
Abstract
Background
Administration of onabotulinumtoxinA (BoNT-A) to peripheral tissues outside the calvaria reduces the number of days chronic migraine patients experience headache. Because the headache phase of a migraine attack, especially those preceded by aura, is thought to involve activation of meningeal nociceptors by endogenous stimuli such as changes in intracranial pressure (i.e. mechanical) or chemical irritants that appear in the meninges as a result of a yet-to-be-discovered sequence of molecular/cellular events triggered by the aura, we sought to determine whether extracranial injections of BoNT-A alter the chemosensitivity of meningeal nociceptors to stimulation of their intracranial receptive fields.
Material and methods
Using electrophysiological techniques, we identified 161 C- and 135 Aδ-meningeal nociceptors in rats and determined their mechanical response threshold and responsiveness to chemical stimulation of their dural receptive fields with TRPV1 and TRPA1 agonists seven days after BoNT-A administration to different extracranial sites. Two paradigms were compared: distribution of 5 U BoNT-A to the lambdoid and sagittal sutures alone, and 1.25 U to the sutures and 3.75 U to the temporalis and trapezius muscles.
Results
Seven days after it was administered to tissues outside the calvaria, BoNT-A inhibited responses of C-type meningeal nociceptors to stimulation of their intracranial dural receptive fields with the TRPV1 agonist capsaicin and the TRPA1 agonist mustard oil. BoNT-A inhibition of responses to capsaicin was more effective when the entire dose was injected along the suture lines than when it was injected into muscles and sutures. As in our previous study, BoNT-A had no effect on non-noxious mechanosensitivity of C-fibers or on responsiveness of Aδ-fibers to mechanical and chemical stimulation.
Discussion
This study demonstrates that extracranial administration of BoNT-A suppresses meningeal nociceptors’ responses to stimulation of their intracranial dural receptive fields with capsaicin and mustard oil. The findings suggest that surface expression of TRPV1 and TRPA1 channels in dural nerve endings of meningeal nociceptors is reduced seven days after extracranial administration of BoNT-A. In the context of chronic migraine, reduced sensitivity to molecules that activate meningeal nociceptors through the TRPV1 and TRPA1 channels can be important for BoNT-A’s ability to act as a prophylactic.
Introduction
Botulinum neurotoxin type A (BoNT-A) is an effective prophylactic drug for chronic migraine (1,2). Although widely used in clinical practice, the mechanism by which it reduces migraine days in chronic migraineurs is not yet fully understood. Of the many questions that await answers, the most puzzling one is how administration of BoNT-A to peripheral sites outside the calvaria reduces the frequency of migraine headache—a neurological condition believed to originate mainly in the brain (3) and involve activation of pain fibers inside the calvaria (4).
A significant advance in answering this question was made by the discovery of a network of sensory fibers that bifurcate from parent axons of intracranial meningeal nociceptors and reach extracranial tissues such as periosteum and pericranial muscles by crossing the calvarial bones—from inside to outside—through the sutures (5–7). Biologically, the importance of this newly discovered anatomic pathway is that it provides a neural substrate for direct communication between extracranial and intracranial sensory events involving activation of peripheral nociceptors at their termination sites in the various tissues they innervate.
Integrating this “suture pathway” into a wider framework for testing novel hypotheses about BoNT-A mechanisms of action in migraine, we recently showed that local administration of BoNT-A to extracranial sutures renders the suture branches of the meningeal nociceptors, but not the meningeal nociceptors themselves, mechanically insensitive (8). In the discussion of that study, we raised the possibility that the intracranial meningeal nociceptors were not inhibited by the extracranial administration of BoNT-A because we did not give the drug enough time to act on more remote sites within the same axon(s) to which it was applied. Specifically, we hypothesized that the findings reflect the very limited four-hour period we allowed the drug to act and that a longer time is required to determine whether extracranial administration can interfere with proximal intracranial transduction. In support of this notion, we showed that when BoNT-A is applied directly to the dura, it can inhibit naïve C-fiber meningeal nociceptors responses to mechanical nociception, and reverse and prevent their sensitization by inflammatory mediators (8).
Attempting to design a study that is more relevant to the therapeutic application in patients and test the time hypothesis more directly, we designed a set of experiments in which effects of extracranial injections of BoNT-A on intracranial meningeal nociceptors were tested 168 hours (i.e. seven days) after drug administration. Based on BoNT-A’s ability to inhibit mechanical nociception (8) and block soluble nethylmaleimide-sensitive factor attachment protein receptor (SNARE)-dependent cell surface expression of transient receptor potential (TRP) receptors (9), on the presence of both transient receptor potential vanilloid type 1 (TRPV1) and transient receptor potential cation channel, subfamily A, member 1 (TRPA1) channels in the dura (10), on their role as transducers of noxious stimuli and their ability to cause release of substance P (SP) and calcitonin gene-related peptide (CGRP) upon activation (11), and on their role in mediating peripheral sensitization of nociceptors in response to inflammation (12–17), in the current study we tested BoNT-A’s effects, when administered extracranially, on intracranial meningeal nociceptors’ responses to dural stimulation with mechanical forces and with the chemical agents capsaicin, a selective TRPV1 agonist (18), and mustard oil (MO), a powerful agonist of TRPA1 (19), which may also act partly through TRPV1 (20).
Methods
All experiments were approved by the standing committee on animals of Harvard Medical School and Beth Israel Deaconess Medical Center, in accordance with the United States National Institutes of Health Guide for the Care and Use of Laboratory Animals as well as the guidelines of the Committee for Research and Ethical Issues of the International Association for the study of Pain.
Experimental paradigm
Male Sprague-Dawley rats weighing 150–170 g were briefly anesthetized (2% isoflurane) and injected with BoNT-A (onabotulinumtoxinA; final dose = 5 units) using two different injection paradigms. In the first paradigm, four injections of BoNT-A (each containing 1.25 units diluted in 5 μl saline) were made along the lambdoid (two injection sites) and sagittal (two injection sites) sutures (Figure 1(a)). In the second paradigm, eight BoNT-A injections (each containing 0.625 units) were made: two along the sagittal suture, two in the temporalis muscle and four in the trapezius muscle (Figure 1(b)). All injections were given at the animal facility between 1 p.m. and 4 p.m. Seven days later, injected rats (by now weighing 250–300 g—housed in a specific pathogen-free facility equipped with solid bottom cages of 1800 cm2 of floor space and hardwood chip bedding; kept at 12 hours light/dark cycle, 22–26℃, and inspected twice daily for signs of stress or discomfort) were anesthetized with urethane (1.8 g/kg intraperitoneally (i.p.)) and prepared for single-unit recording of C- and Aδ-meningeal nociceptors. All electrophysiological experiments were carried out between 9 a.m. and 6 p.m. in a room especially fitted for electrophysiological recording in deeply anesthetized rats. To test for effects of extracranial injections of BoNT-A on responses of meningeal nociceptors to stimulation of their intracranial (dural) receptive fields, testing determined mechanical response threshold and responses to topical application of the TRPV1 agonist capsaicin and the TRPA1 agonist MO. For comparisons, similar experiments were carried out in 47 naïve and 25 sham rats (i.e. rats in which recordings were made seven days after they were anesthetized with 2% isoflurane and injected with vehicle at the same sites as the muscle-plus-suture group). As the results from the two control groups were very similar (e.g. scatterplots in Figure 2(b) and (c)) and not statistically different, they were combined for statistical comparisons.
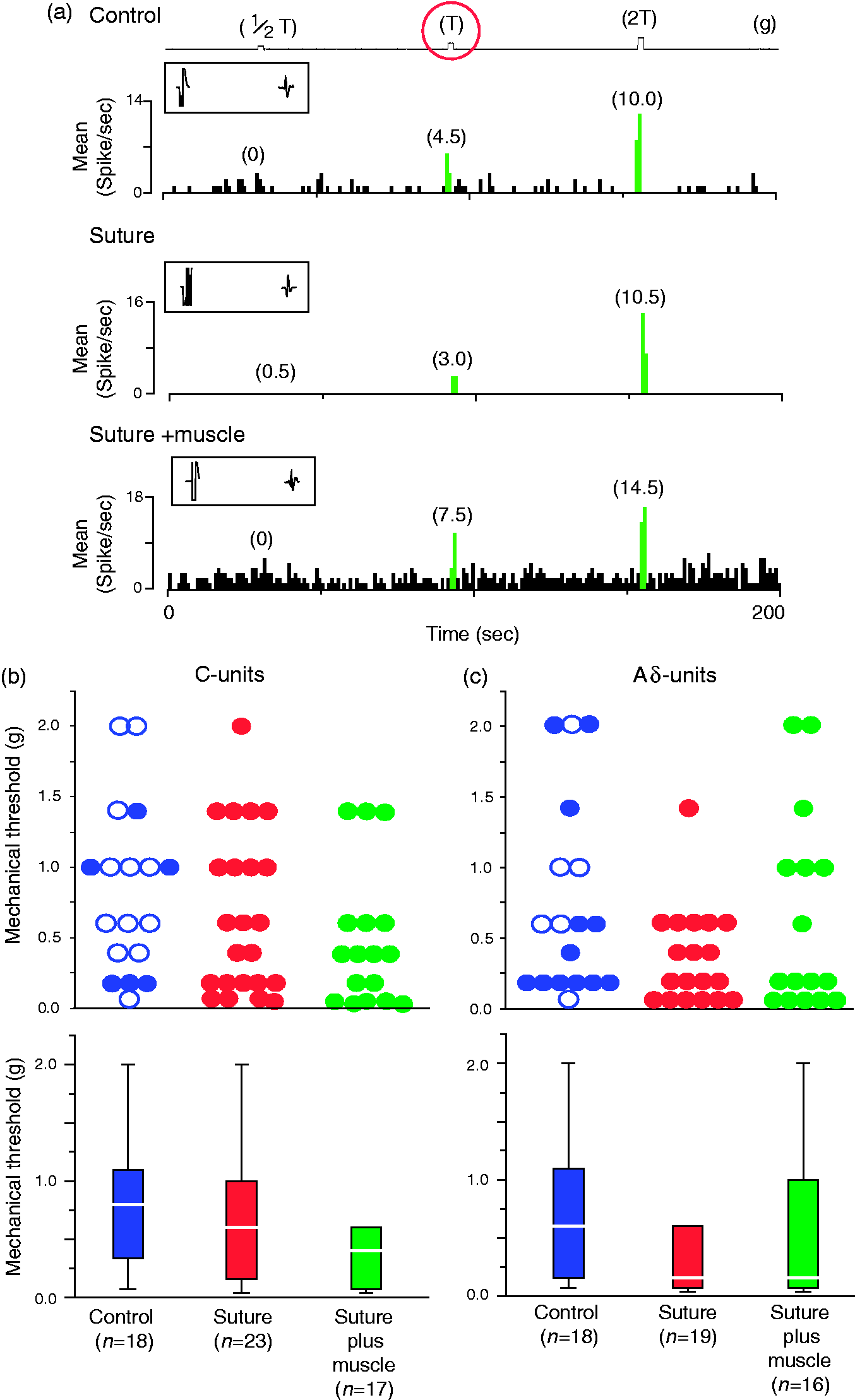
Injection paradigm. (a) The suture paradigm consisted of four injections of BoNT-A, each containing 1.25 units, along the superior sagittal and transverse sinuses. (b) The suture-plus-muscle paradigm consisted of eight injections of BoNT-A, each containing 0.625 units. As depicted by the red dots, four injections were made in the clavicotrapezius muscle, two in the temporalis muscle, and two along the superior sagittal suture. BoNT-A: onabotulinumtoxinA. Mechanical threshold for activation of C- and Aδ-meningeal nociceptors from their dural receptive fields are not affected by extracranial BoNT-A injections. (a) Mechanical response threshold of three different C-type meningeal nociceptors recorded in control rat (top), and in rats treated seven days earlier with BoNT-A injections into the sutures (middle) or the suture plus muscles (bottom). Boxed inset in each plot shows the shock artifact, the spike waveform, and the response latency. (b), (c) Mechanical thresholds for all C-units (b) and Aδ-units (c), shown as scattergraphs (top) and boxplots (bottom). Blue, control rats; red, suture-injected rats; green, suture-plus-muscle injected rats. In the scatterplots for the control rats, the filled and open circles represent values recorded in naïve and saline-injected rats, respectively. Boxplots illustrate median (thick horizontal line), interquartile range (25th–75th percentile; lower and upper box boundaries) and observations below and above the 25th and 75th percentile (whiskers) of the mechanical response threshold. Note that mechanical threshold values represent the smallest mechanical force capable of activating the neurons (i.e. innocuous force). BoNT-A: onabotulinumtoxinA.
Surgical preparation and identification of meningeal nociceptors
Urethane-anesthetized rats were mounted on a stereotaxic frame and their physiological parameters were monitored continuously. Core temperature was kept at 37℃ using a feedback heating blanket. End-tidal CO2 was kept at 3.5–4.5%. To gain access to the trigeminal ganglion, a small craniotomy was performed 2 mm lateral to the sagittal suture and 2 mm posterior to Bregma. To gain access to the dural receptive field, another craniotomy was performed over the ipsilateral transverse sinus, from 1 mm anterior to 1 mm posterior to Lambda. The dura was kept moist throughout the experiment using modified synthetic interstitial fluid (SIF, pH 7.2) containing 135 mM NaCl, 5 mM KCl, 1 mM MgCl2, 5 mM CaCl2, 10 mM glucose, and 10 mM HEPES).
Identification of meningeal nociceptors in the trigeminal ganglion was performed as described in detail before (21,22). Briefly, a platinum-coated tungsten microelectrode (impedance 50 kΩ; FHC Inc, Bowdoinham, ME) was lowered into the left trigeminal ganglion (2 mm behind Bregma and 2 mm lateral midline) while a constant pulse of electrical stimuli (0.5 ms pulse, 5 mA, 0.5 Hz) was delivered to the dura overlying the ipsilateral transverse sinus through a bipolar stimulating electrode. Once a neuron was identified whose response latency to the electrical stimulation of the dura was constant, it was characterized as a C- or an Aδ-unit based on its conduction velocity (C-units ≤1.5 m/sec, Aδ-units between 1.5 and 5 m/sec) and then verified that it was in fact a meningeal nociceptor by exhibiting discrete bursts of activity in response to mechanical stimulation of the dural receptive field with calibrated von Frey monofilaments.
A real-time waveform discriminator was used to create and store a template for the action potential evoked in the neuron under study by electrical pulses on the dura; spikes of activity matching the template waveform were acquired and analyzed online and offline using Spike 2 software (CED, Cambridge, UK).
Mechanical and chemical stimulation
Mechanical receptive fields were initially mapped using a series of calibrated von Frey monofilaments exerting pressure stimuli in the range of 0.04–2.00 g. Once mapped, quantitative mechanical stimuli were delivered to the most sensitive area of the dural receptive field using a servomotor force-controlled stimulator (Series 300B, Aurora Scientific, Aurora, ON, Canada) fitted with a flat-ended plastic cylinder. The diameter of the cylinder (0.5, 0.8, or 1.1 mm) was chosen based on the initial mechanosensitivity of the neuron. Response threshold was defined as the smallest force that triggered consistent (in three of three trials) firing in the recorded neuron. Once defined, mechanosensitivity was determined in response to threshold mechanical stimuli (100 ms rise time, 2 s width). Unlike in the previous study, we were unable to report effects on suprathreshold (presumably nociceptive) mechanical stimuli because the variability in response threshold is so large that in the absence of baseline values (i.e. suprathreshold response magnitude before treatment) for comparisons, the values of the post-treatment response magnitude could not be interpreted in a meaningful way.
Chemical stimuli were applied to the dura topically using a small piece of GelFoam® soaked with capsaicin or MO. The GelFoam was kept on the dura for 5 minutes, after which the receptive field was washed thoroughly with SIF. To determine responses to chemical stimulation, neuronal activity was recorded 30 minutes before and 60 minutes after removal of the GelFoam. The initial concentration of capsaicin was 1 μM. Neurons responding to this concentration were not tested for their responsiveness to higher concentrations. Neurons not responding to 1 μM were tested with 10 μM. Only those unresponsive to 10 μM were tested for 100 μM. Similarly, the initial concentration of MO was 2 mM. Neurons responding to this concentration were not tested for their responsiveness to higher concentrations. Neurons not responding to 2 mM were tested with 5 mM. Only those unresponsive to 5 mM were tested for 10 mM.
Order of experiments, blinding and data analysis
The study was performed in two phases. In the first phase, the person performing the electrophysiological recording was blind to the experimental group of the rats (sham, suture, or suture-plus-muscle injections). In the second phase, the experiments were done openly. Continuous data are presented as median with interquartile range (IQR) due to the deviation from the normal distribution assumption. Categorical data are presented as percentage. Categorical variables were compared using the chi-square test. Continuous variables were examined using the Kruskal-Wallis test for independent observations and Wilcoxon signed rank test for paired measurements. A neuron was counted as responsive to capsaicin or MO if its firing rate increased by >2 SD over baseline (pre-stimulus period). Two-tailed level alpha ≤0.05 was considered a threshold for statistical significance. All data analyses were performed by a biostatistician who was blinded to the different groups (control vs. suture BoNT-A vs. suture-plus-muscle BoNT-A) or rats and different stimuli (mechanical, capsaicin, MO).
Results
Intracranial mechanical sensitivity
Thresholds for mechanical activation of meningeal nociceptors from their intracranial dural receptive field were not affected by the two extracranial BoNT-A injection paradigms (Figure 2). For the C-units, median response thresholds were 0.80 (IQR 0.34 to 1.10) g in the control rats (n = 18), 0.60 (IQR 0.16 to 1.00) g in the suture group (n = 23), and 0.40 (IQR 0.07 to 0.60) g in the suture-plus-muscle group (n = 17) (Figure 2(c)). The two injection paradigm groups did not differ from the control group (p = 0.15). For the Aδ-units, the median response threshold was 0.60 (IQR 0.16 to 1.10) g in the control rats (n = 18), 0.16 (IQR 0.07 to 0.60) g in the suture group (n = 19), and 0.16 (IQR 0.07 to 1.00) g in the suture-plus-muscle group (n = 16). The two injection paradigm groups did not differ from the control group (p = 0.13). The scattergraphs in Figure 2(b), (c) (top panels) illustrate the distribution of response thresholds.
Intracranial responsiveness to TRPV1 activation by capsaicin
C-units response probability
Compared to the control group, the percentage of C-unit meningeal nociceptors activated by topical administration of capsaicin to the dura decreased after suture injections but not after the suture-plus-muscle injections (Figure 3(a)–(e)). In the control group, 67% (14/21) of the tested C-units were activated by capsaicin. In comparison, in the suture group, only 25% (5/20) of the C-units were activated (χ2 = 7.15; p = 0.007), whereas in the suture-plus-muscle group, 47% (9/19) of the C-units were activated (χ2 = 1.52; p = 0.217).
Effects of extracranial injections of BoNT-A on responsiveness of C-type meningeal nociceptors to stimulation of their intracranial dural receptive fields with the TRPV1 agonist capsaicin. (a)–(c) Mean firing rate of three C-type meningeal nociceptors sampled before (baseline/green) and after (red) application of capsaicin to rats that were untreated (a), or treated with BoNT-A injections into suture (b), or suture plus muscle (c). (d) Response probability. Compared to the control group, the percentage of C-unit meningeal nociceptors activated by capsaicin decreased significantly after suture (χ2 = 7.15; p = 0.0075) but not after suture-plus-muscle (χ2 = 1.52; p = 0.2) injections. (e) Cumulative response probability curves showing the cumulative percentage of neurons activated by each successively higher concentration. (f) Response magnitude. Boxplots illustrate median (thick horizontal line), interquartile range (25th–75th percentile; lower and upper box boundaries) and observations below and above the 25th and 75th percentile (whiskers) of neuronal firing rate before (gray boxes) and after (colored boxes) capsaicin administration to the dura in control, suture-injected, and suture-plus-muscle-injected rats. (g) Mean percent change calculated for the capsaicin-responsive units only (e.g. (a), (c)). (h) Response probability of Aδ-units. As indicated, Aδ-units are generally unresponsive to capsaicin. Of interest are the findings that suture injections blocked the capsaicin response more effectively than the suture-plus-muscle injections. BoNT-A: onabotulinumtoxinA.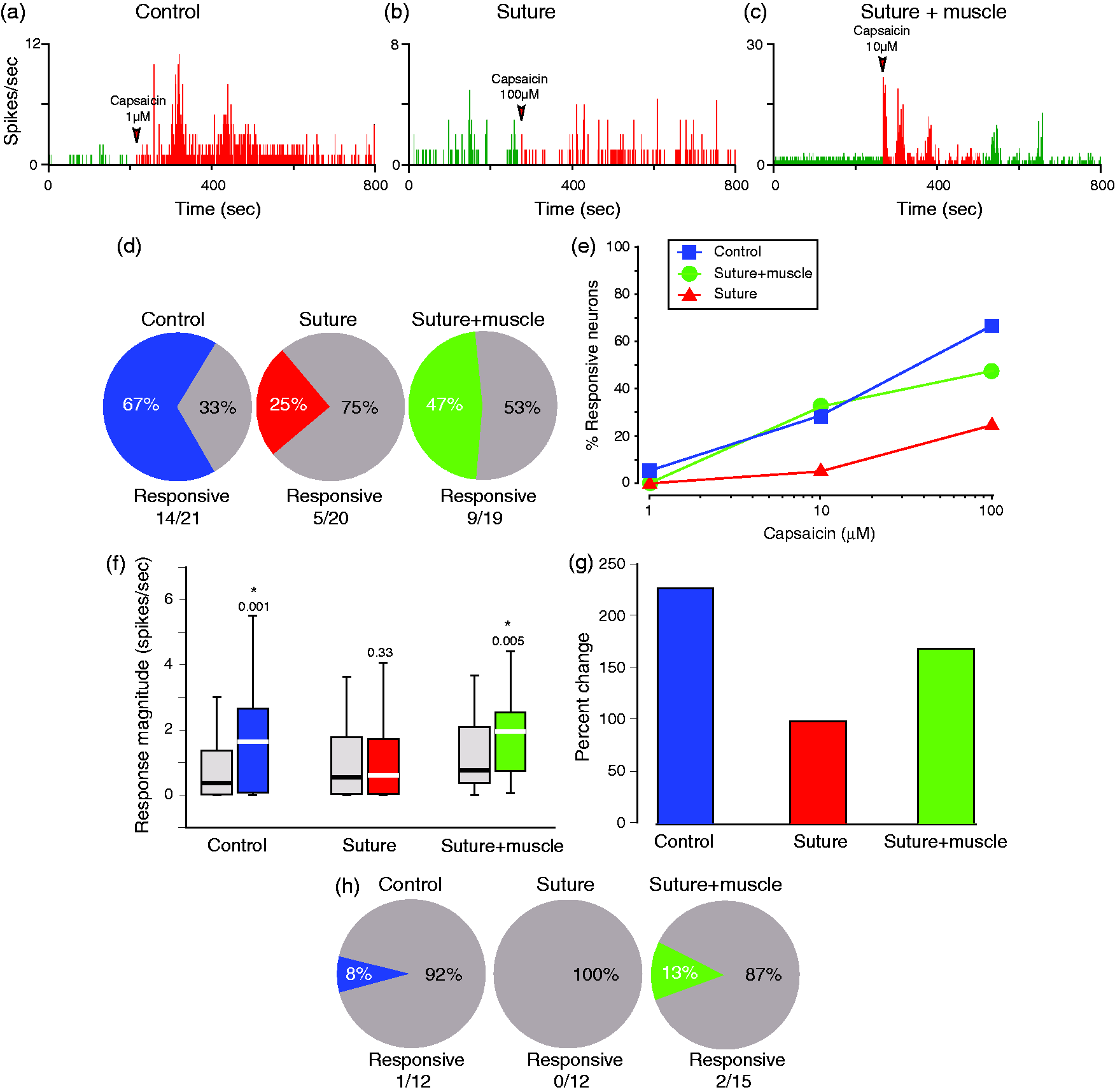
C-unit response magnitude
C-fiber meningeal nociceptor responses to capsaicin and mustard oil.

MO: mustard oil; IQR: interquartile range.
When analysis was based only on C-units that responded to capsaicin (14 in the control group, five in the suture group and nine in the suture-and-muscle group), the percent increase in neuronal firing was 227 for the control group, 98 for the suture group and 168 for the suture-plus-muscle group (Figure 3(g)). While the response magnitude in the capsaicin-responsive units appears lower after BoNT-A injections, small sample size precludes the utilization of formal statistical analysis.
Aδ-unit response probability
Aδ-meningeal nociceptors are usually not sensitive to capsaicin (23–26). Consistent with these reports, the incidence of activation by capsaicin was 8% (1/12) in the control group, 0% (0/12) in the suture group, and 13% (2/15) in the suture-plus-muscle group (Figure 3(g)).
Intracranial responsiveness to TRPA1 activation by MO
C-unit response probability
Compared with the control group, the percentage of C-unit meningeal nociceptors activated by topical administration of MO to the dura decreased significantly after suture as well as after the suture-plus-muscle injections (Figure 4(a)–(e)). In the control group, 67% (10/15) of the tested C-units were activated by MO. In comparison, in the suture and the suture-plus-muscle groups, 32% (9/28) of the units were activated by MO (χ2 = 4.72; p = 0.03). The two injection paradigms did not differ.
Effects of extracranial injections of BoNT-A on responsiveness of C-type meningeal nociceptors to stimulation of their intracranial dural receptive fields with the TRPA1 agonist mustard oil (MO). (a)–(c) Mean firing rate of three C-type meningeal nociceptors sampled before (baseline/green) and after (red) application of mustard oil to rats that were untreated (a), or treated with BoNT-A injections into suture (b), or suture plus muscle (c). (d) Response probability. Compared to the control group, the percentage of C-unit meningeal nociceptors activated by MO decreased significantly after suture as well as suture plus muscle injections (χ2 = 4.72; p = 0.03). (e) Cumulative response probability curves showing the cumulative percentage of neurons activated by each successively higher concentration. (f) Response magnitude. Boxplots illustrate median (thick horizontal line), interquartile range (25th–75th percentile; lower and upper box boundaries) and observations below and above the 25th and 75th percentile (whiskers) of neuronal firing rate before (gray boxes) and after (colored boxes) MO administration to the dura in control, suture-injected, and suture-plus-muscle-injected rats. (g) Mean percent change calculated for the MO-responsive units only (depicted in (a)). (h) Response probability of Aδ-units. The decrease in the number of units responding to MO (compared to control) was insignificant for both the suture (χ2 = 1.53; p = 0.2) and the suture plus muscle (χ2 = 0.89; p = 0.3) injection groups. BoNT-A: onabotulinumtoxinA.
C-unit response magnitude
BoNT-A inhibitory effects on C-unit responsiveness to MO was also reflected in the decrease in response magnitude (Figure 4(f)). In the control group, MO produced a large response of 500% above baseline firing (p = 0.008, including both responsive and unresponsive neurons). In contrast, there was no significant response to MO in either the suture group (p = 0.02) or the suture-plus-muscle group (p = 0.02) (Figure 4(f), Table 1). Here, as well as in the capsaicin experiments, we found no statistically significant differences in baseline firing rate between neurons recorded in control vs. treated rats.
When analysis was based only on C-units that responded to MO (10 in the control group, four in the suture group and five in the suture-plus-muscle group), the percent increase in neuronal firing was 293 for the control group, 118 for the suture group and 145 for the suture-plus-muscle group (Figure 4(g)). While the response magnitude in the MO-responsive units appears lower after BoNT-A injections, small sample size precludes the utilization of formal statistical analysis.
Aδ-unit response probability and magnitude
Aδ-units were generally less sensitive than C-units to MO. In the control group, 46% (6/13) of the tested Aδ-units (compared to the 67% of C-units) were sensitive to MO (Figure 4(h)). Neither suture nor suture-plus-muscle injections appear to alter the Aδ-unit responses to MO. In the suture group, 23% (3/13) of the tested units were activated by MO—an insignificant decrease compared to the probability of response in the control group (χ2 = 1.53; p = 0.22). In the suture-plus-muscle group, 29% (5/17) of the Aδ-units were activated by MO—again an insignificant decrease compared to the control group (χ2 = 0.89; p = 0.35).
Because only a small percentage of Aδ-units responded to MO, the response magnitude in the control group did not reach statistical significance (0.61 ± 0.21 spikes/sec at baseline vs. 1.14 ± 0.30 spikes/sec after MO, p = 0.064). Accordingly, changes in Aδ-units response magnitude to MO were not computed.
Discussion
This study is first to demonstrate that extracranial administration of BoNT-A inhibits responses of meningeal nociceptors to stimulation of their intracranial dural receptive fields. Most significant were the findings that extracranial administration of BoNT-A (i) inhibited responses of C-type meningeal nociceptors to stimulation of their intracranial dural receptive field with the TRPV1 agonist capsaicin, (ii) inhibited responses of C-type meningeal nociceptors to stimulation of their intracranial dural receptive field with the TRPA1 agonist MO, (iii) were more effective when the entire dose was injected along the cranial sutures than when it was divided into two portions (one-quarter of the dose injected near cranial sutures and three-quarters injected into muscles), (iv) required time that is longer than the several hours tested previously (27) for producing effective inhibition of intracranial dural receptive fields, (v) did not reduce threshold of intracranial mechanosensitivity in either C-meningeal nociceptors, and (vi) had no effect on Aδ meningeal nociceptors. In the context of chronic migraine, reduced sensitivity to molecules that activate meningeal nociceptors through the TRPV1 and TRPA1 channels can be important for BoNT-A’s ability to act as a prophylactic. Regarding treatment, superior effects of injections in which the entire dose is targeted to sutures (compared to injections that divided the dose between suture and muscle) raise the possibility that the current injection paradigm could be improved.
We interpret the findings as suggesting that injections of BoNT-A near extracranial nerve endings of suture branches of intracranial meningeal nociceptors may reduce the sensitivity of these nociceptors by preventing the docking of synaptic vesicles containing TRPV1 and TRPA1 receptors into the membrane of dural collateral branches of the same axon (27). Although we do not know how injections of BoNT-A in one area can affect the docking of synaptic vesicles in a collateral along the same axon, it is tempting to speculate that the light chain part of BoNT-A or the cleaved synaptosomal-associated protein of 25 kDa (SNAP-25) travels the very short distance across the calvarial bone—a hypothesis whose validation depends on the development of reliable tools (e.g. antibodies) to image/detect the intra-axonal location of the light chain alone, and on further elucidating intracellular mobility patterning of native and cleaved SNAP-25 (28,29). While raising the former possibility, we must keep in mind that reports of axonal transport of BoNT-A are primarily based on local administration of relatively large amounts of neurotoxin (135 pg per injection, or ∼450 pg/kg) (30). Lawrence and colleagues used rat sympathetic neurons culture to show that cleaved SNAP-25 can be detected in cell somas after administration of high (104 pM) but not low doses of BoNT-A (10 pM, ∼75 U) to their distal neurites, that on its own the cleaved SNAP-25 does not block the functional electrophysiologically recorded synaptic transmission in the cell somas, and that actual blockade requires doses greater than 106 pM (31). Alternatively, cleaved SNAP-25 has a dominant-negative effect on SNARE complex function (32), and only a single defective SNARE complex may be necessary to inhibit vesicular fusion (33,34). Consistent with Montecucco’s recent observations, SNAP-25 cleaved in the extracranial neuron could traverse a short distance within the fluid neuronal membrane or the cytosol over several days, entering a nearby collateral meningeal nociceptor and impair SNARE function.
Multiple studies implicate TRPV1 and TRPA1 channels in migraine pathophysiology (35–37). For TRPV1, clinical evidence for a possible role in migraine pathophysiology includes one genetic study showing a single nucleotide polymorphism in the TRPV1 gene in Spanish migraine patients (38), and reports of complete resolution (33%) or partial decrease in pain intensity (73%) four hours after intranasal administration of the TRPV1 agonist civamide (39), as well as 50%–80% improvement in chronic migraine after repeated intranasal administration of capsaicin (40)—both compounds recognized as capable of desensitizing the TRPV1 receptor. Preclinically, evidence for possible involvement in migraine includes anatomical studies showing presence of neuronal TRPV1 channels in dural pain fibers (41) and in peripheral trigeminovascular neurons (10); physiological studies showing that capsaicin administration to the dura produces TRPV1-dependent dilation of dural blood vessels (42), extracellular signal-regulated kinase (ERK) activation (i.e. sensitization) of trigeminal ganglion neurons (43), and robust activation of meningeal nociceptors (44); behavioral studies showing that activation of dural TRPV1 channels leads to decreases in facial and hind-paw withdrawal threshold (45); and pharmacological studies showing that systemic administration of SB-705498—a TRPV1 antagonist—decreases spinal trigeminal nucleus neurons’ responses to electrical and mechanical stimulation of the dura, as well as the sensitizing effect of inflammatory mediators (46). It is worth noting, however, that blockade of TRPV1 receptors was not effective in all experimental migraine models (47).
Regarding TRPA1 channels, clinical hints for a possible role in promoting migraine include reports that exogenous triggers of migraine such as cigarette smoke, formaldehyde and umbellulone (the active ingredient in the “headache tree” Umbellularia californica) are also TRPA1 activators (48). Preclinically, evidence for potential involvement in migraine include anatomical studies showing expression of TRPA1 channels in trigeminovascular neurons located in the trigeminal ganglion (10), physiological studies showing existence of TRPA1 current in meningeal nociceptors (49), behavioral studies showing that MO produces cephalic and extracephalic allodynia as well as decreased exploratory behavior (49), and pharmacological studies showing that activation of TRPA1 channels by intranasal umbellulone stimulates meningeal nociceptors and causes CGRP release and meningeal vasodilation (48).
The plethora of evidence for multiple mechanisms by which activation of TRPV1 and TRPA1 channels can lead to activation of meningeal nociceptors and the initiation of headache supports the ongoing effort to target these channels for the treatment of migraine. Until now, two mechanistically different approaches have been tried. The first attempted to block activation of these channels using antagonists (46,49,50) whereas the second attempted to reduce their sensitivity using desensitizing agonists (39,40,51–55). The current study raises yet another possibility, namely that effort to prevent activation of meningeal TRPV1 and TRPA1 channels may be achieved indirectly by using a neurotoxin to block their insertion into the synaptic membrane of the receptor. Theoretically, an advantage of this approach may be its minimal unwanted potential systemic adverse effects attributed to a restricted peripheral and local site of action.
Regarding mechanosensitivity, it may appear that the findings that extracranial injections of BoNT-A did not reduce mechanical response threshold of the C-fibers meningeal nociceptors is inconsistent with our previous report (Burstein et al., Cephalalgia 2014 (8)), in which we showed that BoNT-A inhibits c-fibers’ mechanosensitivity. However, a closer look at the results of our previous study indicate that BoNT-A inhibited the responsiveness to suprathreshold but not to threshold mechanical stimuli and accordingly concluded that BoNT-A selectively blocks insertion of membrane channel involved in the transduction of mechanical nociception but not the transduction of tactile mechanical stimuli (8,56). In the current study, we sought to determine whether BoNT-A’s lack of effects on the response threshold of c-fibers was due to the short exposure time rather than its true inability to reduce threshold mechanosensitivity. Our results show that BoNT-A’s inability to reduce the mechanical response threshold is independent of the site of administration (directly on the dural receptive field vs. extracranially) or the exposure time (two to four hours in the first study vs. seven days in the current study). In retrospect, these findings are consistent with the human experience that subcutaneous injections of BoNT-A do not produce local numbness or loss of tactile sensation.
Finally, the therapeutic applicability of the findings that suture injections are more effective than suture-plus-muscle injections must be considered with caution, however, as this observation was limited to C-type meningeal nociceptor responses to stimulation of their dural receptive field with TRPV1 but not TRPA1 agonists. Mechanistically, the theoretical advantage of injecting BoNT-A along suture lines is supported by the existence of collateral branches of parent axons of intracranial meningeal nociceptors that cross the calvarial sutures from inside to outside the head (5–7), thus establishing an axonal network along which nociceptive signals that originate in the cortex (e.g. cortical spreading depression, seizure) can influence the molecular environment in both the intracranial pia and dura (57) and the extracranial calvarial periosteum (8), and vice versa, nociceptive signals that originate in periosteal nociceptors (58) can influence the molecular environment in the dura. The currently approved injection paradigm is directed to several muscle groups adjacent to sutures, in addition to other muscle regions that are innervated by relevant nociceptors. Nevertheless, because the therapeutic significance of injecting the suture vs the muscles is not known, we can only speculate that the current study raises the possibility that adding the suture as one more injection site may be beneficial. We anticipate that this approach would be technically challenging and possible only if aided by outpatient specialized peripheral imaging, such as the use of ultrasound.
Key findings
This study is the first to show that extracranial administration of botulinum toxin A inhibits responses of C-type meningeal nociceptors to stimulation of their intracranial dural receptive fields. Extracranial administration of botulinum toxin A suppresses C-type meningeal nociceptors’ responses to stimulation of their intracranial dural receptive field with TRPV1 and TRPA1 agonists. Reduced sensitivity to molecules that activate meningeal nociceptors through the TRPV1 and TRPA1 channels can be important for botulinum toxin A’s ability to act as a prophylactic. Suppression of intracranial receptive fields following extracranial administration is observed after seven days—potentially explaining the delayed therapeutic effect in patients. Injections along suture lines may be more effective than muscle injections in suppressing responses to stimulation of meningeal nociceptors through TRPV1 and TRPA1 channels.
Footnotes
Declaration of conflicting interests
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Dr Burstein received research funds from Allergan. He is also a consultant to Allergan. Dr Brin is an employee of Allergan plc and receives stock. Drs Zhang, Strassman and Novak have nothing to declare.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: National Institutes of Health (NIH) grants NS-069847 and NS-079678 (RB) and a grant from Allergan Inc (Irvine, CA).