
Abstract
The Oligocene–Miocene Asmari Formation is a sequence of shallow-water carbonate carbonates in the Zagros Basin. This formation is a significant oil reservoir that was formed in the Zagros foreland basin in southwestern Iran. The Razan section, located in the northwest of Zagros in the Lorestan province, was investigated to study microfacies types and evaluate the sedimentary environment. This section was then compared with the MakhmalKuh and Dehloran sections. It comprises limestone units with thin to medium strata, sometimes thick-bedded limestone, and other skeletal and non-skeletal components. Based on the biostratigraphy studies, 24 foraminifer genera and species have been identified. Among the foraminifera, the species of Peneroplis farsensis, Peneroplis evolutus, Austrotrilina howchini, Austrotrilina asmarensis, Dendritinarangi, Quinqueloculina sp., Triloculina trigonula, Pyrgo sp., Polymorphinids, Spiroluculina sp. and Meandropsina anahensis are the most important specimens. Based on identified co-occurrence taxa, the Austrotrilina howchini–Peneroplis evolutus assemblage zone has been recognised in the study area, which indicates early Miocene (Aquitanian). Texture analysis and faunal assemblages led to the recognition of fourteen microfacies. Based on the petrographic analysis, five depositional settings were determined in the studied section: (a) the outer ramp settings dominated by pelagic foraminifera, bryozoan, echinoid and benthic foraminifera such as Nummulites, Rotalia, Dendritina and Meandropesina iranica as a proxy for relatively deep marine conditions; (b) the mid ramp, characterised by sizeable perforate foraminifera such as Amphistegina, Operculina, Heterostegina and planktonic foraminifera; (c) the barrier, marginal marine environment dominated by foraminifera such as Miogypsinoides sp., Peneroplis, Meandropsina, Dendritina and Neoalveolina, as well as peloid and corallinacean algae; (d) the lagoon dominated by benthic foraminifera Austrotrillina, Dendritina, Peneroplis, Miogypsinoides, Borelis, Archaias, miliolids and red algae and (e) the tidal flat, the shallowest part of the inner ramp, dominated by fenestral dolo-mudstone without any fossils. The faunal assemblages represent warm tropical waters under euphotic and mesophotic conditions in a homoclinal ramp.
Introduction
The Zagros Basin has a passive margin with convergent orogeny in the Mesozoic and Cenozoic (Aghanabati, 2004; Motiei, 1994). After the Laramide compression phase, which led to the closure of the Neo-Tethys Ocean, the Zagros Basin was developed in the northeastern Arabian Platform (Heydari, 2008). This basin extends from Turkey to southwestern Iran. Due to tectonic factors, the foreland basin was formed during the Paleogene Period in the Zagros (Lacombe et al., 2011). Afterwards, an intra-shelf basin was created throughout the Oligocene to early Miocene that includes shallow carbonate platform deposits belonging to the Asmari Formation (James & Wynd, 1965; Motiei, 1994; Sherkati & Letouzey, 2004). The intra-shelf basins were established during the Oligo-Miocene, surrounded by deep water basins of the Paleogene (van Buchem et al., 2010; Ziegler, 2001). Therefore, the Asmari Formation was first formed in the intra-shelf basins on the carbonate platform (Ziegler, 2001). Stratigraphically, the Asmari intra-shelf basin sediments formed above the Pabdeh Formation at the end of the Oligocene. These carbonate platforms are widely observed throughout the Zagros and worldwide in the Tertiary (Pomar et al., 2014). They have large hydrocarbon reservoirs, making them a critical hydrocarbon reservoir in the Zagros Basin (Shabafrooz et al., 2015). Iran’s Oligo–Miocene Asmari Formation has been deposited in provinces such as Fars, Khuzestan and Lorestan in the Zagros Basin (James & Wynd, 1965). At its type section, this formation consists of 314 m of limestones, dolomitic limestones and argillaceous limestones (Motiei, 1994), which the Gachsaran Formation overlies during the middle Miocene. In recent decades, extensive studies have mainly been conducted on the biostratigraphic criteria, determination of age, sedimentary and facies analyses of the Asmari Formation (e.g., Ehrenberg et al., 2007; Laursen et al., 2009; Mossadegh et al., 2009; Rahmani et al., 2009; Seyrafian et al., 2011; Shabafrooz et al., 2015; Taheri et al., 2017; van Buchem et al., 2010; Vaziri-Moghaddam et al., 2006, 2010). Among these, Laursen et al. (2009) and van Buchem et al. (2010) introduced new zonation based on faunal associations calibrated by strontium isotopes for the Asmari Formation. In the present study, the primary purpose is to evaluate biostratigraphic data based on strontium isotope stratigraphy by Laursen et al. (2009) and van Buchem et al. (2010), age determination of the Asmari Formation carbonates and reconstruction of the sedimentary environment.
General Geology
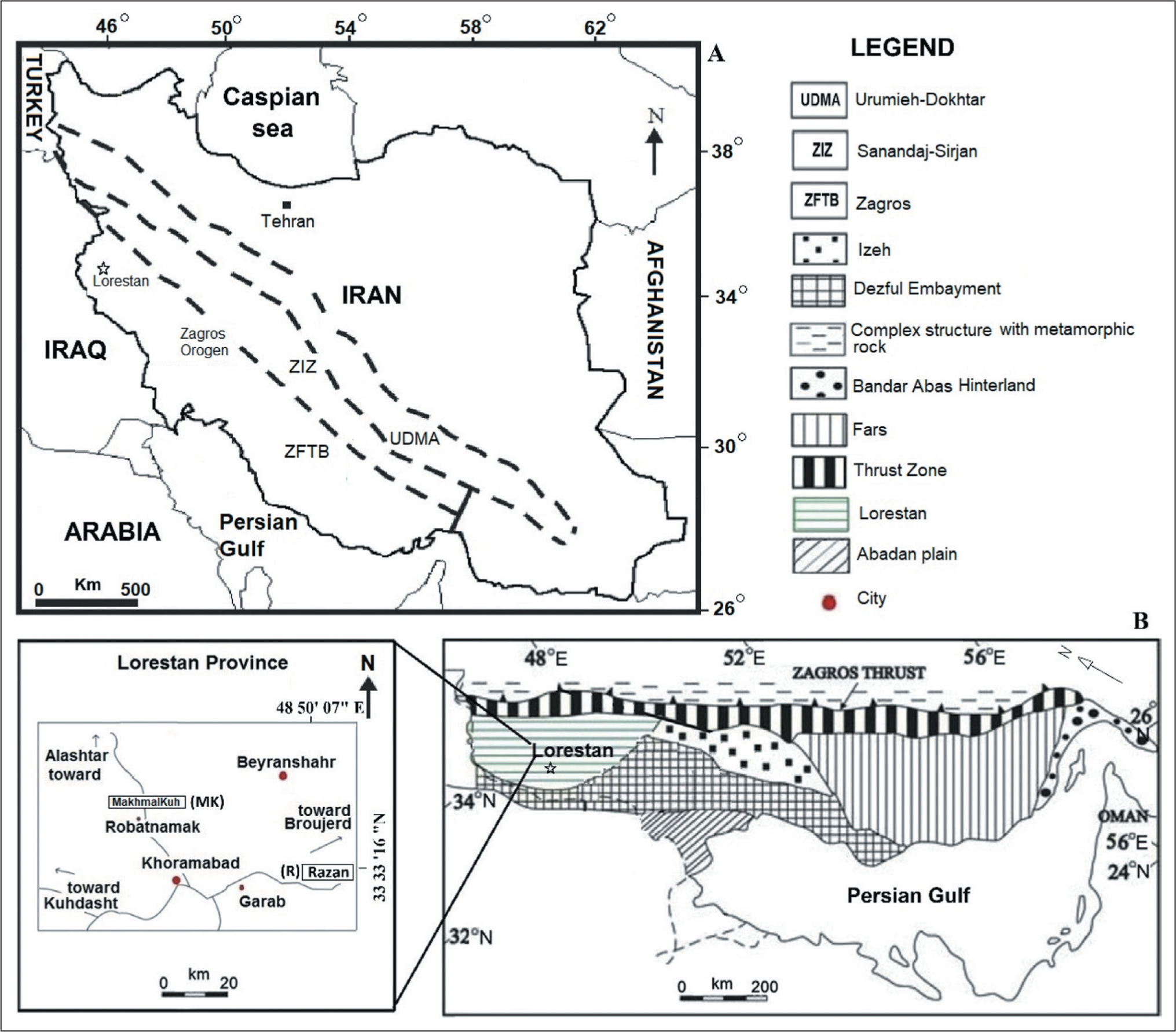
The Zagros Basin is located in southwestern Iran, which is divided into tectonostratigraphic zones, including the Zagros fold thrust (ZFT), the Zagros imbricated (ZI) and the Urumieh–Dokhtar magmatic (UDM) (Alavi, 2004; Stocklin, 1968). The ZFT zone includes Fars province, Khuzestan province (including the Dezful Embayment) and Lorestan province (Motiei, 1995; Sherkati & Letouzey, 2004). The studied sections are located in the ZFT zone, in Lorestan province. The Razan section is situated about 55 km north-east of Khorram Abad in northwestern Iran, in geographical coordinates N: 33 33′ 16″, E: 48 50′ 07″ (Figure 1). The MakhmalKuh section, with geographical coordinates N: 33 36′ 9″ and E: 48 17′ 22″, is located about 15 km northwest of Khorram Abad. The Dehluran section, with coordinates N: 32° 51′ 20″, E: 43° 54′ 3″, is located west of Khorram Abad (Vaziri-Moghaddam et al., 2010). These were compared with the global standard zonations of Wynd (1965), Cahuzac and Poignant (1997) and Laursen et al. (2009).
(A) General Map of Iran and Tectonostratigraphic Units in Southwestern Iran. Abbreviations of the Zagros Basin: UDMA, Urumieh-Dokhtar Magmatic Arc; ZFTB, Zagros Fold-Thrust Belt; ZIZ, Zagros Imbricate Zone (after Alavi, 2004). (B) Geological Map of the Zagros Fold-Thrust Belt with its Structural Provinces (Modified after Falcon, 1961; Sherkati and Letouzey, 2004). The Lower Left Corner Rectangle Shows the Location of the Studied Area and Sections. Abbreviations of the Zagros Basin: UDMA: Urumieh–Dokhtar magmatic arc; ZFTB: Zagros fold–thrust belt; ZIZ: Zagros imbricate zone (After Alavi, 2004).
Materials and Methods
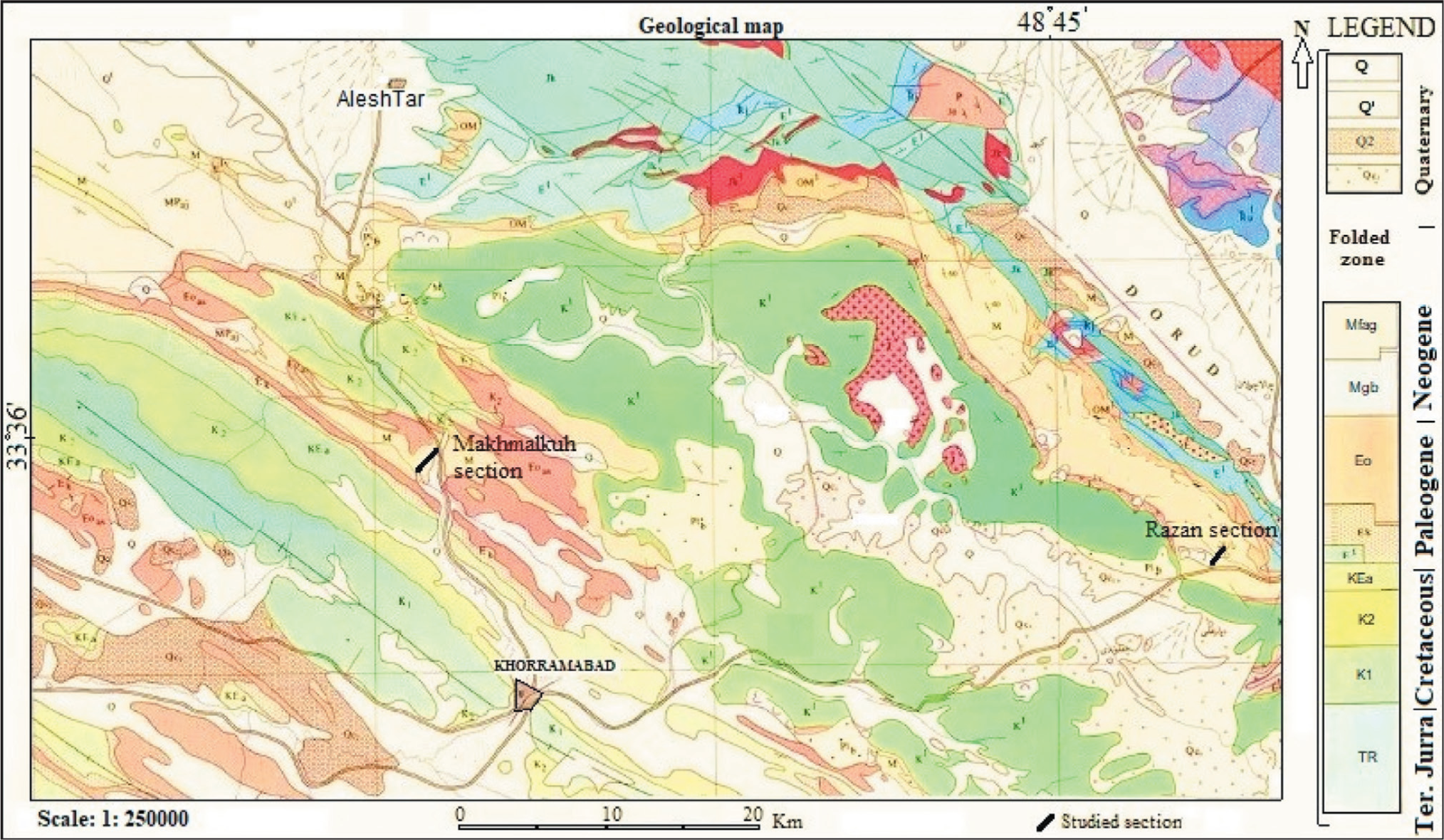
This study was analysed using two stratigraphic sections of the Asmari Formation in Lorestan province. These are Razan (R) in the northeast trend and MakhmalKuh (Mk) in the northwest near Khorram Abad City, located in central Lorestan, northwest Iran. Another outcrop (Dehluran section), measured by Vaziri-Moghaddam et al. (2010), has also been used better to understand the depositional environment and correlation in this basin. Systematic sampling was performed with one sample per 1 m for approximately 85 m of the MakhmalKuh section and 110 m from the Razan section (every 1–2.5 m) with 70 samples. A total of 155 thin sections were examined and studied with an optical microscope to analyse semi-quantitative microfacies and distribution of faunal assemblages (Figure 2). The terminology is used to describe the facies’ texture following the classification schemes of Dunham (1962) and Embry and Klovan (1971). Many researchers’ concepts have been used for biostratigraphic description (e.g., Amiri Bakhtiar et al., 2011; Bolli, 1966; Caron, 1985; Kalantari, 1986; Loeblich & Tappan, 1988; Postuma, 1971; Premoli Silva & Verga, 2004). Therefore, the microfacies characterisation was done based on fossil content, sediment texture, grain composition and grain size.
Geological Map of the Studied Sections and Chronostratigraphic Units in the Lorestan Province. The Rectangle shows the Location of the Studied Area and Sections (After Fakhari, 1985).
Results and Discussion
Lithostratigraphy
The studied Asmari Formation, 110 m in thickness in the Razan section, consists of limestone units with thin (ca. 1 cm) to thick (ca. 1 m) strata, which sometimes are dominated by benthic foraminifera and coralline red algae. Lithologically, the rock units have wackestone, packstone, grainstone and rare mudstone textures. The Asmari Formation outcropping in the MakhmalKuh section, 85 m in thickness, consists of thick (ca. 1 m) to medium (ca. 30 cm) limestone with thin interbedded limestone and dolomitic limestone, which is dominated by benthic foraminifera and coralline red algae. In both sections, the Asmari Formation overlies the dolomite of the Shahbazan Formation and is overlain by the anhydrite and gypsum of the Gachsaran Formation.
Biostratigraphy
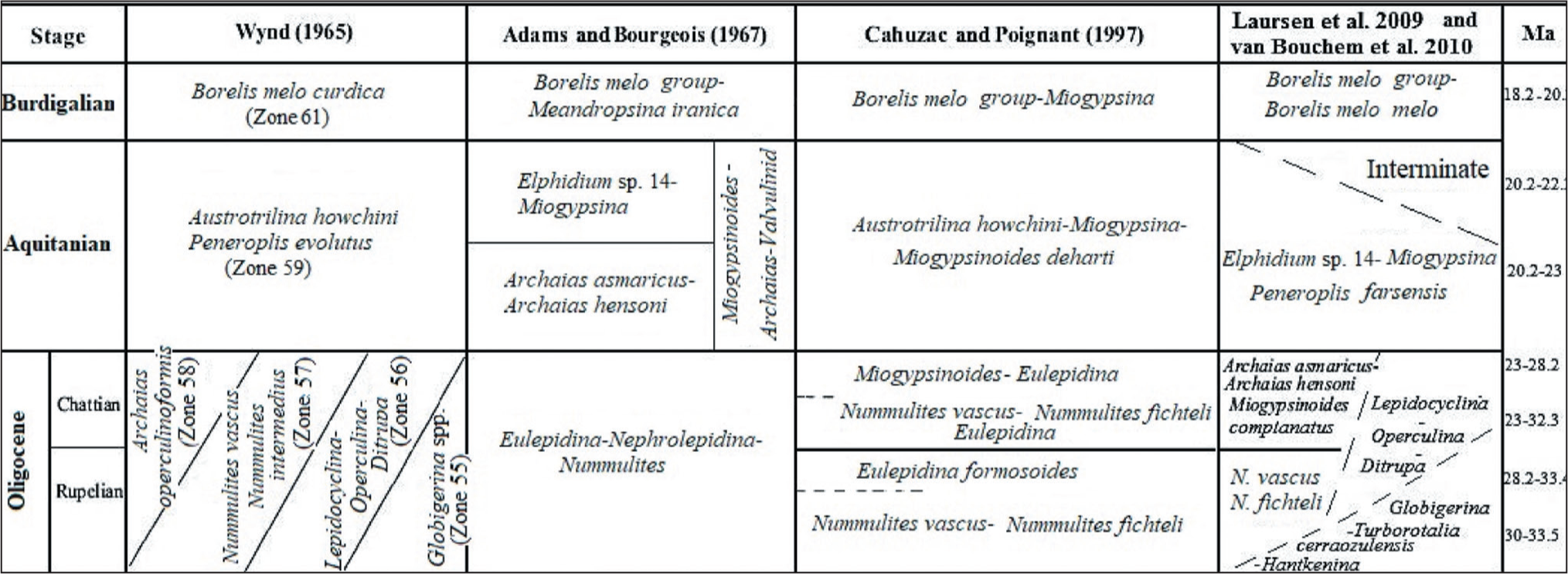
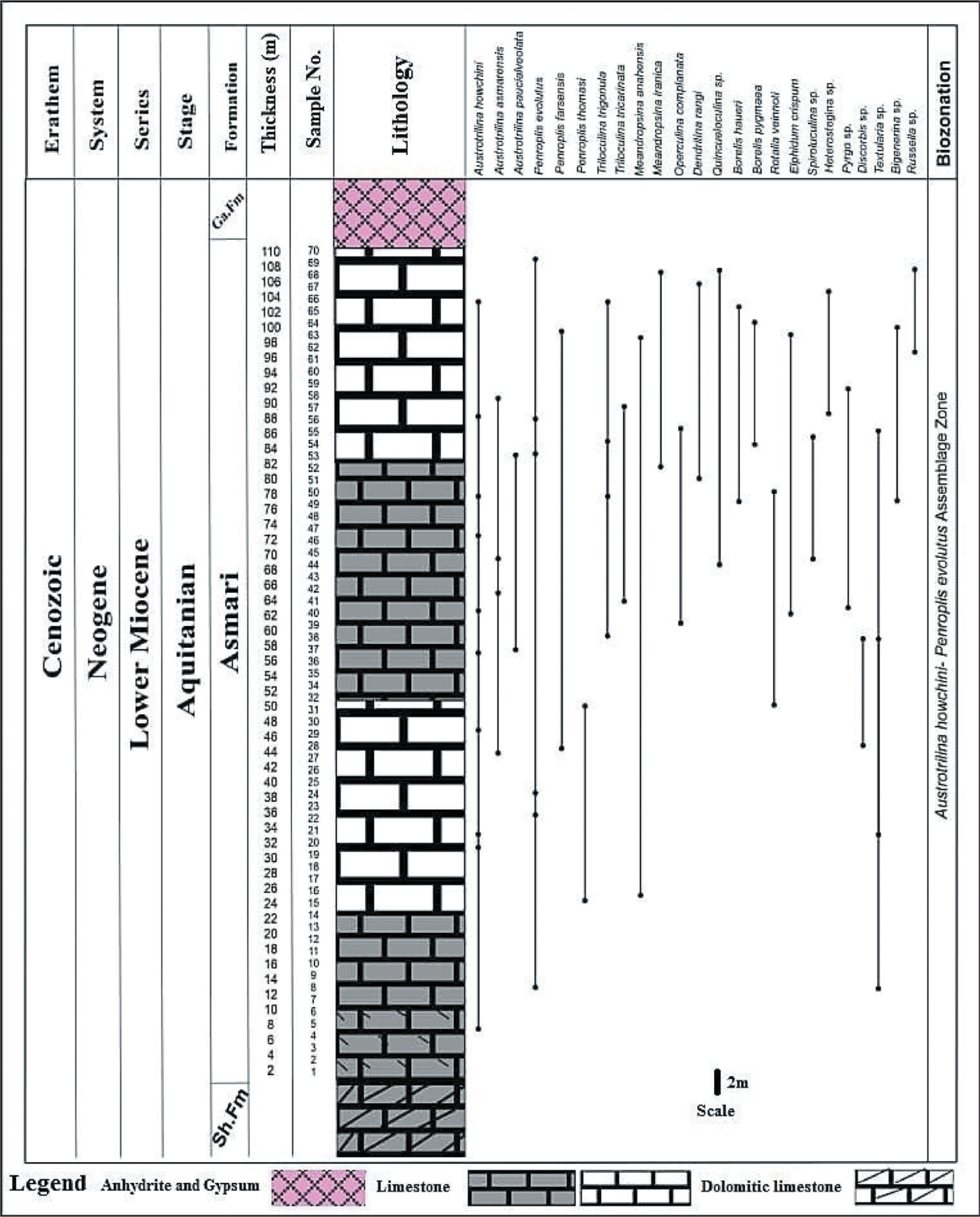
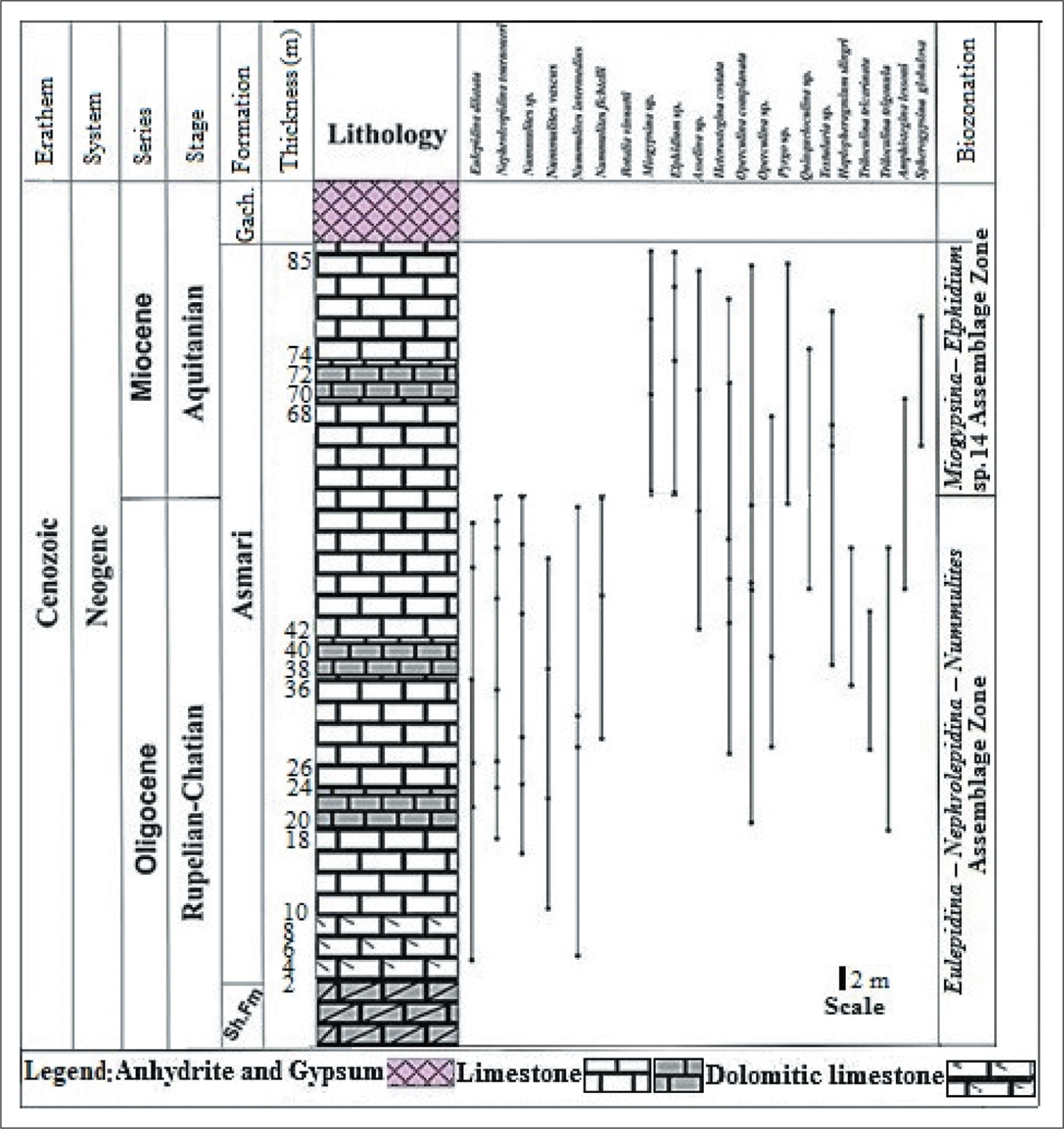
The carbonate platforms are characterised by active producers such as small and large benthic faunal assemblages during the Oligocene to Miocene transition (Hallock et al., 2006; Prothero, 2003). The biostratigraphic framework of carbonate platforms, such as deposits of the Asmari Formation in the Zagros Basin, was first studied by Wynd (1965) using foraminiferal index markers. He introduced biozones 55–61 for the Asmari Formation, which include Zone 55 (Globigerina spp./Oligocene), Zone 56 (Lepidocyclina–Operculina–Ditrupa/Oligocene), Zone 57 (Nummulites intermedius–Nummulites vascus/Oligocene), Zone 58 (Archaias operculiniformis/-Oligocene), Zone 59 (Austrotrillina howchini–Peneroplis evolutus/Aquitanian) and Zone 61 (Borelis melocurdica/Burdigalian). Subsequ-ently, Adams and Bourgeois (1967) modified previous studies and divided the Asmari Formation into three biozones, including the Eulepidina–Nephrolepidina–Nummulites assemblage zone for Oligocene, the Miogypsinoides–Archaias–Valvulinid assemblage zone, along with two sub-zones (Archaias asmaricus–Archaias hensoni and Elphidium sp. 14–Miogypsina) for Aquitanian (early Miocene) and the Borelis melo group–Meandropsina iranica assemblage zone for the Burdigalian (early Miocene). Afterward, Ehrenberg et al. (2007) introduced five biozones based on the distribution of Nummulites and Spiroclypeus blankenhorni, genera of Miogypsina and Archaias, as well as species of Borelis melo curdica. In the meantime, they suggested that the extinction of the Nummulites was in the late Rupelian. Then, Laursen et al. (2009) and van Buchem et al. (2010) proposed a new biozonation by the Sr isotope for the Asmari Formation (Figure 3). Strontium isotope stratigraphy is an established technique that provides the absolute age of past marine fauna. In the present study, in order to determine the age of the Asmari Formation, a comparison was made with the results of Sr analysis. In this regard, a total of 21 genera and species of foraminifera were identified in the MakhmalKuh section and 24 species in the Razan section, whose distribution has been drawn (Figures 4 and 5). Some selected benthic foraminifera are illustrated in the studied sections (Plates 1–3). Therefore, the biostratigraphic framework for the Asmari Formation is based on using the biozones introducted by Laursen et al. (2009) and van Bouchem et al. (2010), which are then compared with other zonations applied by Wynd (1965), Adams and Bourgeois (1967) and Cahuzac and Poignant (1997). In this study, three assemblage zones, one in the Razan section and two zones in the MakhmalKuh, have been recognised based on the stratigraphic distribution of benthic foraminifera. Then the stratigraphic correlation of the Asmari Formation is presented in the studied sections. The assemblage zones in Razan and MakhmalKuh sections are as follows:
Biozonation of the Oligo-Miocene Asmari Formation in the Zagros Basin (after Wynd, 1965; Adams and Bourgeois, 1967; Laursen et al., 2009; van Buchem et al., 2010) and for the European Basin (After Cahuzac and Poignant, 1997).
Vertical Biotic Assemblage Distribution of Asmari Formation at the Razan Section, Lorestan Province.
Distribution of Vertical Biotic Assemblage of the Asmari Formation in the MakhmalKuh Section, Lorestan Province.
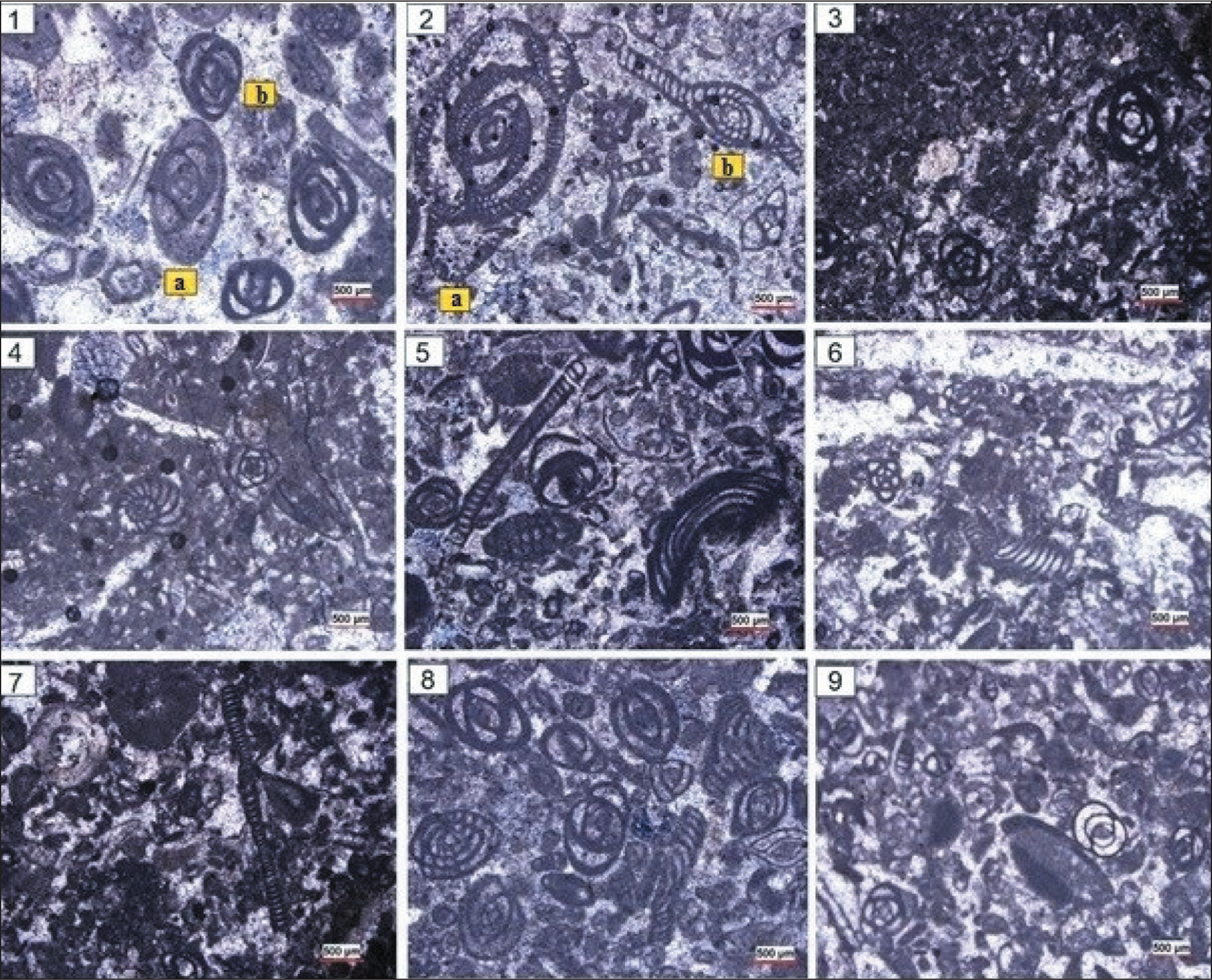
Some of the Microfossils Identified in the Razan Section. 1a) Austrotrilina Asmarensis; 1b) Triloculina Trigonula; 2a) Austrotrilina Howchini; 2b) Peneroplis Evolutus; 3. Austrotrilina Asmarensis; 4. Dendritina Rangi; 5. Meandropsina Anahensis; 6. Peneroplis Evolutus; 7. Peneroplis Evolutus; 8. Peneroplis Farsensis; 9. Polymorphinids. Scale is 500 µm.
Some of Microfossils that Identified in the Razan Section. 1. Pyrgo sp.; 2. Quinqueloculina sp.; 3. Spiroluculina sp.; 4. Triloculina trigonula; 5. Tubocellaria sp.; 6. Red Algal.
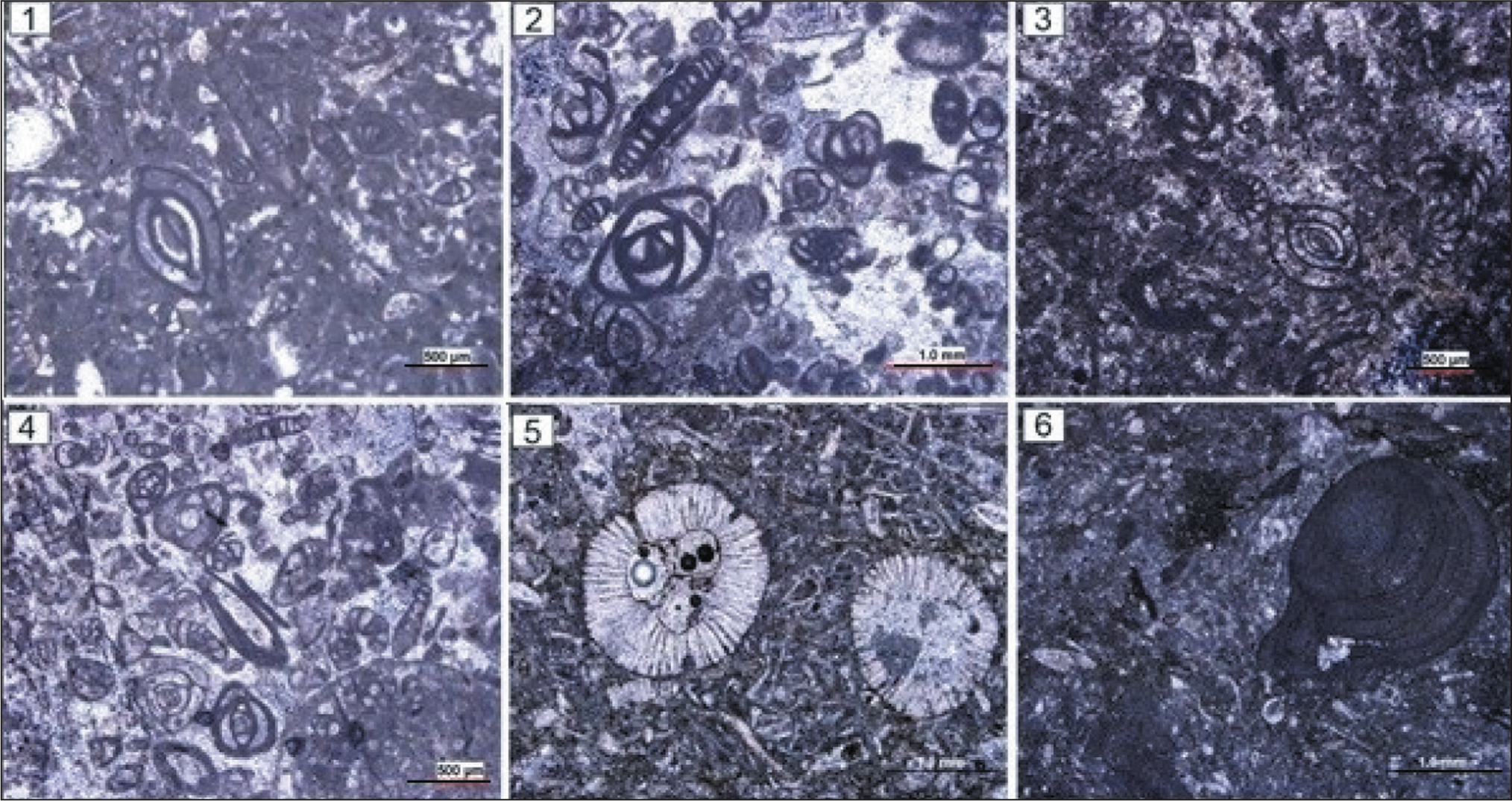
Some of Microfossils are Shown in the Studied Section. a. Nummulites cf. Vascus (Sample No. 2); b. Nummulites Intermedius (Sample No. 3); c. Nummulites cf. Fichtelli (Sample No. 3); d. Nummulites sp. (Sample No. 28); e. Amphistegina sp. (Sample No. 29);f. Amphistegina sp. (Sample No. 43); g. Asselina sp. (Sample No. 3); h. Nummulites sp. (Sample No. 51); i. Elphidium sp. (Sample No. 51); j. Eulepidina sp. (Sample No. 8); k. Nephrolepidina sp. (Sample No. 8); l. Heterestegina sp. (Sample No. 3); m. Operculina sp. (Sample No. 49); n. Pyrgo sp. (Sample No. 35); o. Quinqueloculina sp. (Sample No. 43); p. Quinqueloculina sp. (Sample No. 33); q. Textularia sp. (Sample No. 47); r. Miogypsina sp. (Sample No. 9); s. Miogypsina sp. (Sample No. 8); t. Nummulites sp. (Sample No. 51); u. Polymorphinids sp. (Sample No. 33).
Razan Section
A total of 17 genera and 24 species of foraminifera were identified in the Razan section (Plates 1 and 2). This led to the identification of the following assemblage zone:
Austrotrilina Howchini–Peneroplis Evolutus Assemblage Zone:
This zone is recorded in the Asmari Formation up to 110 m thick (Figure 4). The most important species in the studied section include: Peneroplis farsensis, Peneroplis evolutus, Austrotrilina howchini, Austrotrilina asmarensis, Operculina complanata, Dendritina rangi, Heterostegina sp., Quinqueloculina sp., Triloculina trigonula, Pyrgo sp., Spiroluculina sp., Meandropsina anahensis and Polymorphinids. This biozone is equivalent to the middle Asmari as defined by Thomas (1948). In fact, in this study, due to the absence of index fossils belonging to the lower and upper parts of the Asmari Formation (such as the genus Nummulites, Nephrolepidina, Eulepidina, Meandropsina iranica and Borelis melo curdica), it can be concluded that these parts of the Asmari Formation have not been deposited. Also, the mentioned faunal assemblage is according to biozone No. 59 Wynd (1965) in the Aquitanian age. Compared to the standard biozone of Laursen et al. (2009), due to the presence of Peneroplis farsensis, this microfauna is somewhat compatible with biozone No. 5 (Miogypsina–Elphidium sp. 14–Peneroplis farsensis), which indicates the age of Aquitanian. Also, due to the presence of Austrotrilina howchini, this assemblage zone can be consistent with the SB24 biozone in the Mediterranean region, as introduced by Cahuzac and Poignant (1997).
MakhmalKuh Section
A total of 21 genera and species were identified in the MakhmalKuh section (Plate 3). This led to the identification of the following two assemblage zones:
Eulepidina–Nephrolepidina–Nummulites Assemblage Zone:
This biozone is the first assemblage zone in the lower part of the Asmari Formation deposits in this section, which extends to a thickness of 58 m. The most important foraminiferal taxa are Nummulites vascus, Nummulites fichteli, Nummulites intermedius, Eulepidina elephantina, Eulepidina dilatata, Nephrolepidina tournoueri, Nummulites sp., Triloculina tricarinata, Operculina complanata, Operculina sp., Triloculina trigonula, Pyrgo sp., Amphistegina lessoni, Amphistegina sp. and Asselina sp. A species such as Eulepidina dilatata is distinguished from Eulepidina elephantine due to its large size and the absence of pillars that penetrate the shell surface. Both were identified in the studied sections in Rupelian and Chattian. In fact, the presence of Eulepidina indicates the age of the Oligocene. Adams and Bourgeois (1967), as well as Laursen et al. (2009), reported both Eulepidina dilatata and Eulepidina elephantine species (from genus Lepidocyclina) from the Asmari Formation. In this zone, among benthic foraminifers, intact specimens such as Amphistegina, Nephrolepidina and Nummulites are the most important components of the mesophotic zone (Pomar et al., 2014). In addition, large foraminifera are the most important components of the Oligocene–early Miocene transition. They divided the biostratigraphic sequence chart. Nephrolepidina tournoueri and Operculina complanata have also been recorded in the Zagros Basin since the late Rupelian and Chattian from this Formation (Laursen et al., 2009). Operculina complanata was reported from northeastern Italy in the late Oligocene from the Chattian deposits (Bassi et al., 2007). As mentioned, Nummulites vascus and Nummulites fichteli were recorded in this assemblage zone. The appearance of both species indicates late Rupelian and early Chattian ages, which is in accordance with the SB22 biozone in the Mediterranean basin from Cahuzac and Poignant (1997-). The species Nummulites vascus and Nummulites fichteli were reported from the deposits of Rupelian and Chattian in northeastern Italy, as well as the Zagros Basin located in southwestern Iran, by Bassi et al. (2007) and Laursen et al. (2009). The last occurrence of the Nummulites (namely N. bouillei) was recorded in the Mediterranean basin in the late Oligocene, late Chattian (SB23 zone) (Cahuzac & Poignant, 1997), but was not recorded in this study. Based on the opinion of Ehrenberg et al. (2007), the last appearance of the Nummulites occurred in late Rupelian. BouDagher-Fadel (2008) also believes that the extinction of Nummulites is related to the Rupelian. Moreover, the presence of Nummulites, Eulepidina together with Nephrolepidina, indicates Rupelian–Chattian intervals in many regions of Iran (Daneshian & Ramezani, 2007; Rahaghi, 1980). This zone is introduced by Adams and Bourgeois (1967) with Eulepidina–Nephrolepidina–Nummulites and is attributed to the Rupelian–Chattian interval. In addition, this zone is in accordance with Laursen et al. (2009) (= Nummulites vascus–Nummulites fichteli), which is Rupelian in age (Figure 3).
Miogypsina–Elphidium sp. 14 Assemblage Zone:
The second zone is recorded in the upper part of the Asmari Formation and is 27 m thick. The most important species in this part include: Heterostegina costata, Amphistegina lessoni, Operculina complanata, Spherogypsina globulosa, Miogypsina sp., Operculina sp., Elphidium sp., Asselina sp., Pyrgo sp., Quinqueloculina sp. and Planorbolina spp. This zone is in accordance with the zone of Adams and Bourgeois (1967) (Miogypsina–Elphidium sp. 14 assemblage zone), which is Aquitanian in age. They subdivided Elphidium based on the wall into Elphidium sp. 1 and Elphidium sp. 14. According to them, Elphidium sp. 14, with a thick wall, has a significant stratigraphic value that occurs in the Aquitanian. Hence, given the presence of Elphidium sp. 14, this zone coincides with the Aquitanian. Also, compared to the standard biozone of Laursen et al. (2009) (= Miogypsina–Elphidium sp. 14, Peneroplis farsensis assemblage zone), this faunal assemblage corresponds to the Aquitanian due to the presence of Elphidium sp. and Miogypsina. Genus Miogypsina has been reported in many parts of Iran, such as Fars, Khuzestan and Izeh, in the late Chattian-Aquitanian interval (Amirshahkarami et al., 2007; Sadeghi et al., 2009; Maghfouri Moghaddam et al., 2019). This species is important for biostratigraphic studies because it has been considered an indicator of the Aquitanian stage (Adams et al., 1983). This assemblage zone is in accordance with the middle Asmari Formation introduced by Thomas (1948). On the other hand, Operculina complanata and Nephrolepidina indicate late Chattian, which correlates with SB23 (Cahuzac & Poignant, 1997). Also, the faunal assemblage is consistent with the SB24 biozone in the Mediterranean region (Cahuzac & Poignant, 1997). Therefore, given the location of this zone, which is above the previous zone, the age interval of this faunal assemblage can be attributed to Aquitanian. Hence, due to the distribution of identified fossils in both assemblage zones, the age of the Asmari Formation in the studied section from Rupelian to Aquitanian is suggested (Figures 3 and 5).
Microfacies Analysis
The semi-quantitative analysis of microscopic observations or petrographic studies (e.g., texture, allochems and skeletal components in thin sections) led to the recognition of several microfacies in the studied sections. Here, fossils have been analysed based on some aspects of biostratigraphy. The facies reflect different positions in sedimentary environments such as lagoon (inner ramp) and open marine (mid ramp–outer ramp) that we will discuss.
Microfacies Description and Interpretation in the Razan Section:
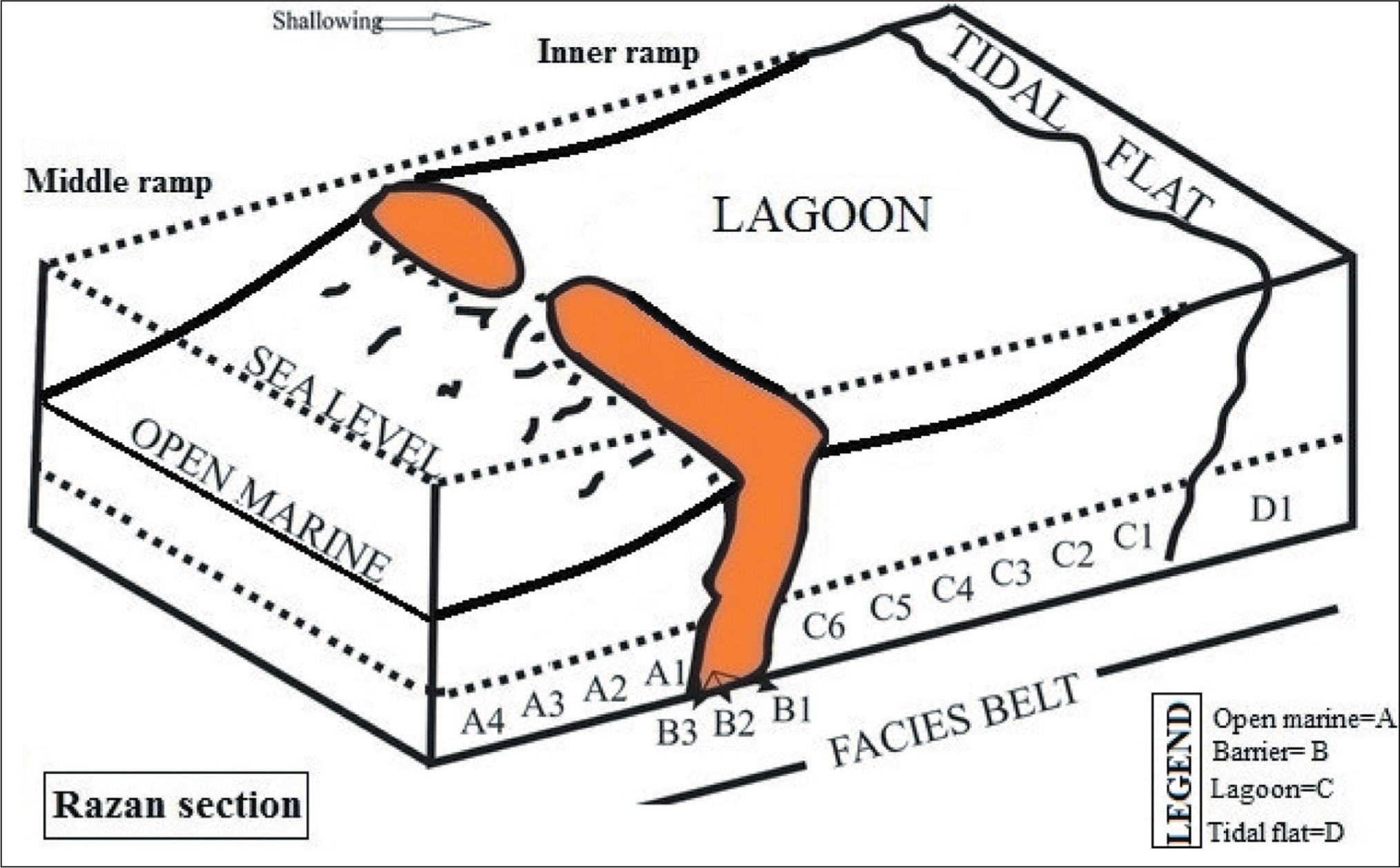
The Asmari Formation at the Razan section is subdivided into 14 different microfacies, each characterised by a petrographic analysis and foraminiferal assemblage (Figure 6). Based on the sedimentological analysis, four facies belts can be recognised: tidal flat/supratidal (D), lagoon (C), barrier (B) and open marine (A). The identified facies arranged by their depositional setting (deep to shallow) at the Razan section as follows:
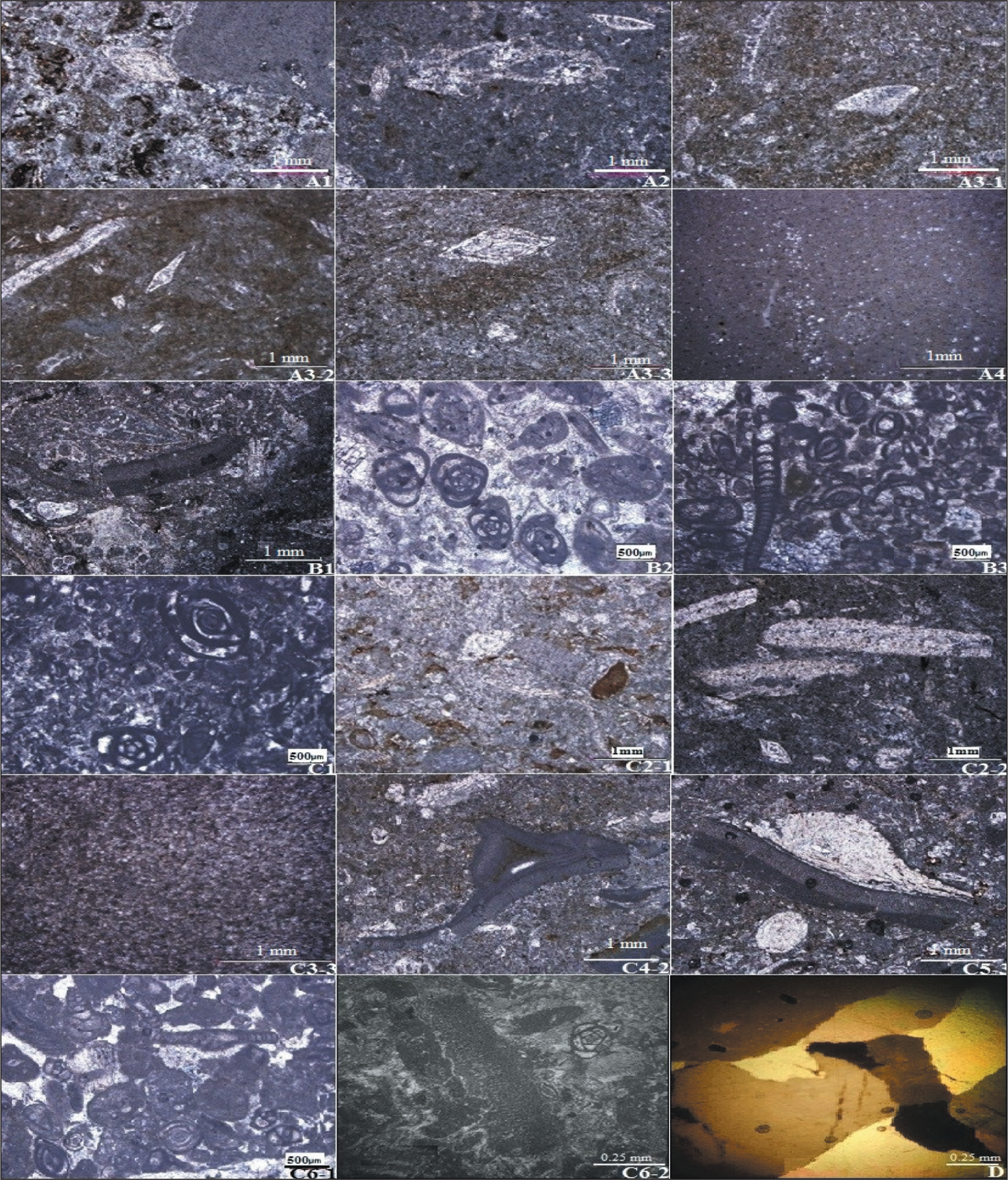
Microfacies of the Asmari Formation in the Razan Section; A1-1 (Foraminifer Bioclast Packstone); A2 (Foraminifer Algal Echinoderm Wackestone–Packstone); A3-1 (Bioturbated Echinoderm Algal Wackestone); A3-2 (Sponge Spicule Foraminifer Pelecypod Echinoderm Wackestone); A3-3 (Pelecypod Foraminifer Echinoderm Wackestone); A4 (Echinoderm Lime Mudstone); B1 (Intraclast Algal Foraminifer Grainstone); B2 (Intraclast Peloid Foraminifer Grainstone); B3 (Peloid Foraminifer Grainstone); C1 (Peloid Wackestone–Packstone); C2-1 (Bioturbated Intraclast Bioclast Peloid Wackestone); C2-2 (Bioturbated Foraminifer Gastropod Pelecypod Wackestone); C3-3 (Bioturbated Bioclast Lime Mudstone); C4-2 (Algal Foraminifer Wackestone); C5-3 (Algal Foraminifer Packstone); C6-1 (Sandy Echinoderm Foraminfer Packstone–Grainstone); C6-2 (Coral Pelecypod Echinoderm Foraminifer Packstone–Grainstone); D (Fenestral Dolomudstone with Evaporate Cast).
(A) Open marine microfacies. The microfacies, mainly characterised by the pelagic and benthic foraminifera, are also composed of other skeletal components such as brachiopods, echinoderms, bivalves, algae and miliolids, which are formed in a packstone, grainstone, mudstone and wackestone textures. Due to the fossil content of these facies, their sedimentary environment is far from shore, with low energy (Beavington-Penney & Racey, 2004). Facies analysis allows the identification of several sub-facies as follows:
(A4) Echinoderm lime mudstone. The microfacies is related to relatively deep open marine with a small amount of echinoderm, which suggests distal middle-ramp settings. Also, a small amount of large foraminifera indicates an oligotrophic environment (O’Connell et al., 2012). These microfacies are located in a greater amount of micritic matrix and have diagenetic processes such as vuggy porosity and hematitic phenomena.
(A3) Echinoderm foraminifer wackestone. The microfacies consists of three sub-facies:
(A3-1) Bioturbated echinoderm algal wackestone. The bulk of this facies consists of algae and its fragments (20%), bivalves and echinoderms (10%), a smaller amount of ostracods and bryozoan, properties of shallow open marine facies and moderate energy. The micritic matrix and the bioturbation processes are other characterised of these facies—the presence of these components suggests proximal middle-ramp settings.
(A3-2) Sponge spicule foraminifer pelecypod echinoderm wackestone. The facies were characterised by a fine-grained micritic matrix along with plagic foraminifera (10%), bivalves (17%) and echinoderms spines (20%). Other bioclasts are Nummulites, Rotalia, Dendritina, ostracods, bryozoans and miliolids. Large and flat tests of perforate foraminifera are the dominant microfauna of the intermediate and middle ramps because they were the best-adopted fauna to the environmental conditions such as low energy, average salinity, oligotrophic and lower limit of the photic zone (Romero et al., 2002).
(A3-3) Pelecypod foraminifer echinoderm wackestone. The facies were dominated by echinoderms and their fragments (15%). Moreover, these facies include perforate large foraminifera (10%) and bivalves (7%). Other bioclasts are foraminifera such as Meandropesina iranica, Rotalia, Dendritina and Neoalveolina (Borelis). The matrix is micritic, in which the diagenetic phenomenon of neomorphism is observed. The presence of planktonic foraminifera accompanied by large perforate foraminifera (Nummulitidae and Lepidocyclinidae) indicated a distal middle-ramp sedimentary setting between the normal wave base and the storm wave base in the lower limit of the photic zone (Corda & Brandano, 2003).
(A2) Foraminifer algal echinoderm wackestone–packstone. This facies is dominated by echinoderms and their fragments (25%). In addition, the facies include ostracods, algal pieces (15%) and a small number of perforate benthic foraminifera. The matrix is a fine-grained micrite. Diagenetic processes in this microfacies include fractures filled with calcite as well as sparite and micro-sparite fossil fragments. The presence of perforate small benthic foraminifera and fragments of echinoid and red algae indicated a proximal middle ramp setting.
(A1) Foraminifer bioclast packstone. This microfacies consists of two sub-facies.
(A1-1) Echinoderm foraminifer brachiopod packstone. The facies, dominated by the fragments of brachiopods (20%), small miliolids (10%), pelagic foraminifera (10%), perforate benthic foraminifera (10%), bivalves and echinoderm fragments, is well sorted and has a packstone texture. The facies is related to shallow parts of the open marine, which suggests proximal middle-ramp settings. The occurrence of Amphistegina indicates the photic zone of shallow water (Romero et al., 2002). The abundance of benthic foraminifera is few due to wave performance on the basin slope.
(A1-2) Pelecypod foraminifer echinoderm packstone. The facies is dominated by the echinoderm (20%). Moreover, bivalves and pelagic foraminifera (10%) in a micritic matrix are seen.
(B) Barrier microfacies. In this microfacies, benthic foraminifera, peloids and red algae make up 50%–55% of the major components. The minor components of this microfacies are brachiopods, bivalves and gastropod fragments, with an amount of 7%–10%. Also, traces of weathering, porosity and diagenesis were seen in this microfacies. Facies analysis allows the recognition of several sub-facies as follows:
(B1) Intraclast algal foraminifer grainstone. The facies identified by a grainstone texture contains foraminifera (30%), red algae (25%), intraclast (10%) and small amounts of brachiopod fragments (7%). Common biota is Miogypsinoides sp., Peneroplis and miliolids. Following the standard microfacies types determined by Flügel (1982, 2004), this microfacies is interpreted as a shoal environment above the fair weather wave base (FWWB). This facies is located at the platform margin, separating the open marine from the restricted marine environments.
(B2) Intraclast peloid foraminifer grainstone. In the microsparite matrix of this facies, foraminifera such as Neoalveolina (Borelis), Peneroplis, Dendritina, miliolids (35%), peloids (15%), intraclasts (5%), gastropods and algae are found. The abundance of miliolids is generally seen as evidence of a relatively rich nutrient in the back reef environment (Geel, 2000).
(B3) Peloid foraminifer grainstone. This microfacies contains benthic foraminifera (35%), peloids (15%), bivalves and intraclasts with microsparite matrix in a depositional texture of grainstone. Other elements are foraminifera such as Dendritina, Meandropsina, Borelis and miliolids.
(C) Lagoon-inner ramp microfacies. The faunal assemblage in the inner ramp was influenced by an increase in light and a decrease in the depth of the intra-shelf basin. Hence, specific environmental conditions led to a restricted environmentthat includes ooids, peloids and imperforate foraminifers. Therefore, these are important biotic associations in the depositional environment. Given this matter, facies analysis allows several microfacies to be identified, as follows (Figure 6):
(C6) Foraminifer packstone–grainstone. This microfacies is characterised by bioclasts of benthic foraminifera, crinoids, coralline red algae, bivalve and echinoderm fragments in a packstone–grainstone depositional texture. These facies are formed in an inner ramp environment. This represents a shallow sedimentary setting above the wave base (Flügel, 2010), which consists of three sub-facies:
(C6-1) Sandy echinoderm foraminfer packstone–grainstone. The depositional texture is packstone to grainstone with drusy cement, fenestral fabric and fine quartz grains (25%) in micritic matrix. Moreover, the facies contains benthic foraminifera (20%), echinoderm (7%), intraclasts (5%). The presence of fine quartz grains indicates the proximity of these microfacies to the tidal flat environment.
(C6-2) Coral pelecypod echinoderm foraminifer packstone–grainstone. The matrix is microsparite as well as the fine-grained micrite on the margin. It has geopolitical syncytial and fabric cement in gastropod. Of this facies, benthic foraminifera account for 30%, coral (10%), bivalves (10%), echinoderm (10%), intraclasts and gastropods (each 5%). The packstone–grainstone texture suggests high to moderate energy conditions (Hallock & Glenn, 1986). Also, the diversity suggests a non-restricted environment.
(C6-3) Algal foraminifer packstone–grainstone. This facies is characterised by packstone–grainstone depositional texture and contains pelagic foraminifera (25%), red algae (7%), intraclast and ooids in a micritic matrix, with some parts of sparite (5%). Fossils of this facies are Peneroplis, Dendritina, Austrotrillina, Borelis and Miliolids. The presence of imperforate foraminifera such as Dendritina, Austrotrillina, Peneroplis and miliolids is generally taken as evidence for restricted lagoonal environments (Geel, 2000; Romero et al., 2002). The depositional setting of microfacies C6-3 is known as the lagoonal end of the platform margin. Similar microfacies has been reported from the Asmari Formation in other sections, such as the Khaviz section (Kimiagari, 2006).
(C5) Foraminifer packstone. This microfacies consists of four sub-facies:
(C5-1) Peloid gastropod pelecypod ooid foraminifer packstone. The facies characterised by a fine-grained sparite matrix that is characterised as a packstone texture and contains benthic foraminifera (40%), ooids (15%), bivalves (10%), peloids and gastropods (each 5%). Other components are Valvulina sp. and Bigenerina sp. The existence of abundant porcelaneous imperforate benthic foraminifera such as Archaias, Dendritina and miliolids indicates deposition in a restricted shelf lagoon and the upper photic zone (Hallock & Glenn, 1986; Holzman et al., 2001; Romero et al., 2002). According to Geel (2000), an abundance of miliolids is taken as evidence for restricted lagoonal and relatively nutrient-rich conditions.
(C5-2) Peloid foraminifer packstone. The facies characterised by a fine-grained sparite matrix that is characterised as a packstone texture and contains benthic foraminifera (20%), peloids (15%) and bivalves (ca. 5%). Diagenetic processes of this facies include recrystallisation and hematite. Other components are Dendritina, Borelis haueri, Textolaria and miliolids. The occurrence of imperforate foraminifera such as Borelis and miliolids indicates restricted lagoon conditions (Geel, 2000; Romero et al., 2002).
(C5-3) Algal foraminifer packstone. This facies is characterised by a fine-grained micritic matrix that is characterised as a packstone texture and contains benthic foraminifera (35%), echinoderm (20%), red algae (15%), bivalves (5%) and gastropods (8%). An important diagenetic phenomenon of this facies is the calcification of bioclasts. Other components are Miogypsinoiedes, Textolaria and Miogypsina. Geel (2000) states that Miogypsinoids lived in shallow waters of average salinity.
(C5-4) Pelecypod algal foraminifer packstone. A packstone texture characterises this facies and contains benthic foraminifera (40%), red algae (Lithothamnion) (15%), bivalves (10%) and very small peloids (ca. 5%).
(C4) Algal foraminifer wackestone. This microfacies consist of two sub-facies:
(C4-1) Ostracods pelecypod foraminifer wackestone. This facies characterised by a fine-grained sparite matrix that is characterised as a wackestone texture and contains benthic foraminifera (10%), bivalves (7%) and ostracods (ca. 5%). The diagenetic process of this facies is secondary porosity.
(C4-2) Algal foraminifer wackestone. The facies is characterised by wackestone depositional texture and contains benthic foraminifera (15%), algal red (7%), intraclast and peloids (ca. 5%) in a fine-grained sparite matrix. The diagenetic process of this facies is boringporosity in bivalves.
(C3) Bioclast lime mudstone. This microfacies consists of three sub-facies:
(C3-1) Lime mudstone. This microfacies has a microcrystalline matrix and very few quartz grains. The mud matrix in this microfacies indicates a low- to moderate-energy environment.
(C3-2) Intraclast lime mudstone. The facies are characterised by non-laminated and homogenous lime mudstone (95%) and intraclast (2%–3%). One of the diagenetic processes in the facies is calcite succession.
(C3-3) Bioturbated bioclast lime mudstone. This facies is characterised by lime mudstone texture and contains micritic bioclast (8%–10%). The diagenetic processes of this facies include secondary porosity and bioturbation.
(C2) Bioturbated bioclast peloid wackestone. This microfacies consists of two sub-facies:
(C2-1) Bioturbated intraclast bioclast peloid wackestone. The facies is characterised by wackestone texture that contains algae (10%), bioclast (5%), micritic intraclast (10%) and peloids (20%). Diagenetic processes of this facies include calcification and bioturbation.
(C2-2) Bioturbated foraminifer gastropod pelecypod wackestone. The facies is characterised by wackestone texture that contains larger foraminifera (30%), gastropods (12%), bivalves (7%), ostracods and micritic intraclast. The association of the biogenic components suggests deposition in a more restricted environment with a more onshore position than the larger foraminiferal microfacies types (Rasser & Piller, 2004). The environmental processes of this facies include bioturbation.
(C1) Peloid wackestone–packstone. This facies consists of peloids (30%) in a wackestone–packstone texture. The presence of peloids suggests a restricted, shallow environment in the inner-ramp settings.
(D) Tidal flat (supratidal)-inner ramp microfacies.
Fenestral dolomudstone with evaporate cast. This facies belongs to the Sabkha environment, which is characterised by yellow–brown, massive and fine-grain dolomite without any fossils. Dolomite has a microcrystalline matrix that has been recrystallised. It has vuggy pores, a bird’s eye and fenestrate structures. Due to the lack of fossils or fauna, bird’s eye fabric and dolomite with silt-size grains, these facies were deposited in a low-energy tidal flat environment. Flügel (2004) states that unfossiliferous fine-grained dolomicrite with autogenetic evaporite minerals occurs in tidal flat and arid evaporitic coasts. Dolomudstone samples can indicate that the weather is warm and dry. This assemblage is similar to the facies belt microfacies No. 9 of Wilson -(1975) and Flügel (2004) as well as the facies belt No. 1 of Buxton and Pedly (1989), deposited in the inner ramp environment in the supratidal zone. In other parts of Iran, the Asmari Formation has shown similar microfacies (Vaziri-Moghaddam et al., 2010).
Facies Description and Interpretation in the MakhmalKuh Section:
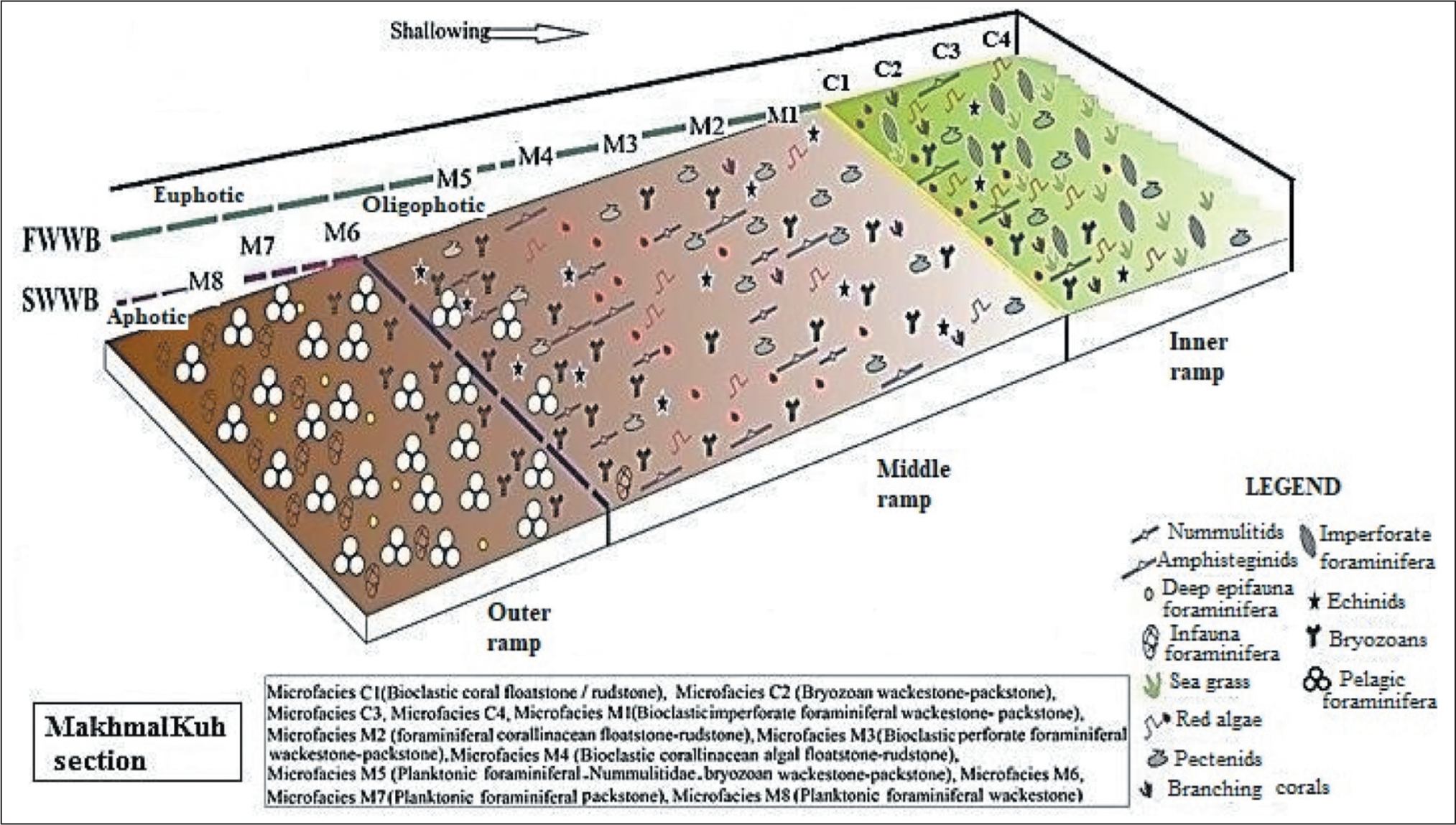
The Asmari Formation at the MakhmalKuh section is subdivided into 12 different microfacies, each characterised by a petrographic analysis and foraminiferal assemblage (Figure 7). Based on the sedimentological analysis, two facies belts can be recognised: lagoon (C) and open marine (M). The identified facies arranged by their depositional setting (shallow to deep) in the studied section as follows:
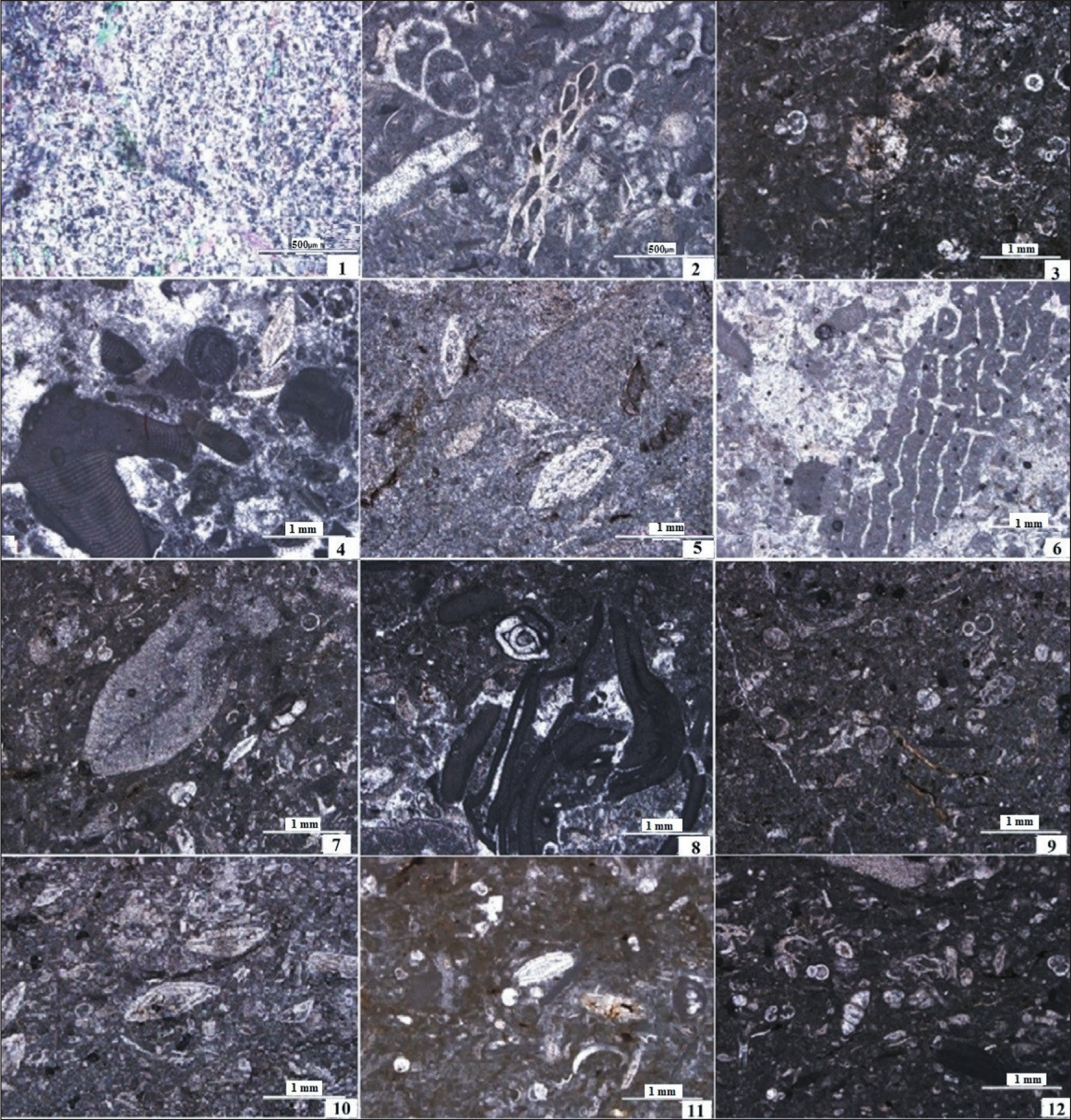
Microfacies of the Asmari Formation in MakhmalKuh Section; 1. (C1, Sample KH.83); 2. (C2, Sample KH.40, Bryozoan Wackestone-Packstone); 3. (C3, Sample KH.85); 4. (C4, Sample KH.25); 5. (M1, Sample KH.17, Bioclast Imperforate Foraminiferal Wackestone-Packstone); 6. (M2, Sample KH.55, Foraminiferal Corallinaceae Floatstone-Rudstone); 7. (M3, Sample KH.62, Bioclast Perforate Foraminiferal Wackestone-Packstone); 8. (M4, Sample KH.11, Bioclastic Coraline Algal Floatstone-Rudstone); 9. (M5, Sample KH.84, Pelagic Foraminifera, Nummulitidae, Bryozoan Wackestone-Packstone); 10. (M6, Sample KH.49, Fossiliferous Marl); 11. (M7, Sample KH.79); 12. (M8, Sample KH.80).
Facies related to the lagoon (Inner ramp). This facies association includes four microfacies:
Microfacies C4 (Bioclast coral floatstone–rudstone). The most fundamental elements of sub-facies C1 generally are coral fragments, benthic foraminifera (Elphidium, Miogypsina, Amphistegina, Borelis and miliolid) and bioclast fragments. Corals represent the euphotic area. Also, coral colonies can spread in seagrass environments. In addition, Elphidium, Borelis, Amphistegina and Miogypsina species have been widely reported from shallow marine environments. These faunas and small miliolids suggest high-energy conditions around coral adjacent to the seagrass meadows (euphotic zone). The presence of miliolids is generally taken as evidence of relatively nutrient-rich environments and lagoons (Geel, 2000). Also, the abundance of calcite microcrystalline (lime mud) in the sub-facies indicates shallow environments under dispersed reef masses. This facies represents the proximal inner ramp environment in the euphotic zone colonised by seagrass (Brandano et al., 2009; Pomar et al., 2014). As stated above, the inner ramp lithofacies consists of horizontal beds with abundant porcellaneous foraminifera, Borelis and miliolids.
Microfacies C3. This facies contains marl along with the benthic foraminifera, which has very little variation. In this facies, Ammonia sp., Ammonia tepida, Ammonia beccarii and Elphidium granosum are present. Pelagic foraminifera are abundant, and the planktonic foraminifera to benthic ratio is about 70%. Ammonia beccarii is a euryhaline taxon that can exist from marine to brackish environments but is particularly predominant in marginal marine environments. Ammonia tepida and Elphidium granosum are also associated with environments such as lagoons. Elphidium granosum is hypersensitive to oxygen variation. Therefore, the Ammonia predominance along with Elphidium granosum indicates that, except for salinity fluctuations, there was no oxygen stress on the environment of these facies.
Microfacies C2 (Bioclast wackestone–packstone). The dominant components of this facies are the abundance of macrofauna consisting of bryozoan fragments, debris of corals, red algae and small foraminifera such as miliolids. Subordinate components are benthic foraminifera such as Textularia, Elphidium and other biological constituents of these facies. The decrease in the number of foraminiferal species and the presence of bryozoan are probably related to an increase in nutrient levels (mesotrophic conditions) in the proximal inner ramp.
Microfacies C1. This facies comprises thin- to medium-layer anhydrite-, which contains pelagic facies. Anhydrite deposition in the pelagic facies indicates sedimentation in the relatively deep hypersaline marine basin with unstable and stressful conditions.
Facies related to the open marine (outer ramp–mid ramp). Facies analysis allows the recognition of eight microfacies:
Bioclastic imperforate foraminiferal wackestone–packstone (Microfacies M1). The major components of this microfacies are skeletal grains of imperforate benthic foraminifera such as Borelis, Peneroplis, Dendritina, Meandropsina, Spirolina and miliolids with a wackestone–packstone texture. The larger benthic foraminifera with hyaline walls include both Amphistegina and Elphidium. Minor components are small benthic foraminifera, Ammonia, Textularia and Discorbis, fragments of echinoids, molluscs and corallinacean algae (Lithoporella). The assemblages of benthic foraminifera (imperforate) indicates deposition in a shallow environment with low turbulent. This indicates oligotrophic to mesotrophic conditions at low depths. The thick-shelled Amphistegina with seagrass also indicates life in the photic zone, relatively shallow water and water conditions with moderate energy (Romero et al., 2002).
Foraminiferal corallinaceae floatstone–rudstone (Micro-facies M2). Nodules and branches of red algae characterise this microfacies. The benthic foraminifera include both perforate (i.e., Amphistegina, Miogypsina) and imperforate forms (e.g., Meandropsina, miliolids). Minor components are fragments of molluscs, bryozoan, echinoids, coral and red algae (Lithothamnion). Perforated foraminifera, such as Amphistegina and Miogypsina, are in close contact with seagrass. Also, the abundant microcrystalline matrix with seagrass indicates shallow water conditions. On the other hand, the coexistence of imperforate and perforate foraminifera indicates the depositional setting was a semi-restricted lagoon (Amirshahkarami, 2013). Therefore, the skeletal compound and the stratigraphic position indicate sedimentation in the initial zone of mid-ramp.
Bioclastic perforate foraminiferal wackestone–packstone (Microfacies M3). The significant components of microfacies are heterotrophic forms (i.e., bryozoan, echinoids and molluscs) with a wackestone–packstone texture. Corallinacean red algae, Mesophyllum, Lithothamnion and perforate benthic foraminifera are other components. Red algae with green algae and coral fragments are other examples of this microfacies. The larger benthic foraminifera include both Amphistegina and Operculina. Large foraminifers are often attributed to light intensity, water energy, rising sea levels due to global warming and the development of tropical habitats (Halluk and Glenn, 1986; Hohenegger, 2000; Bassi et al., 2007). There are also small benthic foraminifera such as Lobatula, Elphidium, Cibicides and Textularia. According to Hottinger (1997), Nummulitids inhabit the deepest environments among the observed components. Other components, such as molluscs shell fragments and bryozoan with echinoids crustaceans, confirm this interpretation. The coexistence of large perforate foraminifera belonging to the deep areas, such as Amphistegina and Nummulitidae, together with smaller foraminifera and Melobesioidae algal, is characteristic of the middle ramp setting and the oligophotic zone.
Bioclastic corallinacean algal floatstone–rudstone (Micro-facies M4). This microfacies is mainly characterised by corallinacean algae colonies (i.e., Sporolithon and Lithotha-mnion) and Rhodolite. Rhodolites are mainly branch-shaped. The texture is floatstone–rudstone—other components, such as echinoids and bivalve shell fragments with coral. Among the large foraminifera, Amphistegina and Operculina have been identified. Elphidium, Textularia, Lobatula, Acervulinid and Sphaerogypsina are also present. The presence of shallow-water fauna, debris bioclasts and deep-sea organisms indicates that marine currents carry out sedimentation. The presence of species such as Operculina and Amphistegina, together with algal components such as Melobesioidae and Sporolithacea, suggests an oligotrophic environment within the middle ramp setting.
Planktic foraminiferal, Nummulitidae, Bryozoan wackestone–packstone (Microfacies M5). The main skeletal grains consist of bryozoa, planktonic foraminifera and fragments of benthic foraminifera (i.e., Cibicides, Textularia, Elphidium, Buliminids, Operculina and Amphistegina). Typical biota includes echinoids and molluscs. The major components of this microfacies are light-independent fauna such as bryozoans, while light-dependent organisms are present in low percentages. The abundance of delicate and straightforward bryozoans reflects low-energy conditions in offshore habitats. Bryozoa belong to the Cyclostomate and Cheilostomate families. This facies is also characterised by deep-water large benthic foraminifera (i.e., Operculina and Amphistegina). The presence of deeper foraminifera, such as Nummulitids, suggests sedimentation in the oligophotic zone (lower limit of the photic zone). Romero et al. (2002) reported the Nummulitidae in open marine conditions. Large and flat tests of perforate foraminifera are the dominant microfauna of the middle ramp, probably because they were the best-adopted fauna for environmental conditions such as low energy, oligotrophic and normal salinity (Romero et al., 2002).
Fossiliferous marl (Microfacies M6). This microfacies is characterised by benthic and planktonic foraminifera in a depositional texture of marl. Globigerina and Globigerinoides are the most important planktonic foraminifera. The benthic foraminifera include Elphidium crispum, E. maculleum, E. granusom, E. fichtellaneum, Cibicidoides sp., Eponides sp., Cibicides sp., Cibicides lobatulus and Heterolepa dutemplei. Echinodermata fragments and spines, bryozoan and gastropods are also present in some specimens. The fine-grained composition and the high abundance of planktonic foraminifera in marl sediment reflect sedimentation in a low-energy middle ramp environment, down of storm waves. Also, the presence of benthic taxa such as Heterolepa, Cibicides and Cibicidoides represents a stable environment with good ventilation, adequate oxygen level, water currents and oligo-mesotrophic nutrient conditions. The distal mid-ramp is differentiated from the shallower depth by a decrease in the flatness and size of the perforate foraminifera.
Planktic foraminiferal packstone (Microfacies M7). This facies is characterised by a shallow diversity of benthic foraminifera and a greater micritic matrix. The most abundant benthic fauna are Uvigerina, Reticulophragmium and Pappina. Here, pelagic foraminifera, including Globigerinoides and Globigerina, are relatively large with a planktonic/benthic ratio reaching up to 70%. The shallow diversity of benthic foraminifera and the dominance of a particular group, such as Uvigerina, reflect low oxygen content at the sediment-water contact surface. Especially, when the inlet flow of organic carbon dominates the environment, it tends to increase population, exhibits opportunistic behaviour and greatly proliferates under conditions of oxygen deficiency. Also, Reticulophragmium is a species able to withstand stressful environmental conditions such as low oxygen. Therefore, this facies represents an unstable environment with low oxygen levels in a distal middle ramp that can be caused by some factors affecting the depositional environment.
Planktic foraminiferal wackestone (Microfacies M8). This facies is characterised by the abundant presence of planktonic foraminifera and some benthic foraminifera. The planktonic/benthic foraminiferal ratio is 70%–90%. The foraminiferal assemblages have good to moderate preservation. This microfacies is characterised by abundant and diverse planktonic foraminifera such as Globigerina, Globigerinoides and Globorotalia. Among the biotic associations, Rectuvigerina is the most well-known species of endemic foraminifera. In addition, Uvigerina, Bulimina, Bolivina and Nonion are also present. The abundance of endogenous taxa compared to surface taxa may represent mesotrophic conditions. In addition, the high ratio of planktonic/benthic and the presence of benthic foraminiferal assemblages represent the outer ramp setting.
Sedimentary Model
To reconstruct the sedimentary model of each basin, several data, such as microfacies analysis and a systematic method for using the data to arrive at an appropriate interpretation are necessary. Based on microfacies analysis, four major environments can be recognised in the Oligo-Miocene Asmari Formation in the Razan section: tidal flat, lagoon, barrier and open marine. The tidal flat and lagoon environments are known as inner ramps. The tidal flat setting is characterised by unfossiliferous fine-grained dolomicrite, and the lagoon (microfacies C1–C6) consists mainly of imperforate benthic foraminifera and a finer-grained matrix. In fact, these facies indicate the area of the upper photic zone and low energy in a restricted lagoon depositional environment or the inner ramp (Geel, 2000; Romero et al., 2002). The barrier setting consists of foraminifera such as Miogypsinoides sp., Peneropelis, Dendritina, Neoalveolina, Meandropsina and miliolids in a grainstone texture. This environment reveals different facies (microfacies B1–B3), which are separable from each other by their components. Microfacies in this environment is characterised especially by miliolids and peloids, which is characteristic of the highly energetic part of the shoal. In the meantime, microfacies B1 separates open marine and restricted marine environments. The open marine environment is recognised by the middle ramp setting. The open marine environment or middle and outer ramp setting (microfacies A1–A4) mainly consists of pelagic foraminifera, benthic foraminifera (Nummulites, Neoalveolina, Rotalia, Dendritina and Meandropesina) and deep marine organisms. The middle ramp can be divided into proximal and distal. The proximal, middle ramp is characterised by oolitic and bioclastic in a skeletal packstone–grainstone texture. In other words, this also shows a transition from grainstone texture (from a barrier environment) to packstone and wackestone textures in the open marine environment. Skeletal grains are dominated by echinoids, Mollusca, bryozoan, corallinacean and benthic foraminifera. The distal middle ramp is identified by the perforate benthic foraminiferal assemblage, such as Nummulites and Lepidocyclina, and the textures corresponding to an increase in the water depth. Therefore, based on the interpretation of microfacies, the transfer to the open marine shows a deeper trend. Also, diversity is more in barrier microfacies but less in open marine microfacies (deepest microfacies), which implies the diversity in these facies is controlled by ecological factors, especially nutrients and light. The gradual transition between all facies indicates a gradient carbonate ramp platform (Figure 8).
Depositional Sub-Environments of the Asmari Formation, Razan Section (Lorestan Province).

In the MakhmalKuh section, the carbonate platform is usually characterised by uniform carbonate production by large benthic foraminifera and red algae above the optical zone and planktic foraminifera below the optical zone. Based on the detailed analysis of the facies as well as the dependence of the creatures on light, the ramp in the Asmari Formation in the MakhmalKuh section is divided into three parts: inner ramp, middle ramp and outer ramp. Therefore, the sedimentary model of the Asmari Formation that is formed from Rupelian to Aquitanian in this area is as follows: (a) inner ramp/euphotic and mesophotic zones, (b) mid ramp/oligophotic zone and (c) outer ramp/aphotic zone or deep zone (Figure 9).
Depositional Sub-Environments in the Asmari Formation, MakhmalKuh Section (Abbreviations: FWWB: Fair-Weather Wave Base; SWB: Storm Wave Base (Adapted from IFP, 2000).
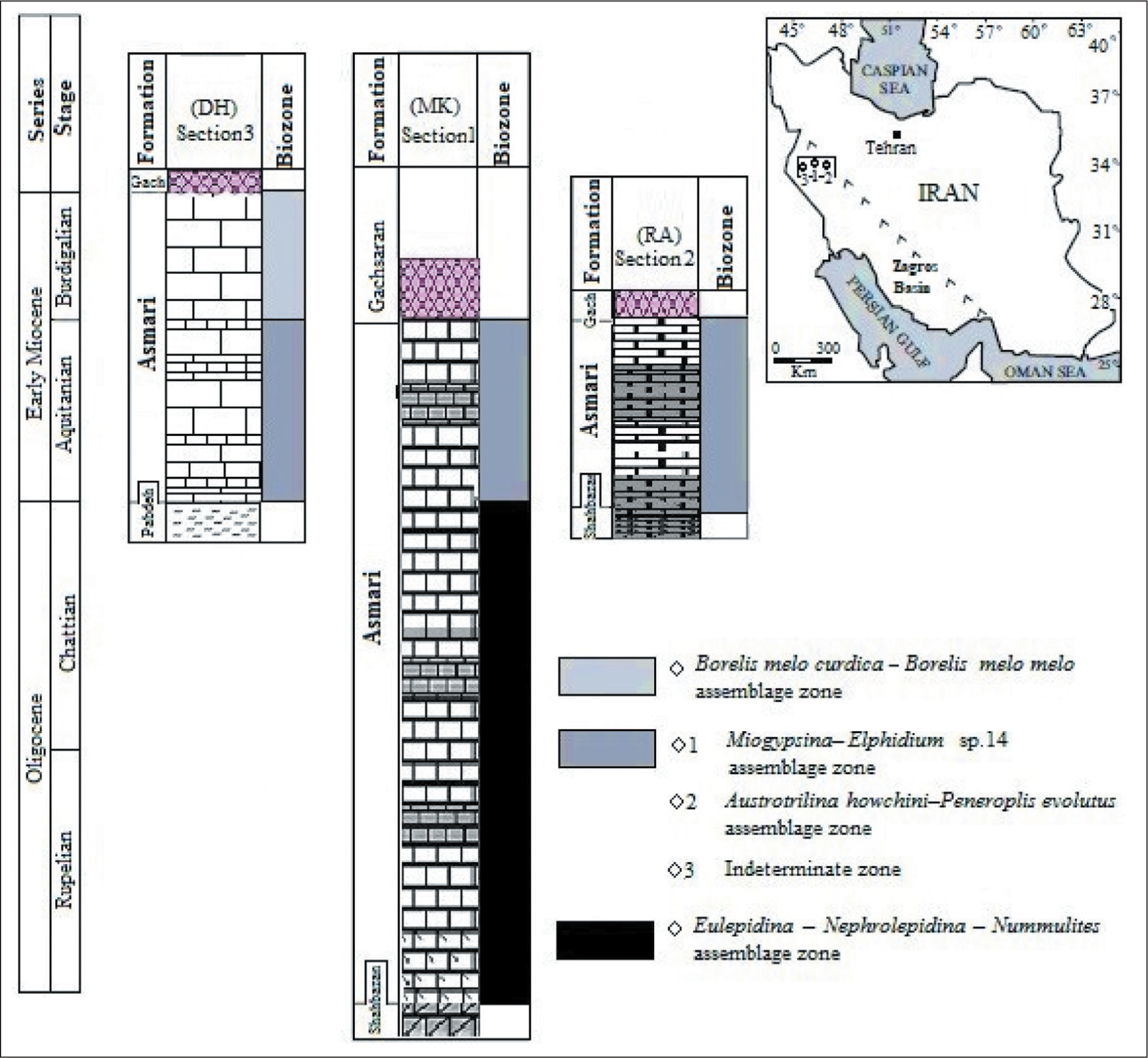
In this study, we also compared the Asmari Formation in the studied sections with the Dehluran section in western Lorestan, in the Zagros Basin (Table 1). Here, during the Oligocene, the Asmari Formation was deposited in the MakhmalKuh section in the northwest of Khorram Abad city. At the same time, the Razan and Dehloran regions were exposed. Alavi (2004) interpreted that this could be due to the response of the basin floor to a new phase of faulting in the hinterland region. Subsequently, at the beginning of the Miocene (Aquitanian), the transgressive sea caused the Asmari Formation to be deposited throughout the region. Gradually, at the beginning of the Burdigalian, with the continuation of the transgressive progress, the Asmari Formation deposited in the western areas (Dehloran section), but some eastern parts of the Lorestan province (Razan section) were exposed. Finally, it is observed that the age of the Asmari Formation becomes younger from the eastern to western parts of Lorestan province (Figure 10).
Biozonation of the Asmari Formation (Laursen et al., 2009; van Buchem et al., 2010) and Comparison Between the Studied Sections (1. MakhmalKuh Section, 2. Razan Section) with Dehluran Section (The Third Section with Coordinate N: 32° 51' 20'' E: 43° 54' 3'', Vaziri-Moghaddam et al., 2010).

Correlation of Studied Sections (MK , MakhmalKuh Section and RA , Razan Section) with Dehloran Section (DH ) in Lorestan Province.
Conclusion
The Razan and MakhmalKuh stratigraphic sections of the Asmari Formation in the Lorestan foreland basin were studied to determine the depositional environment and accurate age. Based on biostratigraphic data, the A. howchini–Peneroplise volutus assemblage zone has been recorded in the Razan section, which is considered to be the early Miocene (Aquitanian) in age. These fauna assemblages are correlated with the SB24 zone in the Mediterranean region. Also, based on biostratigraphic data, two assemblage zones have been recorded in MakhmalKuh section. The first assemblage is Eulepidina–Nephrolepidina–Nummulites, which is considered to be Oligocene in age. The presence of Nummulites and Eulepidina, along with Nephrolepidina indicates Rupelian–Chattian intervals. The second assemblage zone is Miogypsina–Elphidium sp. 14, which indicates the early Miocene (Aquitanian) age. These faunal assemblages are correlated with the SB22, SB23 and SB24 zones in the Mediterranean region. Due to microfacies analysis, the inner ramp, middle ramp and outer ramp settings (open marine environment) are present in the Asmari Formation at the Razan section. Inner ramp settings (between tidal flat and barrier) are represented by facies D1 to B3, and open marine environments, namely middle and outer ramp settings, are represented by facies A1–A4, respectively. Also, based on detailed facies analysis, determined several marine environments, including open marine and lagoon in the Asmari Formation in the MakhmalKuh section. These depositional models of the Asmari Formation, which was formed in this region from Rupelian to Aquitanian, are as follows: (a) Inner ramp, euphotic to mesophotic zones; (b) middle ramp, oligophotic zone and (c) outer ramp, aphotic zone or the deep zone. The outcrops of the Asmari Formation at Razan and MakhmalKuh areas located in the northwest of Zagros are Aquitanian and Rupelian–Aquitanian in age, respectively. In this regard, while deposition of the Asmari Formation continued during the Rupelian–Chattian in the lower part of the MakhmalKuh section, the Razan section was still the site of basinal deposition of the Shahbazan Formation. Also, a comparison of the Asmari Formation in the studied cells with the Dehloran section in the west of Lorestan shows that the age of the Asmari Formation becomes younger from the eastern to western parts of Lorestan province.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors received no financial support for the research, authorship and/or publication of this article.