
Abstract
From 2016 to 2018, an epidemic wave of yellow fever (YF) occurred in Brazil, affecting a large number of Platyrrhini monkeys. Titi monkeys (Callicebus spp.) were severely affected yet pathological characterizations are lacking. This study characterized epizootic YF in 43 titi monkeys (Callicebus spp.) with respect to the microscopic lesions in liver, kidney, spleen, heart, brain, and lung, as well as the distribution of immunolabeling for YF virus antigen, and the flaviviral load in the liver. Of 43 titi monkeys examined, 18 (42%) were positive for yellow fever virus (YFV) by immunohistochemistry or reverse transcription quantitative real-time polymerase chain reaction (RT-qPCR). Affected livers had consistent marked panlobular necrotizing hepatitis, lipidosis, and mild inflammation, with intense immunolabeling for YFV mainly in centrilobular hepatocytes (zone 1; P = .05). In the spleen, consistent findings were variable lymphoid depletion (10/11), lymphoid necrosis (lymphocytolysis; 4/11), and immunolabeling for YFV in histiocytic cells (3/16). The main finding in the kidney was multifocal acute necrosis of tubular epithelium (5/7) that was occasionally associated with intracytoplasmic immunolabeling for YFV (6/15). These data indicate that titi monkeys are susceptible to YFV infection, developing severe hepatic lesions and high viral loads, comparable to humans and Alouatta spp. Thus, Callicebus spp. may be reliable sentinels for YF surveillance.
Keywords
Titi monkeys (family Pitheciidae) are New World primates (NWP) from South America that encompass 3 genera: Cheracebus, Callicebus, and Plecturocebus. 3 Callicebus species (Thomas, 1903) live in Atlantic Forest, distributed in Northeast and Southeast regions of Brazil, 3 and include 5 species that are classified as “near threatened” to “critically endangered” (IUCN red list). 13 Between 2016 and 2018, epidemic waves of yellow fever (YF) occurred in Brazil, resulting in more than 1800 and 700 confirmed cases in NWP and humans, respectively. 21 YF is an infectious, vector-borne hemorrhagic zoonosis caused by yellow fever virus (YFV; family Flaviviridae, genus Flavivirus prototype virus). YF exhibits a sylvatic cycle mainly maintained between NWP hosts and canopy mosquitos, that is, Sabethes and Haemagogus, in addition to an urban cycle with sustained human transmission by Aedes aegypti mosquitos. The latter has not occurred in Brazil since the beginning of the last century. 20 Howler monkeys (Alouatta spp., family Atelidae) are particularly susceptible to YF and may develop fatal hepatitis characterized by necrosis and presence of apoptotic bodies. 15 Titi monkeys were affected during the last YF epizootic, based mainly on morphological evidence, 12 yet pathological characterizations are lacking. This study characterized the histopathological and immunohistochemical features of epizootic YF in Callicebus spp. from São Paulo state within the YF National Surveillance Program.
Materials and Methods
Samples
We analyzed frozen and formalin-fixed paraffin-embedded tissue samples from 43 Callicebus spp. monkeys from a total of 4066 NWP submitted to the Adolfo Lutz Institute, as part of the National Surveillance Program of Epizootics (instituted by Brazilian Ministry of Health, from São Paulo State, Brazil; 21°49′47″S, 49°12′27″W), from January 2016 to December 2018. The samples came from standardized necropsies performed by local surveillance agents, including trained veterinarians and other personnel. 19 Epidemiologic (carcass location and date) and biologic (species/genus identification, sex, and age) data were retrieved from the submission forms (Sistema Nacional de Agravos de Notificação “SINAN”). All these proceedings were approved by the Animal Use Committee by Adolfo Lutz Institute (CEUA-IAL no. 11/2016 and CEUA-VET 4661190618) and ICMBIO Protocol 50551.
Histopathologic and Immunohistochemical Analyses
All formalin-fixed paraffin-embedded tissue samples (liver, kidney, spleen, heart, brain, and lung) were routinely processed and stained with hematoxylin and eosin for histopathologic examination. Histopathologic description was performed only in cases that were positive for YFV by immunohistochemistry (IHC) or RT-qPCR (reverse transcription quantitative real-time polymerase chain reaction), and autolyzed samples were excluded. The grade as 1 to 4 (1 was mild and 4 was severe), distribution (periportal, midzonal, centrilobular, or panlobular for liver; focal, multifocal, diffuse for other organs), and inflammatory cellular components (neutrophils, mononuclear cells, or both) were determined as previously described 10 (Suppl. Table 1).
IHC analysis was performed as described. 10 Selected tissue sections (liver, kidney, spleen, heart, lung, and brain) were submitted to antigen retrieval in a pressure cooker in citrate buffer for 3 minutes (120 °C, pH 6.0) and then incubated overnight with polyclonal anti-YFV antibody (mouse hyperimmune antisera against wild strain SPH144990, Núcleo de Doença de Transmissão Vetorial, Virology Center, IAL, São Paulo, Brazil). Signal amplification was achieved by HRP-conjugated polymer detection system (Reveal biotin-free polyvalent, Spring Bioscience Corp), visualization with diaminobenzidine (D-5637; Sigma-Aldrich), and counterstaining with Harris hematoxylin. Known positive tissues and negative control tissues with omitted primary antibody were included. Viral antigen in liver was semiquantified based in Allred scoring method, by the sum of a proportion score (PS) of cells immunolabeled (0: negative, 1: 1% to 10%, 2: 10% to 50%, 3: 50% to 70%, and 4: 70% to 100%) and score of the intensity of immunolabeling (0: negative, 1: weak, 2: moderate, 3 moderate to strong, 4: strong). 23 The final score (proportion score + intensity score) was obtained for each hepatic zone and the sum of zones indicated the total hepatic score. For other organs, IHC was only classified as positive or negative. IHC was performed even in markedly autolyzed/decomposed samples.
Molecular Analyses
Protocols and procedures were conducted within the enhanced laboratory biosafety level 2 (Animal Biosafety Level 2) facilities at IAL. Total RNA was extracted from fresh frozen liver, using QIAamp RNA Blood Mini Kit (QIAGEN), following the manufacturer’s instructions. Amplification of YFV fragment was performed employing a protocol designed previously, 6 which targets the highly conserved 5′ noncoding region of the genome with 112 bp length and is based on a hydrolysis probe system RT-qPCR protocol. Quantification cycle (Cq) was used as reference to viral load, considering that, in a standard curve with YFV vaccine 17-DD, a titer of 105 PFU/ml had a median Cq = 16, while 1 PFU/ml had a median Cq = 35.6. 27
Statistical Analysis
Data were tabulated and analyzed with Portal Action (http://www.portalaction.com.br) and Minitab software. For categorical data, we applied χ2 and/or Fisher’s exact test (2-tailed). For continuous data, after Shapiro-Wilk normality test, Kruskal-Wallis test was applied. Correlation of nonparametric data was calculated by Spearman test, and a standard curve was elaborated. Statistical difference was considered when P < .05.
Results
The 43 titi monkeys examined included 14 females, 17 males, and 12 of undetermined sex with an age distribution of 5 infant/juvenile, 17 adult, and 21 of undetermined age. Some of the animals were described as Callicebus nigrifons in the files, others as Callicebus spp., but only C. nigrifons occurs naturally in the geographic region analyzed. Eighteen of 43 (42%) animals were positive by RT-qPCR and/or IHC; there was no sex association with YFV positivity (P = .55). Six of 25 (24%) animals that tested negative for YFV had details about case of death in the file: electrocution (n = 1), vehicle collision (n = 2), dog attack (n = 2), and trauma (n = 1). All animals were from São Paulo State, Brazil. Figure 1 illustrates geographic distribution of positive and negative Callicebus, sex and age profile. Supplemental Figure S1 illustrates the flowchart of samples, including samples analyzed by each method and those excluded. The liver histopathology and immunohistochemistry findings are presented in Supplemental Table S2.
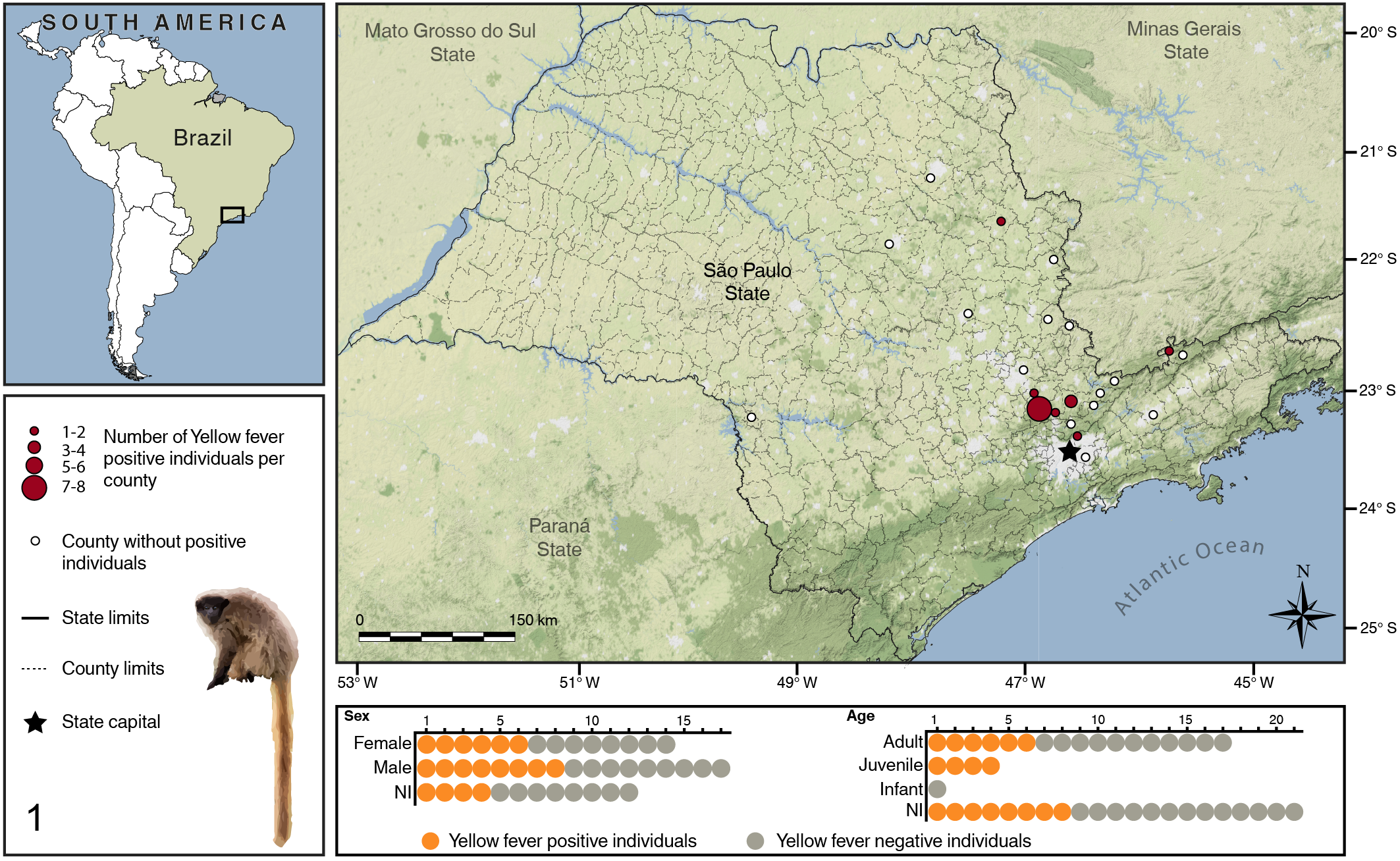
Geographic distribution and demographics of Callicebinae monkeys included in this study. NI: Unknown.
Histopathology
Samples from liver (n = 15/18), spleen (n = 11/16), kidney (n = 7/15), lung (n = 8/13), heart (n = 11/14), and brain (n = 9/16) were evaluated (denominators less than 18 indicate those excluded because of autolysis). The main histopathologic changes were consistently present in the liver and were necrotizing hepatitis (13/15, 87%) with median intensity 3 (range 2–4) and panlobular (diffuse, massive) distribution (10/13; 77%); 3 cases had midzonal necrosis (Fig. 2). Two of 15 (13%) animals had no liver necrosis and while no flaviviral antigen was detected by IHC, flaviviral nucleic acid was detected by RT-qPCR (Cq values of 33 and 35). The inflammatory reaction patterns included an admixture of neutrophils and mononuclear cells including lymphocytes and plasma cells (6/13; 46%) or only mononuclear cells (7/13; 54%). These were typically dispersed through the hepatic lobule or rarely confined to portal areas (3/13; 23%). Apoptotic bodies (Fig. 2) were consistently present in necrotic areas. Microvacuolar lipidosis was present in 14/15 (93%; median intensity 3, range 1–3) cases, and was occasionally associated with macrovacuolar lipidosis (6/14; 43%). One of the negative cases by IHC (RT-qPCR, Cq: 33) had moderate panlobular lipidosis as the only microscopic finding in the liver. Hemorrhage was present in 4/15 (27%) cases and was restricted to centrilobular (3/4) or midzonal (1/4) areas. Five cases (5/15, 33%) had a ductular reaction (Fig. 3).
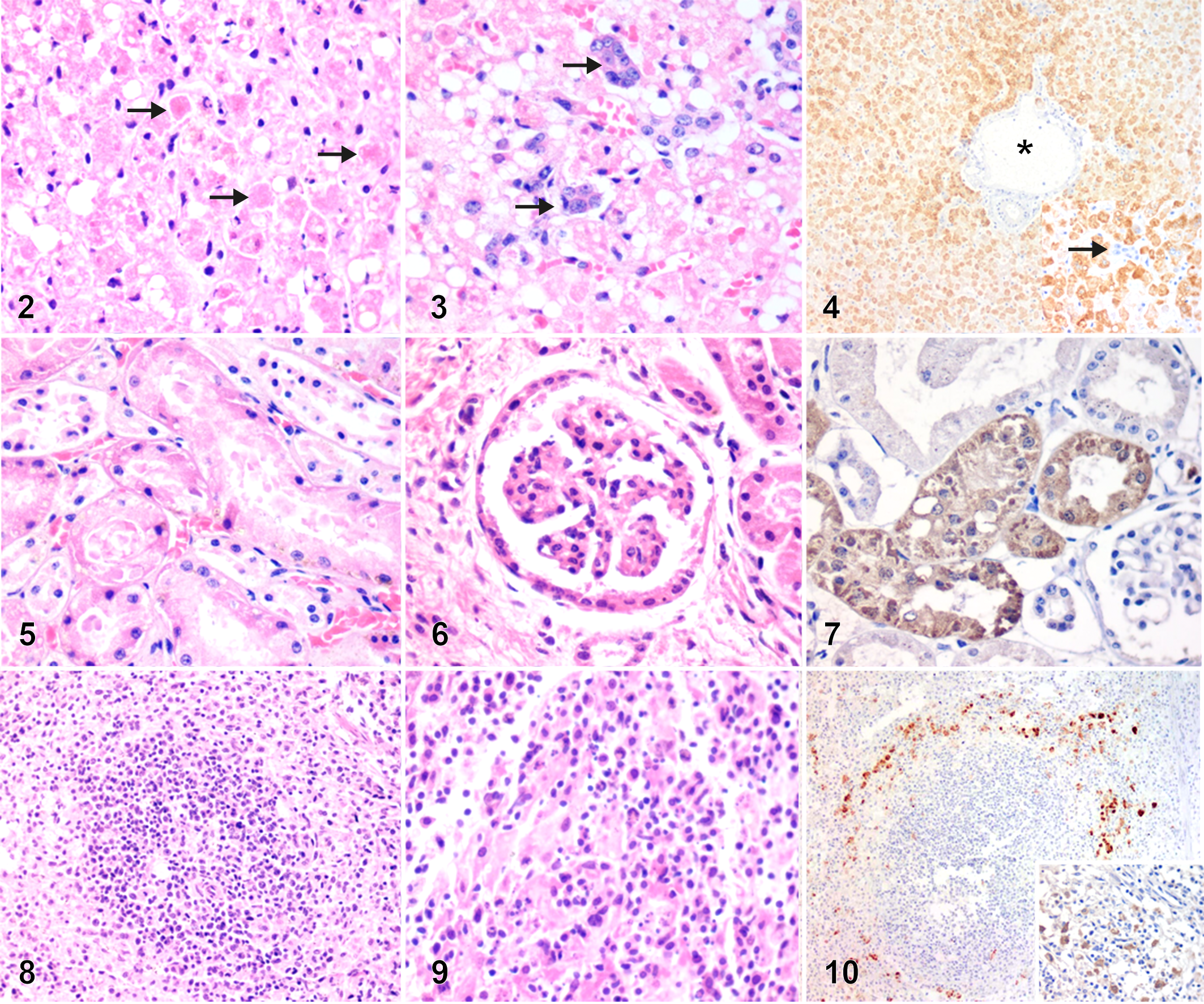
Yellow fever, titi monkeys.
In the kidney, 5/7 cases had acute necrosis of tubular epithelium (Fig. 5), and 6/7 had glomerular and tubular proteinosis. Renal inflammation was not observed. Less common findings were hypertrophy of the glomerular parietal epithelium (2/7; Fig. 6), and vacuolation of tubular epithelium (2/7).
In the spleen, depletion of the white pulp prevailed (10/11; Fig. 8), often associated with lymphoid necrosis (4/11; Fig. 9), disruption of red pulp, or necrosis of splenic cords (2/11). In the lungs, there was occasional alveolar hemorrhage (2/8) and edema (4/8). Mild lympho-histiocytic interstitial pneumonia, mild, focal, mononuclear myocarditis, mild non-suppurative meningoencephalitis and meningitis were present in one case each.
Immunohistochemistry and RT-qPCR
IHC for YC was performed in all cases (n = 18) regardless of autolysis status. Flaviviral antigen was detected primarily in liver in 16/18 (89%) cases (Suppl. Fig. S3). The liver of 2/18 (13%) cases were negative by IHC but were RT-qPCR-positive with Cq values of 33 and 35. Flaviviral antigen was consistently detected in the cytoplasm of hepatocytes (Fig. 4), in all hepatic zones albeit with higher intensity in periportal areas (P = .05; Suppl. Fig. S2). There was no correlation between viral load (Cq value) and intensity scores for immunolabeling of viral antigen (P = .121, ρ = −0.434). There was positive correlation among intensity scores for viral antigen in different hepatic zones (periportal [PP] vs midzonal [MZ]: P = .004 and ρ = 0.719; PP vs centrilobular [CL]: P = .001 and ρ = 0.661; MZ vs CL: P = .03 and ρ = 0.571). Viral antigen was also detected in spleen (3/16; 18%), specifically within macrophages/dendritic cells near the white pulp (Fig. 10), and in renal tubular epithelial cells (6/15, 40%; Fig. 7). Viral antigen was not detected in other organs examined (lung, heart, brain). In the 14/18 samples that were adequate for RT-qPCR analysis, Cq values exhibited a nonnormal distribution, with median 7 (range 5–35).
Discussion
We described the histopathological and immunohistochemical features of epizootic YF in titi monkeys from São Paulo State, Brazil. YFV is a zoonotic flavivirus with a sylvatic cycle involving NWP in South America. YFV is maintained in NHP and may affect humans, especially when they encroach forest environments, resulting in epidemics as observed in Brazil during 2016 to 2018. 21 The high susceptibility and frequent early fatalities among NWP compared to humans is one of the principles of the Brazilian surveillance for YF. Thus, NWP act as sentinels. 1 Platyrrhini, a heterogeneous group composed of 5 families, 21 genera, and 176 species, 24 is described as susceptible to YFV. However, most reports are focused in Alouatta spp., and, more recently, in Callithrix spp. 1,4,10,15
In this study, most of the animals evaluated were identified as Callicebus nigrifons from the northeast portion of São Paulo State, an area with known YFV circulation. 4 Callicebus genus is rarely described as susceptible to YFV. 12,25 The first record of a Callicebus with detectable YFV in the Information System on Diseases of Compulsory Declaration from Brazilian Ministry of Health (SINAN) occurred in 2016; YF cases in Callicebus spp. comprised 0.45% of the records in the country. 12 In our caseload, Callicebus spp. represented 1% of NWP analyzed during the epidemic waves. Despite this relatively low representation in the databases, they are susceptible to YF highlighting potential implications of the disease in populations, especially in fragmented areas.
In humans, YF has a broad spectrum of clinical findings, from asymptomatic disease to fulminant hepatitis. Classically, the histopathological signature of YF includes severe midzonal to panlobular necrotizing hepatitis with apoptotic bodies (Councilman-Rocha Lima bodies), variable lipidosis, and mild inflammation. 14,28 Zonal necrosis with macro- and microvacuolar lipidosis, centrilobular hemorrhage, and cholestasis have been reported in livers of humans affected by YF. 7 Comparatively, YF-infected Callicebus exhibited extensive panlobular necrosis with no evident midzonal demarcation, and consistent apoptotic bodies. Furthermore, YFV antigen was detected in all hepatic zones but more prominently in periportal areas, and restricted to hepatocytes. This differs from the predominant midzonal distribution of YFV antigen in human fatalities. 2 The preferential distribution of lesions in zone 2 is also reported in other arboviral infections, and it is hypothesized to be related to a greater YFV tropism for midzonal hepatocytes and a greater susceptibility of these hepatocytes to low oxygen concentrations resulting from endothelial injury. 22 It is believed that YFV binds in a nonspecific manner to glycosaminoglycan heparan sulfate on the surface of host cells such as hepatocytes or dendritic cells; 11,16 however, specific receptor-mediated viral entry and receptor distribution within the hepatic lobule remain to be investigated. The differences observed, namely, panlobular necrosis and diffuse YFV-antigen distribution, indicate high susceptibility of Callicebus to YFV. Further studies to characterize potential species-specific pathological signatures as well as local inflammatory response patterns to YFV among NWP are warranted.
Nearly all animals examined had some degree of lymphoid depletion, frequently associated with lymphoid necrosis, in the spleen. Experimentally, golden hamsters (Mesocricetus auratus) developed lymphoid hyperplasia after YFV inoculation, followed by severe lymphoid depletion associated with widespread lymphoid necrosis and lymphocytolysis, 26 and infected rhesus macaques (Macaca mulatta) developed white pulp necrosis and peripheral lymphopenia. 9 While the liver remains the primary tissue for histopathological and IHC diagnosis of YF, concomitant splenic lymphoid necrosis lends further support to the diagnosis within current YF surveillance. Lymphoid depletion is also noted in human YF cases, 8 although other conditions such as stressful events may be associated with lymphocytolysis.
Acute renal tubular epithelial necrosis was common in affected animals and is described in rhesus macaque models with kidney abnormalities found in late stages of disease, but without YFV immunolabeling. 9 Hyperbilirubinemia, rhabdomyolysis, and intravascular coagulation are mechanisms suspected to contribute to kidney injury in YF; direct cellular damage by YFV in the kidney has not been proven. 17 We did not detect relevant histological findings in lungs, heart, or brain, except for acute hemodynamic alterations, such as hemorrhage, congestion, and edema. Similarly, a low frequency of changes in these organs was described in Alouatta monkeys during the 2009–2010 epidemic, and extrahepatic histologic lesions related to YF were not identified in NWP, mainly Alouatta and Callithrix. 25 In humans, pulmonary alveolar hemorrhage, interstitial pneumonia, renal tubular necrosis, proliferative glomerulonephritis, and splenic white pulp depletion have been described. 8 In the present study, YFV antigen was also detected in spleen and kidney of a small number of animals, which is in agreement with observations in YF fatalities in humans. 5,7 Furthermore, in humans with fulminant hepatitis undergoing liver transplant, the engrafted liver from a YF-negative donor developed YF lesions with viral antigen in hepatocytes some days after the surgical procedure. This lends support to extrahepatic replication of YFV, for instance, in kidney and spleen, 7 and further underscores the potential relevance of extrahepatic locations in YF pathogenesis and diagnosis.
Callicebus spp. monkeys exhibited a high viral load in liver, with a low median Cq value. High viral loads were described in various organs and blood from Callicebus, Alouatta, and Sapajus genera. 4,18 This may suggest that monkeys of genus Callicebus, similarly to Alouatta, could play a role as an amplifier of YFV. The potential epidemiological implications remain unknown because Callicebus spp. comprise small populations restricted to some geographic areas. Two animals presented low viral loads and had neither microscopic lesions nor detectable YFV antigen in the organs examined. We could not conclude the cause of death of these animals based in microscopic lesions and absence of gross information. This suggests that viral load may be related to lesion severity in liver; thus, higher viral loads may be associated with extensive liver injury. This finding could potentially be used to identify species-specific susceptibilities among NWP.
In conclusion, titi monkeys (Callicebinae) are susceptible to YF and may develop extensive hepatic necrosis, diffuse flaviviral antigen distribution with preferential periportal zone location, and high viral loads. The histopathological, immunohistochemical, and molecular findings differ from most human cases and are comparable to cases in other highly susceptible NWP.
Supplemental Material
Supplemental Material, sj-pdf-1-vet-10.1177_03009858211009781 - Yellow Fever as Cause of Death of Titi Monkeys (Callicebus Spp.)
Supplemental Material, sj-pdf-1-vet-10.1177_03009858211009781 for Yellow Fever as Cause of Death of Titi Monkeys (Callicebus Spp.) by Natália C. C. de A. Fernandes, Mariana S. Cunha, Juliana M. Guerra, Josué Diaz-Delgado, Rodrigo A. Ressio, Cinthya S. Cirqueira, Cristina T. Kanamura, Danny Fuentes-Castillo and José Luiz Catão-Dias in Veterinary Pathology
Footnotes
Acknowledgements
To Julia de Carvalho, Marcelo Sansoni, Filipe Nagamori, Simone Guimarães, and Isis Paixão de Jesus for excellent technical support. To Thamires Goes, in memoriam; to Victor Yunes, for taxonomic counseling; Secretaria da Saúde do Estado de São Paulo, for general support; local health agents; and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). This article is part of the PhD thesis project of Natália Coelho Couto de Azevedo Fernandes.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: To FESIMA/GAPS for funding of personnel (28/2020). José Luiz Catão-Dias is the recipient of a professorship from CNPq (304999-18).
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.