
Abstract
Erethizon dorsatum papillomavirus 1 (EdPV1) and Erethizon dorsatum papillomavirus 2 (EdPV2) are associated with cutaneous papillomas in North American porcupines (Erethizon dorsatum). This study defined gross, histopathologic, and molecular characteristics of viral papillomas in 10 North American porcupines submitted to the New York State Animal Health Diagnostic Center. Investigation for the presence of EdPV1 and EdPV2 DNA via polymerase chain reaction (PCR) was performed in 9 of the 10 (90.0%) porcupines, and all porcupines were investigated for the detection and localization of EdPV1 and EdPV2 E6 and E7 nucleic acid via chromogenic in situ hybridization (CISH). Next-generation sequencing (NGS) was performed in 2 porcupines. Papillomas were diagnosed on the muzzle (n = 4), caudal dorsum (n = 1), upper lip (n = 1), chin (n = 1), gingiva (n = 2), and nasal planum (n = 1). Histologically, the lesions consisted of hyperplastic epidermis or epithelium with orthokeratotic keratin, prominent keratohyalin granules, and intranuclear inclusion bodies. PCR identified EdPV1 in 6 of 9 samples and EdPV2 in the remaining 3 samples. NGS resulted in 100% genome coverage of EdPV1 and 76.20% genome coverage of EdPV2 compared with GenBank reference sequences, with 99.8% sequence identity to the complete EdPV2 L1 gene of a novel subtype recently identified in France. Hybridization patterns in 9 of the 10 (90.0%) porcupines were characterized by strong nuclear signals in the superficial epidermis, with strong nuclear and punctate cytoplasmic signals in the stratum spinosum and basale. In one animal, CISH suggested dual EdPV1 and EdPV2 infection.
Keywords
Papillomaviridae is a large family of viruses that causes disease in a diverse range of animal hosts. These are nonenveloped, circular, double-stranded DNA viruses with relatively small genomes that are typically close to 8 kilobases. 6 Infection occurs at the stratified squamous epithelium of skin or mucosa, inducing epithelial proliferations such as papillomas, fibropapillomas, and malignant neoplasia.19,29 Most papillomaviruses are highly host-specific and often site-specific, with the notable exception of the delta papillomaviruses, which are capable of tumor induction in distantly related hosts.11,15 Transmission occurs through direct and indirect contact, with virus entry through breaks in the skin or mucosa and subsequent infection of the basal epithelium. 12
The North American porcupine (Erethizon dorsatum) is an arboreal rodent within the New World porcupine family Erethizontidae and is widely distributed throughout North America. They are facultative specialist herbivores capable of adapting to harsh climates and low-quality diets, which allows them to survive across a wide range of habitats. 4 This species was listed as “least concern” by the International Union for Conservation of Nature Red List in 2016, with densities of up to 12 individuals per square kilometer; however, hunting, collisions with cars, and loss of suitable habitat are recognized threats. 7 Porcupines support ecosystem health by altering tree structure and composition, which creates habitats and food sources for other species. 2 They additionally act as important prey items for the fisher (Pekania pennanti) where their habitats overlap. 2
Papillomavirus infection has been reported predominantly in mammalian and avian hosts, including several species of rodents such as mice, rats, and hamsters. 22 Most reports of papillomaviruses in the veterinary literature are focused on domestic animals, with a more limited understanding of those affecting free-ranging wildlife. Despite this, wildlife has been postulated as a reservoir for papillomavirus infection, particularly for the delta papillomaviruses, which are capable of infecting multiple host species. 20 Thus, wildlife may play an important role in the pathogenesis of papillomavirus infection at the interface of animals managed under human care. A subset of the reported papillomavirus infections of wildlife are additionally associated with tumor development, with a higher incidence of fibropapillomas over carcinomas.8,10,23 Fewer recognized wildlife hosts across a diverse taxonomic range suggests the presence of undiscovered papillomaviruses in this group. 10
Two papillomaviruses have been identified in the North American porcupine. Erethizon dorsatum papillomavirus 1 (EdPV1) was first detected from a captive North American porcupine in New York. 18 This animal presented with multiple lobulated and sessile to exophytic masses along the foot pads and facial skin. A captive North American porcupine in Arizona with multiple exophytic papules, nodules, and plaques on the muzzle, pinnae, palpebrae, and periocular skin was found to be infected with Erethizon dorsatum papillomavirus 2 (EdPV2). 25 More recently, a captive North American porcupine in France was identified with EdPV1 and EdPV2 coinfection, with a novel lineage of EdPV2 with 93.33% whole-genome sequence identity to the previously published sequence in GenBank (MH376689). 24 Virus-induced lesions were identified in all porcupines and included prominent keratohyalin granules and koilocytes, in addition to epidermal hyperplasia with orthokeratotic to parakeratotic hyperkeratosis.18,21,24 EdPV2 infection was additionally associated with intranuclear amphophilic and glassy material suggestive of viral inclusions within keratinocytes in the stratum spinosum and granulosum. 21
Despite infection in the same host species, EdPV1 and EdPV2 are highly divergent. EdPV1 is the sole member of the Sigmapapillomavirus genus and in a monophyletic clade with the human papillomavirus genus Nupapillomavirus. EdPV2 was found to cluster nearest to members of the rodent genus Pipapillomavirus; however, given its 59.97% nucleotide identity to the closest related virus (Rattus norvegicus papillomavirus 1), EdPV2 is thought to represent a novel genus within the family. 25
As reports characterizing EdPV1 and EdPV2 infection are limited to 3 captive animals, the prevalence of these viruses in both captive and wild porcupine populations is unknown. This study describes the morphologic and molecular features of EdPV1 and EdPV2 infection in captive and wild North American porcupines and outlines diagnostic methods to detect and differentiate these infections.
Materials and Methods
Study Population and Histopathology
North American porcupine biopsy (n = 3) and necropsy (n = 7) submissions from 10 individuals were evaluated at the Cornell University College of Veterinary Medicine Animal Health Diagnostic Center between November 2019 and November 2021. Skin was sampled, fixed in 10% neutral-buffered formalin, and processed routinely. From all cases, 5-μm sections were cut, mounted on glass slides, and stained with hematoxylin and eosin. In individuals with multiple papillomas, a single lesion was evaluated.
Polymerase Chain Reaction and Sanger Sequencing
Polymerase chain reaction (PCR) tests were performed by the Virology Laboratory at the Animal Health Diagnostic Center following protocols and procedures established in the laboratory. Commercial kits were used to extract DNA from fresh skin (QIAamp Viral RNA Mini Kit, QIAGEN, Germantown, MD, USA) or formalin-fixed paraffin-embedded (FFPE) skin (RecoverAll Total Nucleic Isolation Kit, Invitrogen, Carlsbad, CA, USA), and blank extractions were used as negative controls. Samples were considered positive if target viral DNA was extracted and amplified. After the initial positive samples were identified, a previously characterized sample was used in subsequent reactions as a positive control. The extracted DNA was amplified via the SuperScript III One-Step RT-PCR System (Invitrogen, Carlsbad, CA, USA) for EdPV1 and EdPV2 in parallel to using primers designed to target the E1 gene. Primers for EdPV1 including EdPV11382F (5’-GTGATAAAACTATGTCTGCAAATTGGGTA-3’) and EdPV1-1775R (5’-CATTGCACCATTTTTGTGAGTTCA-3’) were designed based on available reference sequence (GenBank: AY684126). Primers for EdPV2 including EdPV2-1313F (5’-CAAGACATGTGGTCAAAGCTGGGTA-3’) and EdPV2-1714R (5’-CACTGCACCATTTTTGAAAGCTCA-3’) were additionally designed based on the reference sequence (GenBank: MH376689). PCR amplicons from 2 cases were resolved in 1.5% agarose gels, purified using a commercial kit (QIAquick PCR Purification Kit, QIAGEN, Germantown, MD, USA), and subsequently sequenced via the Sanger method and analyzed with an automated DNA sequencer.
Next-Generation Sequencing
Sequence-independent, single-primer amplification (SISPA) was used for random amplification and sequencing of nucleic acids in the FFPE extraction product, modified from a previously reported protocol. 3 Briefly, 11 μL of nucleic acid was used in a reverse transcription reaction with 100 pmol of primer FR20RV-12N (5’-GCCGGAGCTCTG CAGATATCNNNNNNNNNNNN-3’) using SuperScript IV reverse transcriptase (Thermo Fisher Scientific, Waltham, WA, USA), followed by second-strand synthesis using the Klenow Fragment of DNA polymerase (New England Biolabs, Ipswich, MA, USA) with primer FR20RV-12N at 10 pmol. After purification using Agencourt AMPure XP beads (Beckman Coulter, Chaska, MN), SISPA PCR amplification was conducted with TaKaRa Taq DNA Polymerase (Takara Bio, Shiga, Japan) using the primer FR20RV (5’-GCCGGAGCTCTG CAGATATC-3’) at 10 pmol. Sequencing libraries were prepared using 290 ng of double-stranded DNA as input to the SQK-LSK109 kit and barcoded individually using the EXP-NBD196 Native barcodes (Oxford Nanopore Technologies, Oxford, United Kingdom) in a modified version of one-pot native barcoding of amplicons (https://www.protocols.io/view/one-pot-native-barcoding-of-amplicons-j8nlke8q6l5r/v1). Libraries were sequenced in a FLO-MIN106 flow cell on the Mk1C device (Oxford Nanopore Technologies, Oxford, United Kingdom). Fast5 reads were basecalled and demultiplexed using Guppy version 6.2.1 (Oxford Nanopore Technologies, Oxford, United Kingdom). Fastq reads were then filtered by size and quality using Nanofilt and classified using Kraken version 2.1.0.5,27 A reference-based alignment was conducted to generate consensus sequences using a combination of Medaka (Oxford Nanopore Technologies, Oxford, United Kingdom), SAMtools, and BCFtools utilities. 14
Chromogenic In Situ Hybridization
Probes for chromogenic in situ hybridization (CISH) were designed and synthesized using the proprietary RNAscope CISH method (Advanced Cell Diagnostics, Newark, CA, USA) from the sequences obtained via next-generation sequencing. These included a proprietary 15 ZZ probe targeting the E6 and E7 genes of EdPV1 (V-EDPV1-E6-E7-O1-C1) and a proprietary 14 ZZ probe targeting the E6 and E7 genes of EdPV2 (V-EDPV2-E6-E7-O1). CISH was performed on 5-μm-thick sections of FFPE skin mounted on charged slides using the RNAscope 2.5 High-Definition RED Assay. 26 Grossly and histologically normal North American porcupine skin, in addition to a probe to the bacterial gene DapB, served as negative controls.
Results
Study Population
The age and sex of the 10 North American porcupines in this study are listed in Table 1. All animals are male. One biopsy was from a captive animal of unknown previous origin, and the remaining biopsy (n = 2) and necropsy (n = 7) cases were from wild animals that were either free-ranging or under medical care at the time of submission. For necropsy submissions, death was attributable to an infectious disease in most cases (n = 5), including bacterial sepsis (n = 3), viral infection (n = 3), and/or encephalitis associated with Baylisascaris sp. larval migration (n = 2). One necropsy case was found to have colonic feed impaction with rupture, and the final porcupine died due to acute trauma secondary to automobile strike.
Signalment, location, PCR, and CISH findings of North American porcupines with papillomas submitted to the New York State Animal Health Diagnostic Center.

Abbreviations: PCR, polymerase chain reaction; CISH, chromogenic in situ hybridization; NOS, not otherwise specified; n/d, not done; EdPV, Erethizon dorsatum papillomavirus.
Gross Findings and Histopathology
Gross examination of all cases (n = 10) revealed individual to dozens of multifocal to coalescing, pigmented, 1 mm3 to 7 mm × 3 mm × 2 mm, slightly raised to exophytic papules (Fig. 1a). Papillomas were distributed on the muzzle (n = 4), caudal dorsum (n = 1), upper lip (n = 1), chin (n = 1), gingiva (n = 2), and nasal planum (n = 1).
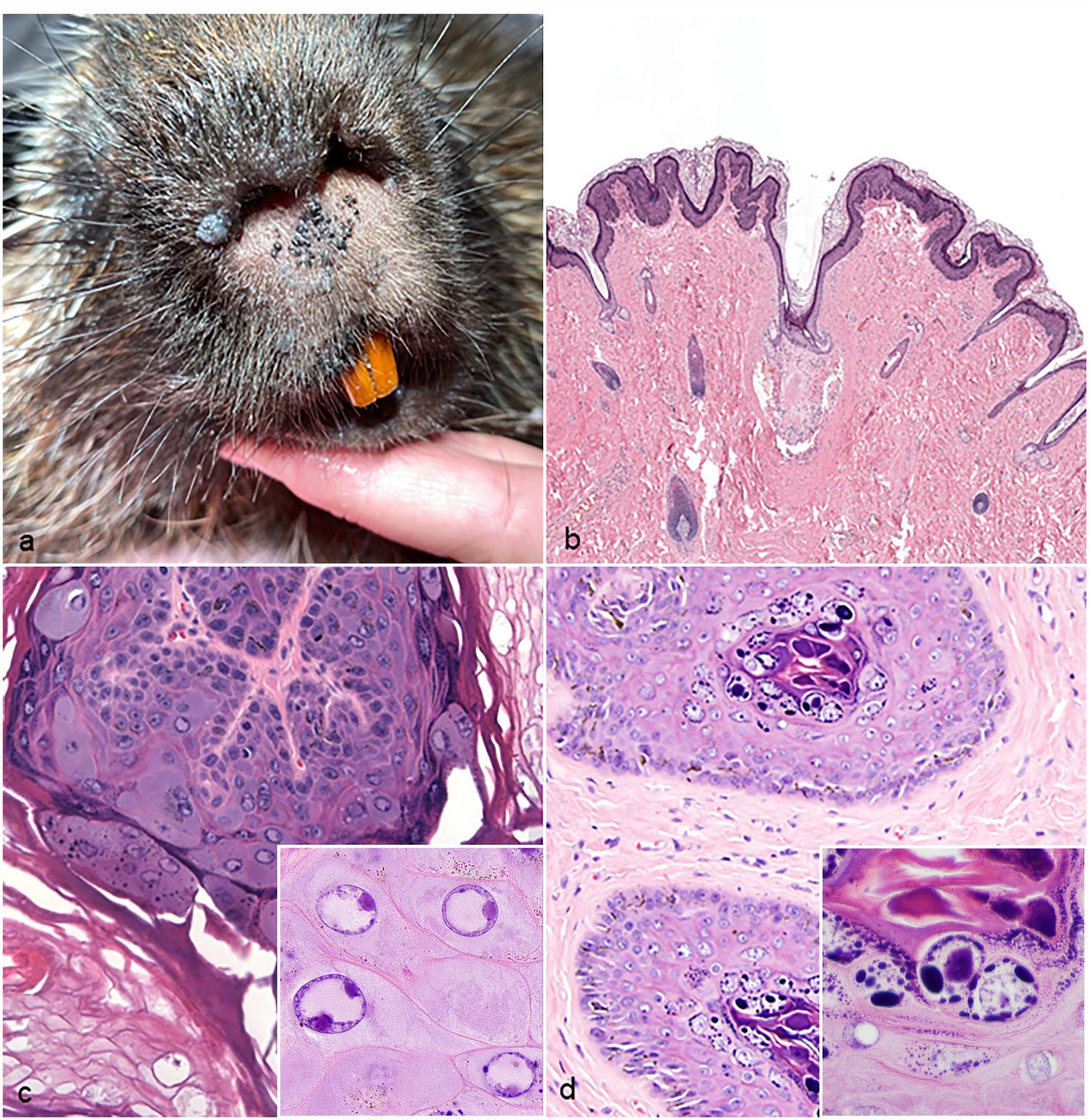
Skin from affected porcupines. (a) Papillomas, muzzle, North American porcupine. Extending across the muzzle are approximately two dozen, pigmented, exophytic, 1- to 3-mm-diameter papules. A larger exophytic papilloma is present along the right naris. (b) Papilloma, North American porcupine. The papilloma comprised hyperplastic epithelium overlain by orthokeratotic keratin. Hematoxylin and eosin (HE). (c) EdPV1 papilloma, North American porcupine. Keratinocytes within the stratum spinosum and granulosum contain numerous lightly amphophilic to eosinophilic, 8- to 10-μm-diameter, intranuclear inclusions. Inset highlights inclusion bodies; HE. (d) EdPV2 papilloma, North American porcupine. Deeply amphophilic, 15-μm-diameter, intranuclear inclusions are present in the stratum granulosum and corneum. The inset highlights inclusion bodies; HE.
Histologic examination of all cases (n = 10) revealed hyperplastic epithelium ranging from exophytic proliferations to sessile plaques, overlain by abundant amount of orthokeratotic keratin (Fig. 1b). Prominent and clumped keratohyalin granules were present within the stratum granulosum, and koilocytes were multifocally present within the stratum spinosum and granulosum. Koilocytes were defined as keratinocytes with a swollen, clear, cytoplasm and a shrunken nucleus with condensed chromatin. Occasional intranuclear inclusions were observed and included 2 distinct morphologies: approximately 8- to 10-μm-diameter, lightly amphophilic to eosinophilic, intranuclear inclusions in the stratum spinosum, granulosum, and follicular epithelium (Fig. 1c), and approximately 15- to 30-μm-diameter, deeply amphophilic, intranuclear inclusions in the stratum granulosum and corneum (Fig. 1d).
PCR and Intranuclear Inclusion Morphology
EdPV1 DNA was amplified from 6 porcupines (60.0%), including the single captive porcupine, and EdPV2 DNA was amplified from 3 remaining porcupines (30.0%). In one case, PCR was not pursued as the sample had been processed in a fixative decalcifier solution (Formical-4; StatLab), and DNA integrity was considered compromised. Intranuclear inclusion morphology on histologic assessment correlated to PCR results, with all EdPV1-positive cases (n = 6) having the smaller inclusions described above, and all EdPV2-positive cases (n = 3) having larger inclusions. Intranuclear inclusion morphology suggested EdPV2 infection in the case fixed in the decalcifier solution, as it had occasional, up to 30-μm-diameter, glassy, amphophilic inclusions within the stratum granulosum and corneum.
Next-Generation Sequencing
The consensus sequence of EdPV1 resulted in 100% genome coverage with 99.84% identity to EdPV1 (GenBank: MZ647948.1) and was submitted to GenBank under accession OP605550. The consensus sequence of EdPV2 resulted in 76.20% genome coverage at 10X reads depth, with 95.98% identity to EdPV2 (GenBank: MZ647949.1). Of the two published EdPV2 genomes in GenBank, the L1 gene shared 99.8% identity to one accession (GenBank: MZ647949) and 92.9% identity to another (GenBank: MH376689). Despite the lower genome coverage of EdPV2 sequence, the region encoding L1 protein had complete coverage, allowing for the characterization of the virus based on this conserved region. This sequence was submitted to GenBank under accession OP605549. The EdPV1 and EdPV2 sequences obtained shared 34.3% whole-genome identity and 53% L1 protein identity (Supplemental Fig. S1). EdPV1 is the sole member of the genus Sigmapapillomavirus, whereas the EdPV2 sequence belongs to a separate genus in the family Papillomaviridae, with 71% identity to chimpanzee papillomavirus and 77%–83% identity to bovine papillomavirus 12.
Chromogenic In Situ Hybridization
Positive hybridization signals for EdPV1 or EdPV2 were detected in 8 of 9 papillomas, with intense nuclear hybridization signals within the superficial epithelium, including strata corneum, lucidum, granulosum, and spinosum. Punctate nuclear and cytoplasmic hybridization signals were limited to basal epithelium in all cases (Fig. 2). Hybridization was not detected in the dermis and was diffusely negative in sections of grossly and histologically normal North American porcupine skin. All papillomas with positive CISH hybridization signals were also positive on EdPV1 or EdPV2 PCR. One papilloma did not hybridize with either EdPV1 or EdPV2 probes, and PCR was not pursued in this case given fixation in a formical solution. All EdPV2 PCR-positive cases did not exhibit EdPV1 CISH hybridization, and 5 of 6 EdPV1 PCR-positive cases did not exhibit EdPV2 CISH hybridization. In one case where EdPV1 DNA was amplified, CISH revealed positive hybridization signaling for both EdPV1 and EdPV2, suggestive of a coinfection. This case additionally contained both intranuclear inclusion morphologies described above upon histopathologic evaluation.
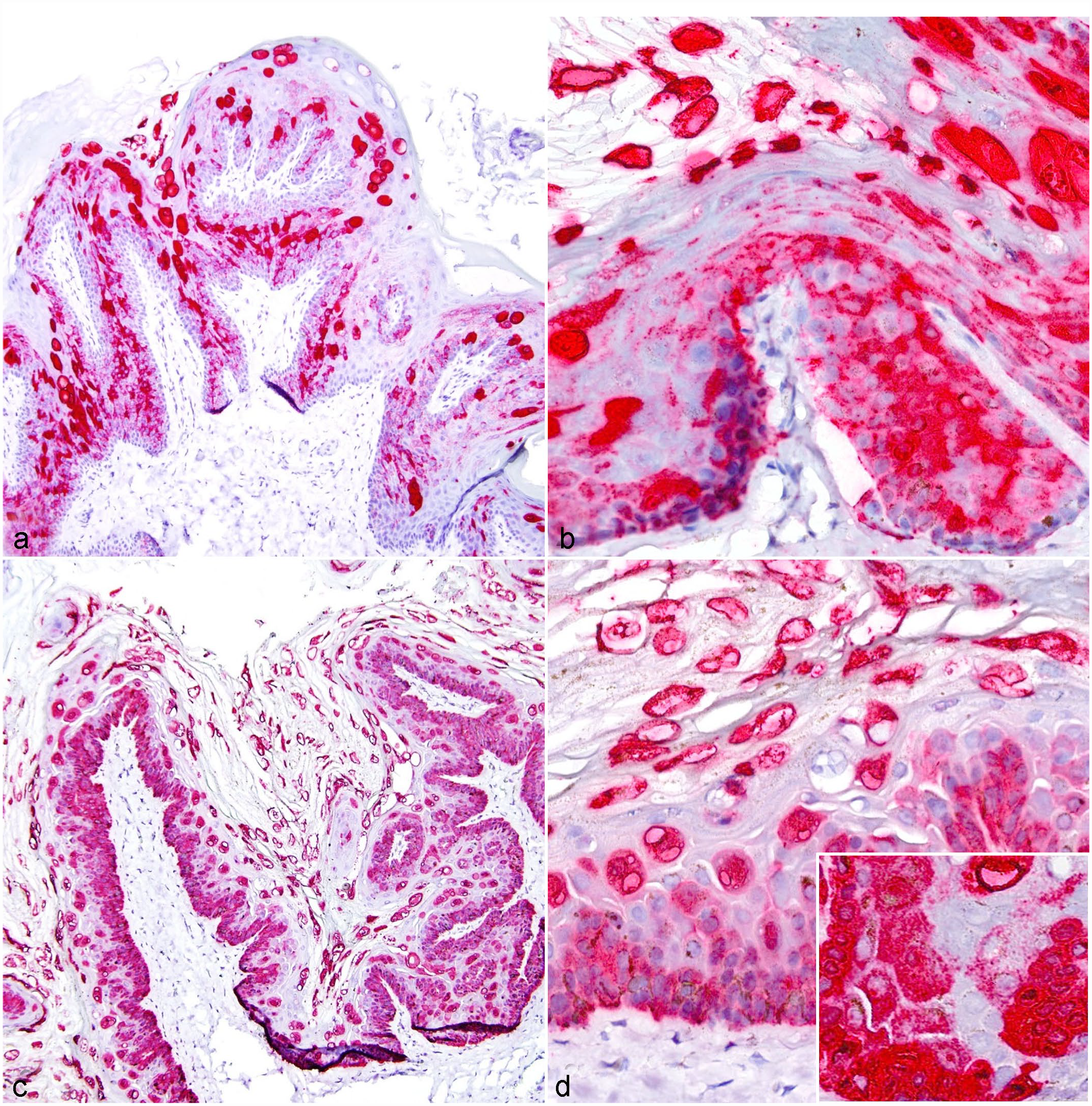
Chromogenic in situ hybridization detection of Erethizon dorsatum papillomavirus 1 (EdPV1) and 2 (EdPV2) in the epidermis of North American porcupines. (a, b) EdPV1 infection, skin, North American porcupine, EdPV1 chromogenic in situ hybridization. (a) There is intense nuclear hybridization signals within the superficial epidermis and punctate nuclear and cytoplasmic hybridization signals limited to the basal epithelial layers. (b) A higher magnification image of (a). (c, d) EdPV2 infection, skin, North American porcupine, EdPV2 chromogenic in situ hybridization. (c) There is intense nuclear hybridization signals within the superficial epidermis and punctate nuclear and cytoplasmic hybridization signals limited to the basal epithelial layers. (d) A higher magnification image of (c).
Discussion
This study identified and characterized EdPV1- and EdPV2-associated papillomas in wild and captive North American porcupines in New York State. Gross and histologic features were similar to those previously described in single case reports of infection in captive North American porcupines. Interestingly, unique intranuclear inclusion morphologies and distributions were identified in EdPV1 versus EdPV2 infections and correlated to PCR and CISH results. In all cases, EdPV1 infection was associated with smaller (8–10 μm in diameter) and paler intranuclear inclusions in the stratum spinosum, granulosum, and follicular epithelium, whereas EdPV2 infection was associated with larger (15–30 μm in diameter) and darker intranuclear inclusions limited to the stratum granulosum and corneum. A similar EdPV2 inclusion morphology has been previously reported, and evaluation of inclusion morphology may provide preliminary information on distinguishing between EdPV1 and EdPV2 infection. 21 Screening additional cases at multiple laboratories is necessary to further strengthen this claim.
Gross lesion distribution also correlated to the EdPV species, with papillomas on the muzzle exclusively corresponding to EdPV1 infection and accounting for most EdPV1 PCR-positive cases (5/6). The remaining EdPV1 papilloma was present on the nasal planum. EdPV2 infection was associated with a papilloma on the caudal dorsum in one porcupine, a location not previously reported for EdPV infection to the authors’ knowledge.18,21,24,25 Remaining EdPV2 PCR-positive papillomas were distributed on the chin (n = 1) and gingiva (n = 1). An additional case fixed in a decalcifier solution that was suspected to reflect EdPV2 infection based on inclusion morphology was also located on the gingiva. Papillomas were identified in both ill and apparently healthy porcupines, with most necropsy cases diagnosed with a concurrent bacterial, viral, or parasitic disease. Although immunosuppression has been associated with papillomavirus-associated disease in a number of species, the role it plays in EdPV1 and EdPV2 infections in North American porcupines is unknown. 16
All EdPV1 and EdPV2 PCR-positive cases exhibited correlative positive hybridization signaling via CISH. One EdPV1 PCR-positive case had positive hybridization signals to both EdPV1 and EdPV2 probes, suggesting dual infection and greater sensitivity for virus detection with CISH over PCR alone. In addition to screening for the presence of EdPV nucleic acid, CISH also highlighted the localization of E6 and E7 hybridization signals. In human and veterinary medicine, E6 and E7 are recognized viral oncogenes associated with tumor pathogenesis.1,17,28 Strong nuclear hybridization signals in the superficial epithelial layers and punctate cytoplasmic and nuclear hybridization in the basal and suprabasal keratinocytes have been thought to indicate productive papillomavirus infection, whereas punctate cytoplasmic and nuclear hybridization throughout the epidermis has been associated with transformative phase infections.9,13 The hybridization pattern of papillomas in this study was suggestive of productive infection, which correlated with a lack of histologic evidence of malignant transformation. Whether EdPV infection can be associated with the development of malignancy has yet to be determined.
Marked divergence of EdPV1 and EdPV2 suggest these viruses arose independently within North American porcupines. Based on the obtained whole-genome sequence from one EdPV2 PCR- and CISH-positive papilloma, nucleotide identity of L1 was found to be 92.9% to a whole-genome sequence of EdPV2 published in 2018 (GenBank: MH376689) and 99.8% to a whole-genome sequence of EdPV2 published in 2023 (GenBank: MZ647949). 25 Current taxonomic guidelines of papillomaviruses state that differences in nucleotide identity between 2% and 10% classifies a novel subtype, thus characterizing at least one papilloma in this case as a novel subtype of EdPV2 only previously identified in a captive porcupine in France.6,24
All EdPV2 PCR- and CISH-positive papillomas in this study were identified in wild North American porcupines in New York State. A more extensive distribution is suspected given previous case reports of EdPV2 infection in captive porcupines in Arizona and France, and it remains uncertain whether EdPV2 originated in captive or wild populations. Within the United States, EdPV1 infection has been exclusively diagnosed in New York in both captive and wild populations, which may suggest a more narrow range or may alternatively be a consequence of limited reports in the literature as papillomavirus infections in wildlife have been suggested to extend in an environmental range similar to the range of their host species. 10 Transmission of papillomaviruses typically occurs via cutaneous or mucosal contact, which is suspected to be the case with porcupine papillomaviruses given a high prevalence of lesions on facial or oral epithelial surfaces. Given all porcupines in this study were male (n = 10), it was considered possible that male North American porcupines face increased intraspecies contact; however, this may alternatively reflect an increased prevalence of males received to the New York State Animal Health Diagnostic Center over the course of the study period. Given papillomavirus route of transmission, widespread distribution of EdPV is plausible as the North American porcupine can be found across the Western and Northeastern United States, in addition to Mexico and most of Canada. 7
In conclusion, EdPV1 and EdPV2 are present within wild and captive populations of North American porcupines in New York State. Histopathology, in combination with PCR and CISH, can be used to detect and differentiate these infections. Further studies are warranted to fully characterize the viruses’ prevalence and clinical significance.
Supplemental Material
sj-pdf-1-vet-10.1177_03009858231176564 – Supplemental material for Histopathologic and molecular characterization of Erethizon dorsatum papillomavirus 1 and Erethizon dorsatum papillomavirus 2 infection in North American porcupines (Erethizon dorsatum)
Supplemental material, sj-pdf-1-vet-10.1177_03009858231176564 for Histopathologic and molecular characterization of Erethizon dorsatum papillomavirus 1 and Erethizon dorsatum papillomavirus 2 infection in North American porcupines (Erethizon dorsatum) by Zoe E. Mack, Leonardo C. Caserta, Randall W. Renshaw, Shotaro Nakagun, Rhea S. Gerdes, Diego G. Diel, Sara E. Childs-Sanford and Jeanine Peters-Kennedy in Veterinary Pathology
Footnotes
Acknowledgements
We would like to thank the histology laboratory at the Cornell University College of Veterinary Medicine Animal Health Diagnostic Center for slide preparation and processing. We would also like to thank Dr. Mason C. Jager, DVM, PhD, DACVP, for assistance in CISH processing.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This project was supported by research funds provided by Cornell University College of Veterinary Medicine.
Supplemental Material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.