this study, angular agglutinated foraminifera, that is, Textularia spp., are dominant compared to the previous study by Jernas et al. (2018), where rounded agglutinated foraminifera, Adercotryma glomeratum, Recurvoides spp. and Labrospira crassimargo, have been reported as the dominant group. The present study shows rounded agglutinated foraminifera are considerable (4%–5%) in the middle and scarce (1%) in the distal part.
So, in summary, the environment of Kongsfjorden can be divided into three parts. The presence of agglutinated foraminifera and stress-tolerant species in this part indicates high turbidity, low salinity, meltwater runoff and sedimentation. The environment gradually changes to increase organic productivity and oxygen concentration in the middle part by accommodating increased concentrations of rounded calcareous. Towards the distal part, the environment experiences high surface primary productivity, nutrient and food availability, diatom concentrations (Jernas et al., 2018), well-oxygenated waters and high organic carbon, which is reflected by the presence of diverse (Figure 10) rounded calcareous specimens along with Nonionellina labradorica and the scarcity of agglutinated fauna.
Krossfjorden
Calcareous benthic foraminifera species that have been found in Krossfjorden are Cassidulina reniforme, Elphidium clavatum, Lobatula lobatula, Islandiella helenae, Stainforthia loeblichi and Nonionellina labradorica. Accessory species are Melonis affinis, Haynesina orbicularis, Astrononion hamadaense, Buccella frigida, Globobulimina sp., Islandiella norcrossi, Quinqueloculina sp., Elphidium bartletti, Bolivinellina sp., Pyrgo sp., Dentalina sp., Uvigerina sp., Lagena sp., Fissurina sp., Robertina sp., Ammodiscus sp. and Trifarina fluens.
Agglutinated benthic foraminifera species that have been found in Krossfjorden are Labrospira crassimargo, Adercotryma glomeratum, Textularia torquata, Textularia earlandi, Spiroplectammina biformis, Recurvoides turbinatus, Reophax sp., Portatrochammina bipolaris, Ammotium sp., Miliammina fusca and Eggerella sp.
The present study shows an overall high abundance of total foraminifera specimens of all test sizes (Figures 12, 14 and 15). This has made Krossfjorden distinct from other fjords in Svalbard. A low number of living foraminifera specimens has been observed.
It has been found that the proximal part of the fjord is greatly occupied by the Cassidulina reniforme and Elphidium clavatum assemblages; N. labradorica and Islandiella helenae occupy significant proportions in the proximal glacial sediment (Figure 16). The middle and distal fjords are characterised by Nonionellina labradorica, Lobatula lobatula and Stainforthia loeblichi assemblages.
Glacial Proximal Part::
Cassidulina reniforme occurs mainly at the proximal part (48%), in addition to E. clavatum (28%), N. labradorica (16%–25%) and Islandiella helenae (10%–20%) (Figure 16). One interesting observation of our study is the notable occurrence of N. labradorica and Islandiella helenae near glaciers (stations Kr 2 and Kr 3). Nonionellina labradorica is associated with temperate and saline TAW (Hald & Korsun, 1997). The assumption matches Adercotryma sp. (10%), which relates to Atlantic source waters (Hald & Korsun, 1997). Therefore, increased N. labradorica frequencies (stations Kr 2 and Kr 3) in the proximal part of Krossfjorden indicate nutrient enrichment in the site (Korsun & Hald, 1998). Islandiella helenae has been found in cold temperatures and a range of salinities from 33 to 34.5 ppm in polar waters and flourishes with pulse productivity at sea-ice margins (Cage et al., 2021). The stable salinity (32–34 ppm) and temperature (3 °C to 4 °C) condition of Krossfjorden may be the reason behind this abundance of foraminifera. Studies have shown that benthic fauna use macroalgal detritus as their food, which also constitutes a significant source of the Arctic fjord’s food web (Jernas et al., 2018). So, it can be assumed that AW influence on the fjord and the elevated concentration of microalgal detritus have made the Krossfjorden ecosystem more productive and suitable for the growth of N. labradorica and Islandiella helenae. Among the agglutinated taxa, Textularia sp. occurs only near glaciers (Kr 1). However, the deficiency of oxygen and low organic carbon in the proximal glacier environment cannot be ruled out, supported by the abundance of agglutinated foraminifera (Figure 17) in the proximal part and the enhanced ratio (0.2) of angular asymmetric by rounded symmetric (Figure 18).
Glacial Middle Part:
The co-occurrence of Cassidulina reniforme, Elphidium excavatum and Nonionellina labradorica (stations Kr 4 and Kr 5) indicates less glacial activity and a regular saline, cold environment. The co-occurrence of E. clavatum, C. reniforme, N. labradorica and I. helenae in this part of the fjord indicates the high organic carbon and phytoplankton density of Krossfjorden (Korsun & Hald, 1998). This assumption is consistent with the abundance of angular calcareous foraminifera Stainforthia loeblichi (12%) (Figure 16) in local oxygen-restricted with a high organic carbon zone in the middle part (Saraswat et al., 2018). This causes a high angular asymmetry to the rounded symmetric ratio observed in 2016 (Figure 18). However, a sudden decrease in the foraminiferal population is observed in the middle part, which denotes low food availability and depleted oxygen concentration. The notable presence of the distal glacial species Lobatula lobatula reflects a higher energy environment due to suspended organic carbon enrichment (Korsun & Hald, 1998).
Glacial Distal Part:
The presence of N. labradorica constitutes the bulk of the foraminiferal population at the distal sediment (Kr 6 and Kr 7), along with rounded, symmetric calcareous forms, that is, Islandiella helenae and Lobatula lobatula, indicates a well-productive, oxygenated and nutrient-rich environment in Krossfjorden.
The more significant amount of calcareous benthic foraminifera in the current study matches well with a notable preceding study by Hald and Korsun (1997) at Krossfjorden, which showed the presence of more than 75% of calcareous benthic foraminifera assemblage throughout the fjord. Nonionellina labradorica (36%–48%) and Elphidium clavatum (28%) in the present study resemble the previous one and differ in the presence of Cassidulina reniforme (22%–48%), followed by Islandiella helenae (21%) and Stainforthia loeblichi (12%). Stainforthia loeblichi has been found to occur in less than 10% of the previous study. Adercotryma glomeratum, Recurvoides sp. and Labrospira crassimargo were recorded in the distal part. The present study includes L. crassimargo and Recurvoides sp. less than 1%. The present study matches the dominance of calcareous fauna and rounded agglutinated forms.
In summary, the ecology and environment of Krossfjorden can be divided into three regions. The glacial proximal starts with a low organic carbon and oxygen-restricted zone comprising agglutinated foraminifera, a high ratio of angular asymmetric to rounded symmetric forms and a low count of total benthic foraminifera. It changes to a more productive, nutrient-enriched zone with the occurrence of N. labradorica and I. helenae. Then the environment experiences the local oxygen-depleted zone, characterised by a sudden decrease in TFN and angular asymmetric forms increase in abundance in the middle part. After that, a more productive, higher energy and elevated nutrient concentration zone at the distal part of the fjord reflected from abundant and diverse foraminifera (Figure 19).
Kongsfjorden and Krossfjorden
The two fjords, Kongsfjorden and Krossfjorden, reveal important perceptions towards foraminifera ecology and the fjord environment. Cassidulina reniforme and Elphidium clavatum assemblages in the proximal part characterise both fjords. Krossfjorden differs from Kongsfjorden by having N. labradorica and I. helenae in the proximal part. This difference indicates that meltwater flux and sedimentation rate are high in Kongsfjorden, resulting in a high frequency of agglutinated forms (10%–20%) in three samples (KO 1, KO 2 and KO 3) compared to Krossfjorden. On the contrary, the proximal part of Krossfjorden exhibits cold, stable salinity and less turbidity, and is more influenced by Atlantic source waters. The agglutinated forms have been found concentrated only at the innermost station, Kr 1 (30%). Krossfjorden shows a greater abundance of calcareous forms from the proximal to the distal part, while Kongsfjorden shows an abundance from the middle to the distal part. Nonionellina labradorica has been found to occur from the middle to the distal part of Kongsfjorden, while the species notably occurs in the proximal sediment of Krossfjorden. The oxygen minimum zone is pronounced in the middle part of Krossfjorden, as evidenced by the occurrence of Stainforthia loeblichi (10%–12%) compared to Kongsfjorden (5%–6%). Angular agglutinated forms, that is, Textularia sp., are common in Kongsfjorden compared to rounded agglutinated forms, that is, Adercotryma sp., that are abundant in Krossfjorden. Kongsfjorden shows an abundance of a larger size fraction of foraminifera specimens compared to Krossfjorden, which shows an abundance of foraminifera specimens of both size fractions. Both fjords reflect a well-oxygenated environment in the distal parts with primary surface productivity. The comparative study of Kongsfjorden and Krossfjorden has been summarized in (Table 3).
Conclusion
Studying the foraminiferal ecology of Kongsfjorden and Krossfjorden, it can be concluded that:
The proximal parts of Kongsfjorden and Krossfjorden constitute E. clavatum and C. reniforme assemblage. Abundant angular, asymmetric agglutinated forms characterise the area of Kongsfjorden, that is, Textularia sp. and Spiroplectammina biformis. In addition, Krossfjorden contains Adercotryma glomaratum. The assemblage indicates a low saline, increased sedimentation and meltwater runoff environment. The middle and distal parts constitute N. labradorica, L. lobatula and I. helenae assemblages. The agglutinated foraminifera mainly comprise Adercotryma glomaratum. The foraminiferal populations indicate well-oxygenated, low sedimentation rate, high surface primary productivity and influence of AW. Krossfjorden appears more enriched in productivity and phytodetritus than Kongsfjorden, with abundant N. labradorica in the proximal and distal parts, followed by I. helenae and a more rounded calcareous fauna. The presence of Stainforthia loeblichi in the middle part of the Krossfjorden shows the occurrence of low oxygen conditions.
Supplemental Material
Supplemental material for this article is available online.
Footnotes
Acknowledgements
The author would like to acknowledge the National Centre for Polar and Ocean Research, Goa, for field sample collection and other logistic support during the Indian Arctic Expedition in 2016 and 2018.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The author (DC) received funding from the UGC-SRF fellowship for this research work at Jadavpur University.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.