
Abstract
Background:
Understanding leg stiffness (Kleg) in amputee sprinters is important for the evaluation of their sprint ability and development of running-specific prostheses (RSP).
Objectives:
To investigate Kleg during hopping in amputee sprinters.
Study Design:
Cross-sectional study.
Methods:
Seven transtibial (TT) and seven transfemoral (TF) amputee sprinters, as well as seven non-active able-bodied subjects, performed one-legged hopping matching metronome beats at 2.2 Hz. Amputees hopped on their sound limb whereas able-bodied (AB) subjects hopped on their dominant limb. Using a spring-mass model, Kleg was calculated from the subjects’ body mass, ground contact and flight times.
Results:
Both TT and TF sprinters demonstrated significantly higher Kleg than AB subjects. Kleg during hopping on the sound leg significantly correlated with personal records attained in a 100-m sprint in both TT (r = −0.757) and TF sprinters (r = −0.855).
Conclusion:
The results of the present study suggest that amputee sprinters have a greater Kleg during hopping than inactive non-amputees, and that their sprint ability can be predicted from the Kleg during hopping at 2.2 Hz on the sound limb.
Clinical relevance
Exercise challenges need to be taken into consideration when planning routine training regimens for amputee sprinters. Furthermore, increased understanding of Kleg in amputee sprinters is relevant to evaluate their sprint ability and develop running-specific prostheses.
Background
During hopping, jumping and running, our legs exhibit characteristics similar to those of a spring. 1 Thus, in these movements, the musculoskeletal structure of the legs is often modelled with a spring-mass model, which consists of a body mass and a linear leg spring supporting the body mass.1,2 The leg spring is compressed during the first half of the stance phase and rebounds during the second. The stiffness of the leg spring or leg stiffness (Kleg, defined as the ratio of maximal ground reaction force to maximum leg compression at the middle of the stance phase) has been shown to increase with hopping height at a given hopping frequency.3-5 Further, some cross-sectional studies suggest that endurance training enhances Kleg,6,7 but that plyometric training has a stronger influence on Kleg than endurance training.8-11 Moreover, Kleg during hopping has been shown to correlate with sprint ability in sprinters and in handball and tennis players.12-14 Thus, identifying Kleg and its relation to sprint performance in specific populations would be helpful for the development of more effective training methods.
Recent studies suggest that analyzing Kleg in amputee sprinters would be useful in evaluating their sprint ability and develop sprint-dedicated prostheses.15,16 However, little is known about Kleg and its relationship to sprint ability in amputee sprinters. Therefore, the purpose of this study was to investigate Kleg in amputee sprinters. For this, we compared Kleg of amputee sprinters and inactive non-amputees in a cross-sectional study based on the analysis of one-legged hopping, which facilitates quantification of the components of the spring-mass model.
Overall, leg stiffness partly depends on the combination of the individual stiffness of muscles and tendons, 2 which is an important factor influencing force production in human movements. Some studies demonstrated that during isokinetic contraction and running at constant speed the sound limb of active amputees has stronger force-producing capacity than the dominant limb of non-amputees.17,18 Furthermore, the fastest non-amputee sprinters are those who can apply greater forces to the ground. 19 Similarly, Grabowski and colleagues 20 found that in unilateral transtibial (TT) amputees the average vertical ground reaction forces during the stance phase of running were greater in the sound leg than in the prosthetic leg, across a range of speeds including top speed. Thus, we hypothesized that:
amputee sprinters would show greater Kleg during hopping than inactive non-amputees
Kleg during hopping would correlate with sprint ability in amputee sprinters.
Methods
Participants
The study participants comprised of 14 subjects with either unilateral TT (five male and two female subjects) or unilateral transfemoral (TF) amputation (four male and three female subjects) and no vascular disease or skin abnormalities in their residual legs. All the participants were sprinters on a track and field team and had competed at recreational, regional, national and international levels within the preceding year. Their personal recorded times in the official 100-m race (100-m PR) ranged between 12.43 and 20.43 s for TT amputees and between 17.62 and 24.08 s for TF amputees. Their physical characteristics are shown in Table 1. Seven able-bodied subjects (AB subjects; five male and two female subjects) also participated in this study as a control group. All AB subjects were either sedentary or mildly active but none had been involved in any type of regular exercise or training for at least one year prior to the test. The protocol was approved by the local ethical committee and complied with the guidelines set out in the Declaration of Helsinki (1983).
Characteristics of the study participants.
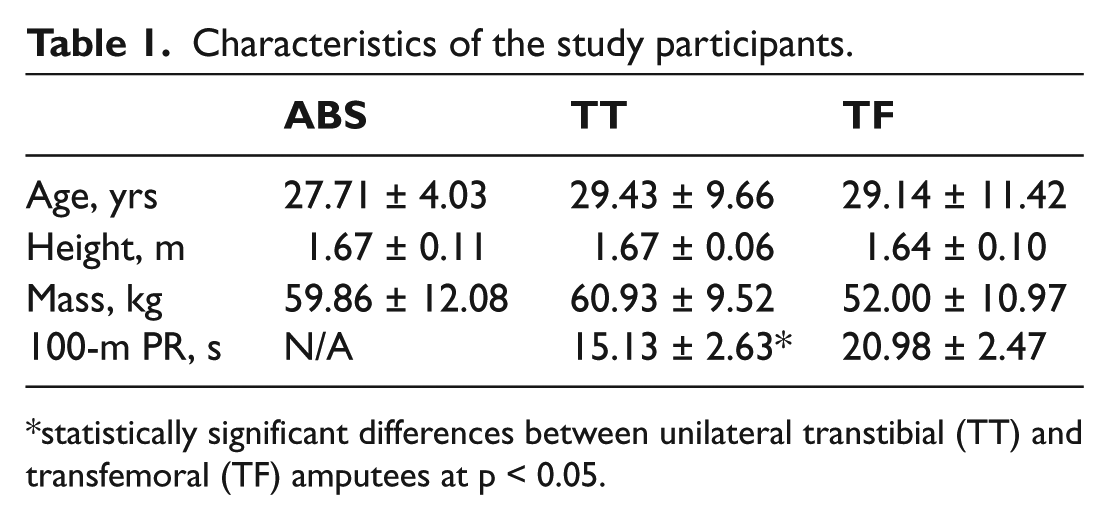
statistically significant differences between unilateral transtibial (TT) and transfemoral (TF) amputees at p < 0.05.
Task and procedure
The amputees’ task consisted of doing seven successive hops on their sound leg. During hopping the subjects kept their hands on their hips. The task of AB subjects was to hop the same number of times on their dominant leg. Prior to the experiment, all AB subjects used ball kicking to determine their dominant leg; they were asked which leg they preferred to kick a ball with, and the limb used to kick the ball was identified as the dominant leg. 21 Further, five TT were instructed to hop not only on the sound leg, but also on their prosthetic leg. They wore running-specific prostheses (RSP) made of carbon fibre. One TT sprinter wore Cheetah (Össur, Reykjavík, Iceland) and four TT sprinters wore KATANA (IMASEN Engineering Corporation, Kakamigahara, Japan). The hopping mechanics were then compared between sound and prosthetic legs.
The subjects were instructed to match the timing of landings with audible clicks generated by a metronome (2.2 Hz; preferred hopping frequency in humans3,5). Because the contact time can affect stiffness regulation during hopping at a given frequency, 22 the participants were asked to hop with as short a contact time as possible. Before data collection, all participants were instructed to practise for as long as they needed until they felt comfortable with the task. The practice period was three to four minutes in length. According to the participants’ subjective impression, this session was sufficient to get used to the task.
Data collection and analysis
All subjects completed seven continuous hops and all hops were used in the analysis. Leg stiffness (Kleg) during the contact phase was calculated with a spring-mass model, which utilizes the body mass and a single leg spring supporting the body mass (Figure 1). During hopping, the peaks of vertical ground reaction force (Fmax) and leg compression (Δyc) coincide in the middle of the ground contact phase. At this point, Kleg can be calculated as the ratio of Fmax to Δyc. According to Dalleau, 23 Fmax during hopping can be calculated through the following equation
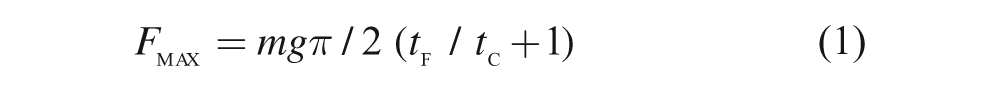
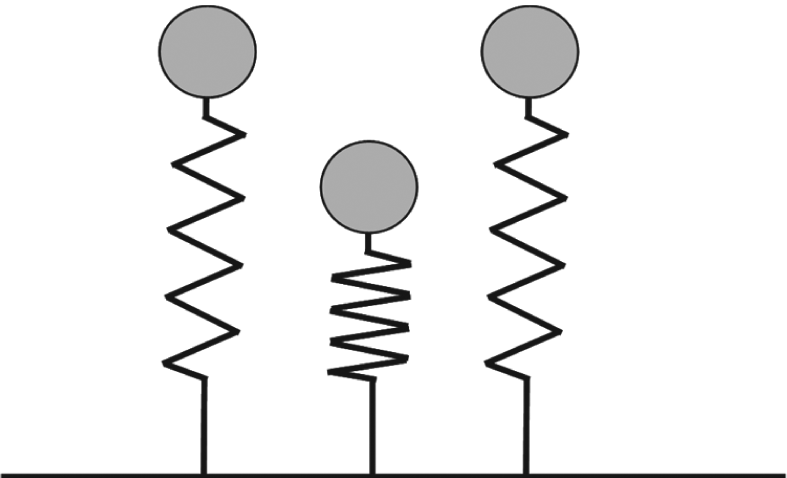
Spring-mass model for hopping. This model consists of a body mass and a massless linear spring supporting the body mass. Mass is equivalent to body mass. The model is shown at the beginning of ground contact phase (left), the middle of ground contact phase (middle), and at the end of ground contact phase (right). Adapted from Blickhan 1 .
where m is the total body mass (in kg), g the standardized acceleration due to gravity (9.81 m/s2), t c the ground contact time, and t f the flight time (both in s). Δyc was then calculated using the following equation:
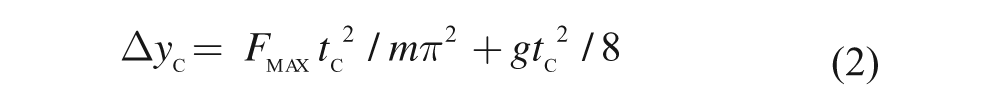
Using expressions (1) and (2), Kleg can be calculated through the following equation 23 :
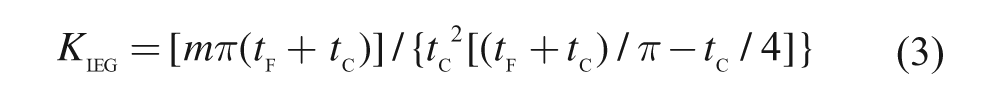
In this study, we used a triaxial accelerometer (Free-Jump; SENSORIZE Inc., Roma, Italy), sampled at 100 Hz, to determine tc and tf. Because body size influences the stiffness value, 24 Kleg was divided by the subject’s body mass.
Statistical analysis
Before the results were interpreted, Shapiro-Wilks and Levene’s tests were performed to ensure that the assumptions of normality and homogeneity of variance were met. These tests revealed that our groups are homogenous. One-way ANOVA and post hoc multiple comparison tests using Fisher’s protected least significant difference (PLSD) were performed to compare physical characteristics and spring-mass parameters among the three groups. All data are presented as mean values with standard deviation (SD). In addition, the t-test for independent samples was used to compare 100-m PR between TT and TF amputees. Similarly, the t-test for independent samples was used to compare Kleg between sound and prosthetic legs for five TT. The Pearson’s correlation coefficient was used to examine the relationship between Kleg and 100-m PR in TT and TF amputees. The slopes of the regression lines of 100-m PR on Kleg obtained for the groups of amputees were compared by a t-test. The significance level was set at p < 0.05 for all tests performed. All the statistical analyses were performed with the package SPSS for Windows (Version 11.0.1J; SPSS Inc., Chicago, IL, USA).
Results
General characteristics of the three groups of participants are presented in Table 1. No statistically significant differences in age, height and body mass were found among the groups.
As shown in Figure 2, Kleg was significantly greater in TT and TF amputees than in the AB subjects. However, there were no significant differences in Kleg between the TT and TF groups.
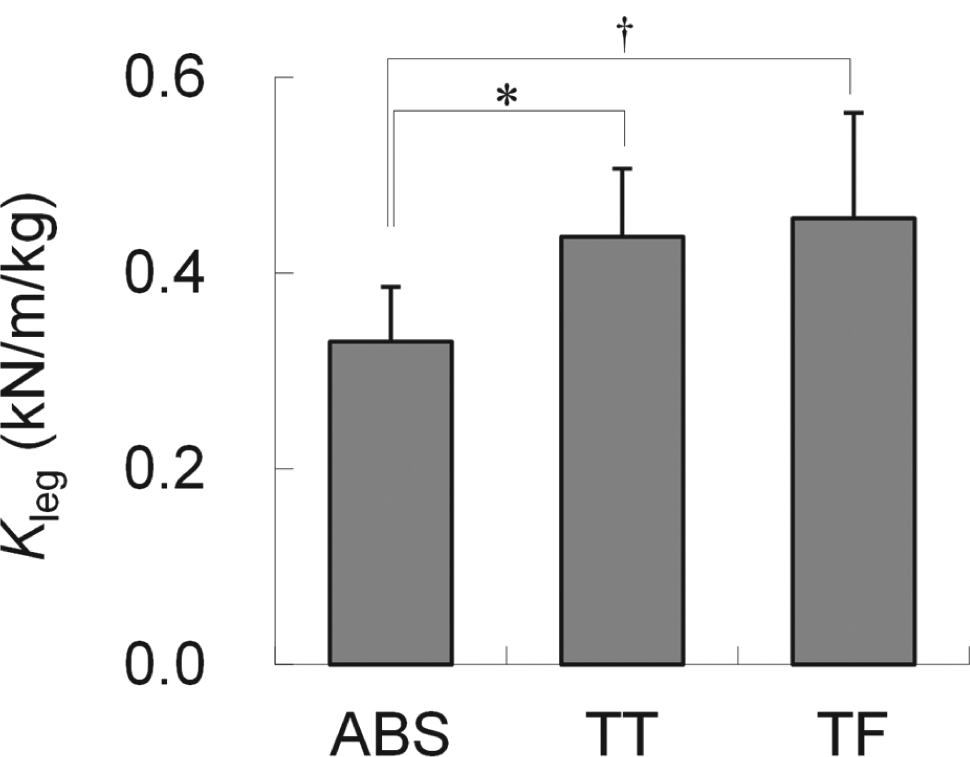
Mean leg stiffness (Kleg), and corresponding standard deviation, of able-bodied (AB) subjects and unilateral transtibial (TT) and transfemoral (TF) amputees. (†) and (*) indicate statistically significant differences between 2 groups at p < 0.01 and p < 0.05, respectively.
Figure 3 shows comparison of Kleg between sound and prosthetic legs for five TT. On average, there was no significant difference in Kleg between the legs. (0.46 ± 0.07 for sound leg and 0.43 ± 0.10 for prosthetic leg).
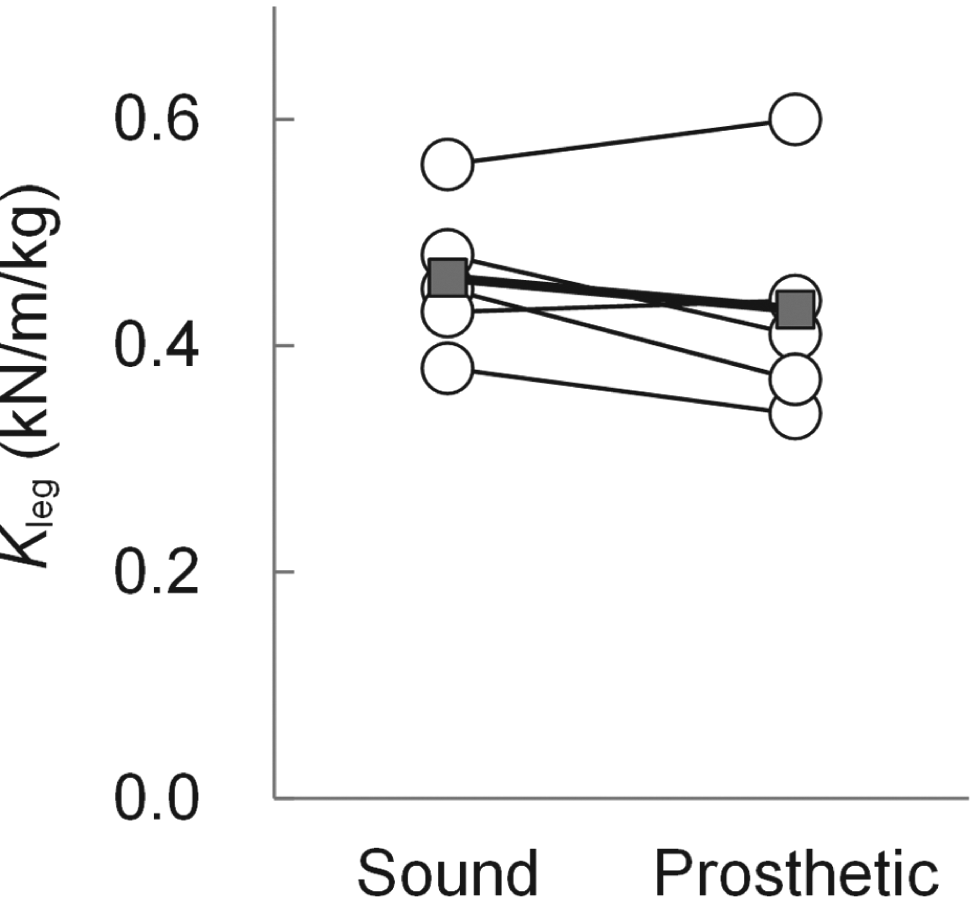
Comparison of Kleg for sound leg (SND) and prosthetic leg (PST) in TT sprinters (n=5). Unfilled circles represent an individual value for each subject. Filled squares represent the average value for the 5 subjects.
Comparison of hopping frequency, tc, and tf among the three groups is shown in Table 2. Although no statistically significant differences among groups were found in actual hopping frequency, tc was significantly shorter in the TT and TF groups than in the AB group. Similarly, tf was significantly longer in TT amputees than in the AB subjects. No significant differences between the TF group and the AB or TT groups could be depicted.
Comparison of temporal parameters among the able-bodied (AB) subjects and unilateral transtibial (TT) and transfemoral (TF) amputees.
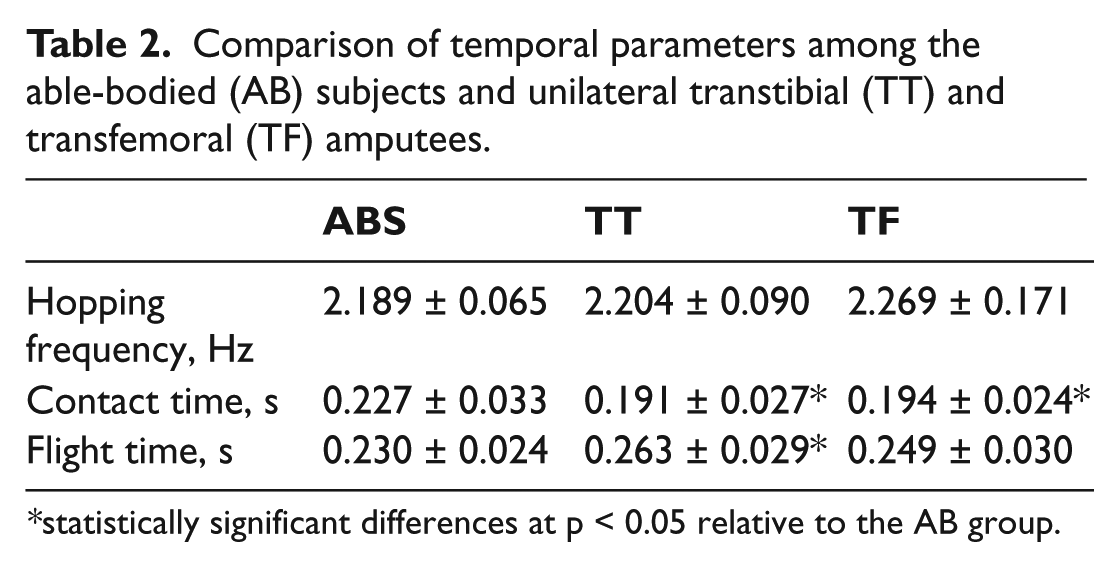
statistically significant differences at p < 0.05 relative to the AB group.
Figure 4 shows the relationship between Kleg and 100-m PR in TT and TF amputees. Statistically significant negative correlations were found between Kleg during hopping on the sound leg and 100-m PR of TT (r = -0.757, p < 0.05) and TF (r = -0.855, p < 0.01) amputees. In addition, no significant difference was found between the slopes of the regression lines obtained for these two groups. Furthermore, although there were only five transtibial amputee sprinters, we found that the regression coefficient was -0.755.
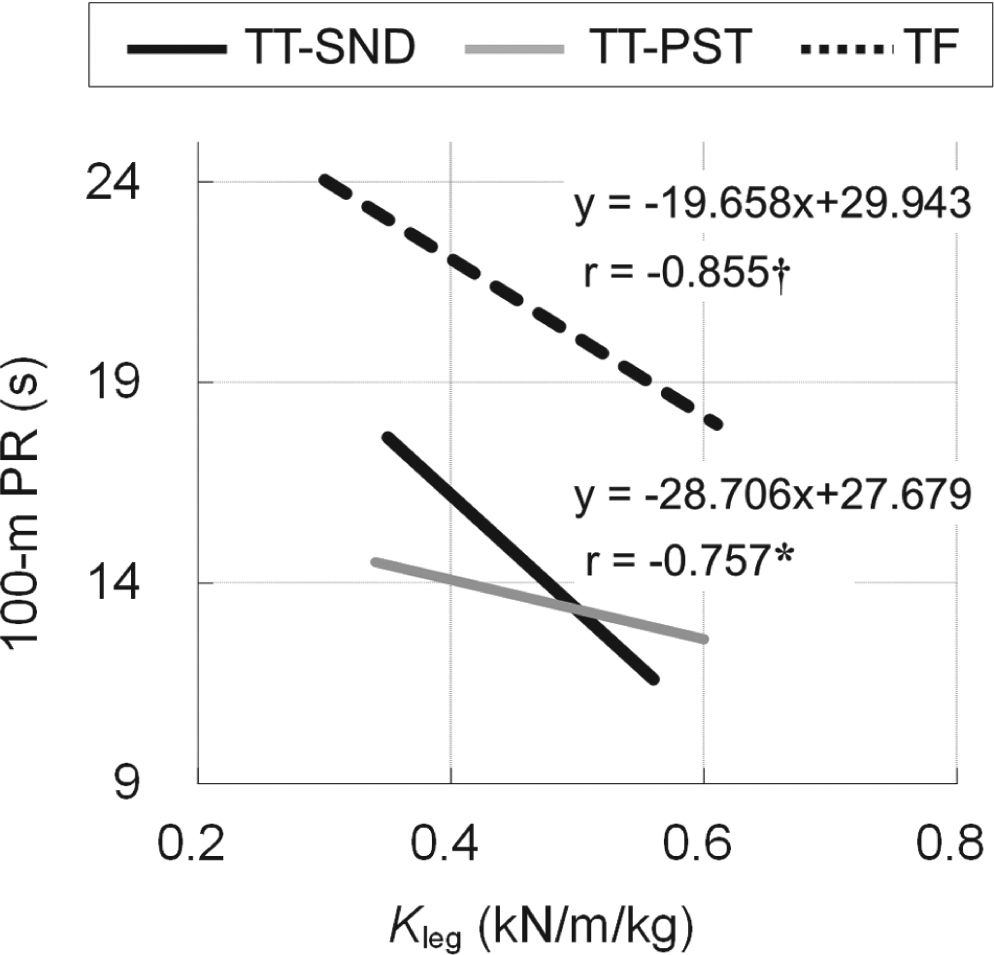
Regression of personal records attained in a 100-m sprint (100-m PR) on leg stiffness in unilateral transtibial (TT) and transfemoral (TF) amputation groups. (†) and (*) indicate statistical significance at p < 0.01 and p < 0.05, respectively. Although there were only five transtibial amputee sprinters, the regression coefficient was -0.755.
Discussion
The purpose of this study was to investigate Kleg in amputee sprinters. We compared Kleg during one-legged hopping of amputee sprinters and inactive non-amputees in a cross-sectional manner. The TT and TF groups showed higher Kleg than the AB group. In addition, both amputee groups exhibited shorter tc than the AB group; the TT group also showed longer tf than the other two groups. The results thus indicate that at a given frequency amputee subjects hopped higher than AB subjects. To our knowledge, this is the first study examining Kleg during hopping in amputee sprinters. The results presented herein support our first hypothesis stating that the amputee sprinters would show greater Kleg during hopping than inactive non-amputees.
A Kleg of 0.71 kN/m/kg (SD 0.19) was reported recently during hopping at 2.2 Hz in well-trained male sprinters, long jumpers or long distance runners belonging to a university track and field team. 26 This is greater than the values obtained in the present study (TT, 0.44 kN/m/kg (SD 0.07); TF, 0.46 kN/m/kg (SD 0.11)). Although the previous study utilized double-legged hopping, the current study calculated Kleg using one-legged hopping. A past finding demonstrated that the Kleg during one-legged hopping at 2.2 Hz was on average 34.6% less than the Kleg in two-legged hopping. 27 When we convert the previous study’s data (double-legged hopping) 26 to one-legged hopping, the Kleg would be approximately 0.46. This value is similar to our data, indicating that our TT and TF groups would have a similar level of Kleg in their sound leg to the able-bodied sprinters.
Determinants of the differences in Kleg during hopping found between the two groups of amputees and between amputees and AB subjects are still unknown. Leg stiffness is known to depend in part on the combination of individual stiffness values of muscles and tendons. 2 Sherk et al. 28 reported, however, that muscle cross-sectional areas of the intact lower extremities of TT and TF amputees were similar to those of age- and sex-matched non-amputee control groups. On the other hand, pre-activity (muscle activity before ground contact), muscle activity including the short-latency stretch reflex response at landing, 3 and antagonistic muscle co-contraction 29 also regulate Kleg. Accordingly, Chen et al. 30 compared lower extremity amputees and control subjects by measuring the thresholds for transcranial magnetic stimulation to determine intracortical inhibition and facilitation of the intact leg (quadriceps femoris). They found no significant differences between the two groups in the inhibitory and facilitatory cortical circuitry controlling the intact leg. 30 Given this, it can be hypothesized that the contrasting Kleg values might be due to differences in stiffness of tendinous tissues. This hypothesis is supported by previous findings indicating that the differences in Kleg between power-trained athletes and distance runners may be attributed to an intrinsic stiffness of the tendons and/or aponeuroses. 9 Future studies should focus on investigating the determinants of differences in Kleg during hopping among TT, TF and AB subjects.
Despite the fact that no significant differences were found in Kleg between the TT and TF groups, 100-m PR was significantly lower in TT than in TF sprinters (Table 1). Consequently, regression lines of 100-m PR on Kleg obtained for the TT and TF groups show different elevations but similar slopes. The present results indicate that sprint ability in both the TT and TF groups can be predicted from Kleg during hopping at 2.2 Hz on the sound limb. Nevertheless, the relationship between Kleg and sprint ability was not comparable between different amputation levels, probably due to whether the prosthetic leg included the knee unit and not to differences in the sound leg.
So far, no study has yet quantified the Kleg during hopping on a prosthetic leg. As shown in Figure 3, there was no significant difference in Kleg between the legs in TT sprinters. Further, there were linear relationships between the Kleg and 100-m PR in both sound and prosthetic legs in TT sprinters (Figure 4). These results suggest that both sound and prosthetic legs of TT sprinters would contribute to sprint performance in the same manner as able-bodied athletes.12-14 In other words, sprint performance in amputee sprinters is not necessarily dependent only on physical property of RSP, which is made of carbon fibre. Since there were also linear relationships between the Kleg of sound limb and 100-m PR for TF sprinters (Figure 4), we conclude that utilizing the Kleg in sound leg is valid in predicting sprint performance in amputee sprinters.
Several limitations of the study may hinder the interpretation of the findings. First, this study is limited in terms of the number of subjects included. In fact, although there was no significant difference in Kleg between the legs in TT sprinters, two of five amputees showed a slightly higher Kleg in the prosthetic leg than the sound leg (Figure 3). In addition, there was a linear trend for the relationship between the Kleg and sprint ability in the prosthetic legs of five amputees (Figure 4). This result indicates the possibility that the amputees’ sprint ability may be predicted from Kleg during hopping on the prosthetic limb, at 2.2 Hz, rather than during hopping on the sound limb. Caution should therefore be used in the interpretation and generalization of these findings to the population of TT and/or TF sprinters. Secondly, we compared the Kleg among TT, TF amputee sprinters and non-active able-bodied subjects. However, results acquired from the amputee sprinters should be compared to a matched group of non-amputee sprinters of similar ability. Thirdly, our results were based on cross-sectional, not longitudinal, observations. Therefore, the current data cannot determine whether the higher Kleg in the amputee sprinters was due to training or had a genetic component. Clearly, additional work is necessary for a deeper characterization of sprint ability in amputee sprinters.
Conclusion
The results of the present study suggest that amputee sprinters have stiffer leg springs than do inactive non-amputees, as evaluated in a one-legged hopping task. Statistically significant correlations were depicted between Kleg during hopping on the sound leg and personal recorded times attained in a 100-m sprint for both TT and TF subjects, indicating that amputees’ sprint ability can be predicted from Kleg during hopping.
Footnotes
Funding
This study was supported by a Grant-in-Aid for Research from Mizuno Sports Promotion Foundation 2011.
Conflict of interest
None of the authors has any conflict of interest associated with this study.