
Abstract
Squamous cell carcinoma (SCC) is the most common neoplasm of the equine stomach. However, the mechanisms underlying malignant transformation are unknown. As Equus caballus papillomavirus–2 (EcPV-2) is a likely cause of some genital SCCs, we hypothesized that EcPV-2 is associated with a subset of equine gastric SCCs. To this aim, we performed polymerase chain reaction (PCR) and in situ hybridization (ISH) for EcPV-2 E6/ E7 oncogenes on 11 gastric SCCs and on gastric samples from 15 control horses with no SCC. PCR for EcPV-2 was positive in 7/11 (64%) gastric SCCs; non-SCC gastric samples were all negative. Intense hybridization signals for EcPV-2 E6/E7 nucleic acid were detected by ISH within tumor cells in 5/11 (45%) gastric SCCs, including distant metastases. No hybridization signals were detected within any of the non-SCC gastric cases. This study provides support for a potential association between EcPV-2 infection and a subset of equine gastric SCC.
Squamous cell carcinoma is the most common type of stomach cancer in the horse. 9 These cancers are most often recognized late in the course of disease and are associated with a poor prognosis. 9 Thus far, an underlying cause for this type of cancer has not been identified. Human papillomaviruses (HPVs) are oncogenic viruses with a well-established causal association with several types of cancer, including squamous cell carcinoma. 2 Virtually all cases of human cervical cancers, as well as a subset of other genital and oropharyngeal cancers, are causally associated with certain types of HPV. 2 The HPV oncogenes E6 and E7 are upregulated within these cancers and drive carcinogenesis. 2 These oncogenes interfere with host cell cycle regulation and apoptosis, resulting in unregulated cell growth and genetic instability. 2
Similarly, there is increasing evidence that equine papillomaviruses may play a role in oncogenesis of certain types of tumors. 7 Equus caballus papillomavirus type–2 (EcPV-2) likely plays a causative role in the development of a subset of genital squamous cell carcinomas. 3,4,6 –8,10 Accumulating evidence for this association includes consistent detection of EcPV-2 DNA within genital SCC and precursor lesions, as well as identification of oncogenic E6 transcripts within genital SCC. 3,4,6 –8,10 These lesions typically progress from precursor lesions, such as papillomatous plaques and papillomas, to carcinoma in situ, and ultimately to SCC. 6
Given the association of EcPV-2 with a subset of genital SCC, we hypothesized that EcPV-2 is also associated with a subset of gastric SCC. To this aim, we performed PCR and in situ hybridization for EcPV-2 E6/ E7 on 11 equine gastric SCC cases. Tissue samples were collected from archived biopsies and necropsies from multiple institutions (Table 1): (1) North Carolina State University’s College of Veterinary Medicine (archives searched 2008–2018); (2) Rollins Animal Disease Diagnostic Laboratory (archives searched 2012–2018); (3) University of Pennsylvania, School of Veterinary Medicine (archives searched 2007–2018); and (4) Animal Health Laboratory, University of Guelph (archives searched 1997–2018). A full necropsy was performed on all cases except for 2 (Cases 2 and 10). Median age at time of diagnosis was 19 years, with an age range from 3 to 24 years. All cases were male. Metastases were identified in 7 of 9 (78%) cases with a full necropsy. Fifteen control horses with no SCC were used; these included normal stomach, gastric hyperplasia, gastritis, and leiomyoma (Controls 12–26, Suppl. Table S1). Median age at time of biopsy/ necropsy for controls was 14 years, with an age range from 1 to 35 years. Controls included 13 males and 2 females and spanned similar breeds and years of diagnosis as gastric SCC cases.
PCR and In Situ Hybridization for EcPV2 E6/E7 Viral Nucleic Acid Within Gastric Squamous Cell Carcinomas in Horses.
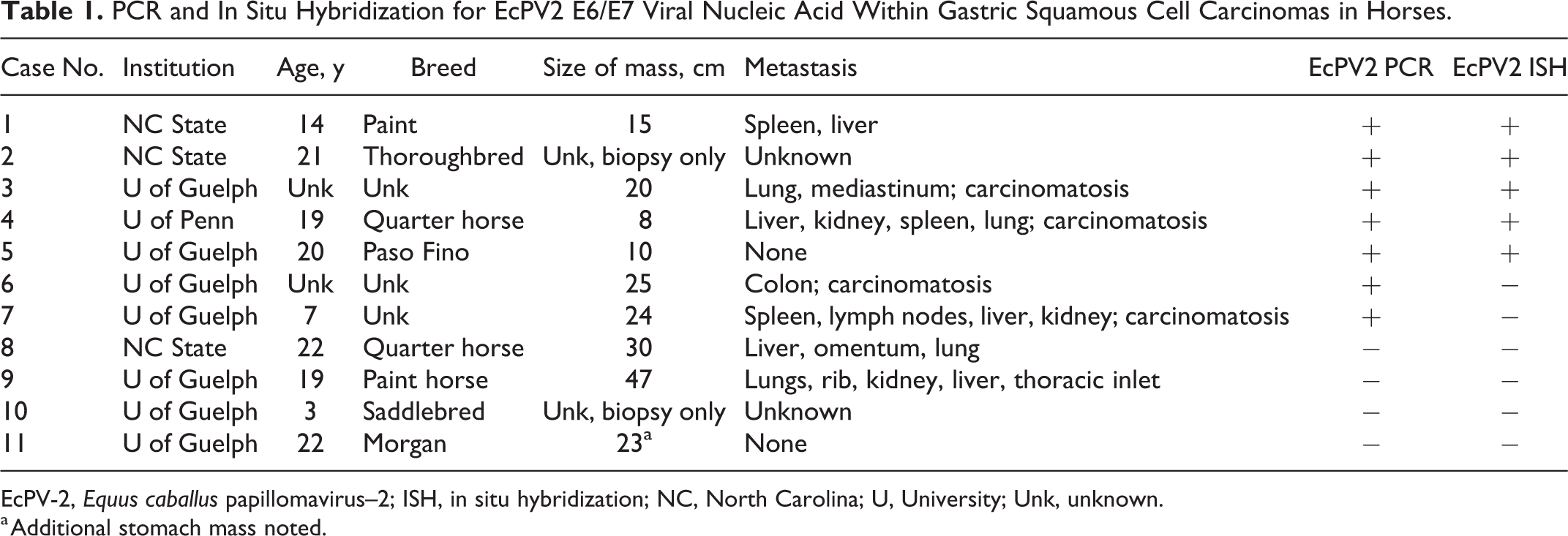
EcPV-2, Equus caballus papillomavirus–2; ISH, in situ hybridization; NC, North Carolina; U, University; Unk, unknown.
a Additional stomach mass noted.
Hematoxylin and eosin–stained slides were evaluated to confirm the diagnosis and further characterize the cases. Gastric SCC was characterized by infiltrative, expansile, and nonencapsulated masses of neoplastic keratinocytes arranged in islands, trabeculae, and cords supported by scirrhous stroma. The neoplasms variably merged with the overlying, hyperplastic squamous mucosa and frequently invaded the submucosa and tunica muscularis. Foci of keratinization and abortive cornification (keratin pearls) were often present. Central degeneration, lytic necrosis, and neutrophilic infiltration were common within islands of neoplastic tissue. Anisocytosis and anisokaryosis were mild to marked, and the mitotic count ranged from 1 to 11 per 10 400x fields (area per field: 2.37 mm2). Moderate numbers of lymphocytes and plasma cells infiltrated the supporting scirrhous stroma. Metastasis was observed in 7 cases, most commonly involving the liver (5), lung (4), peritoneal cavity (4), and spleen (3). Other sites of metastasis included the mediastinum (1), colon (1), kidney (2), thoracic inlet (1), omentum (1), lymph nodes (1), and rib (1). Microscopic evidence of vascular invasion was present in 2 of the primary SCC and 3 of the metastatic neoplasms.
PCR for the EcPV-2 E6 gene was performed on total DNA isolated from two 25-μm-thick scrolls of formalin-fixed, paraffin-embedded (FFPE) tissue samples using a Qiagen DNeasy Blood and Tissue Kit (Qiagen, Valencia, CA) following manufacturer’s recommended protocols. PCR primers for EcPV-2 E6 gene were designed using Primer 3 design software based upon the published EcPV-2 sequence between nucleotides 5 and 622 (Genbank reference sequence EU503122.1). Primers amplified a 226 base pair (bp) product and were as follows: EcPV-2 E6-For (5’-CAT TTG GTG GAA CAC AGC AG-3’) and EcPV-2 E6-Rev (5’-TTT TTC GTC CCT GGT CAG TC-3’). Primers for equine GAPDH were designed using Primer 3 and were as follows: Eq-GAPDH-For (5’-GAT TGT CAG CAA TGC CTC CT-3’) and Eq-GAPDH-Rev (5’-AAG CAG GGA TGA TGT TCT GG-3’). Equine GAPDH primers amplified a 196 bp product. PCR reaction mixtures contained 1 X buffer, 200 uM of each dNTP, 1uM each primer, 1U Taq Polymerase (HotStarTaq DNA Polymerase; Qiagen), and 10 uL (100 ng) total DNA in a 50 uL reaction mixture. A reaction without template DNA served as the negative control. PCR reaction conditions were as follows: 95°C for 15 minutes followed by 35 cycles of 95°C for 30 seconds, 58°C for 1 minute, and 72°C for 1 minute, followed by 72°C for 10 minutes. PCR products were run on a 1% agarose gel and visualized using SYBR Safe DNA gel stain (Invitrogen; Thermo Fisher Scientific, Pittsburgh, PA). EcPV-2 DNA was amplified from 7/11 (64%) gastric SCC cases but none of the non-SCC control cases (Table 1). GAPDH was amplified from all gastric SCC samples and all non-SCC control samples.
In situ hybridization was performed using a previously validated probe set designed to detect EcPV-2 E6/E7 nucleic acid in genital SCCs (RNAscope; Advanced Cell Diagnostics, Hayward, CA). 10 The probe is designed to detect a double-stranded DNA virus and may therefore detect both viral DNA and messenger RNA (mRNA). 10 ISH was performed on 5-μm-thick FFPE sections using the RNAscope 2.5 RED assay kit according to the manufacturer’s recommended protocols. Successful hybridization results in deposition of a red stain that is in direct correlation with the amount of EcPV-2 nucleic acid. A negative control was run concurrently using a predesigned probe to the bacterial gene DapB (ACD). One equine genital SCC was included as a positive control. The RNAscope 2.5 BROWN assay kit was used whenever the RED assay kit produced background staining on negative control sections. The BROWN assay kit produced superior results with reduced background on six cases.
Hybridization signals for EcPV-2 were detected within 5 of 11 gastric SCCs, including distant metastases (Table 1). Two of the 5 positive cases had overlying surface squamous epithelium to evaluate, which in both cases was characterized by mucosal hyperplasia (Figs. 1A and 2A; Suppl. Fig. 1A). These regions corresponded to intense nuclear staining for EcPV-2, predominantly within the upper layers of epithelium (Figs. 1B and 2B; Suppl. Fig. 1B). This distribution is congruent with the productive papillomavirus life cycle, where vegetative viral replication takes place within keratinocytes in the upper layers of the mucosal epithelium. 2 The presence of epithelial hyperplasia and intense nuclear ISH signals within the upper layers of epithelium suggests that SCC may develop from precursor hyperplastic lesions, as is the case for equine genital SCC and HPV-associated cancers. Less intense dot-like signals were present within the basal epithelium, where only mRNA transcripts for E6/E7 would be expected (Suppl. Fig. 1B). No hybridization signals were detected using the negative control probe (Fig. 2B inset; Suppl. Fig. 1C). Normal adjacent squamous epithelium was present within 2 of the cases positive for EcPV-2; within the normal epithelium, no EcPV-2 hybridization signals were detected.
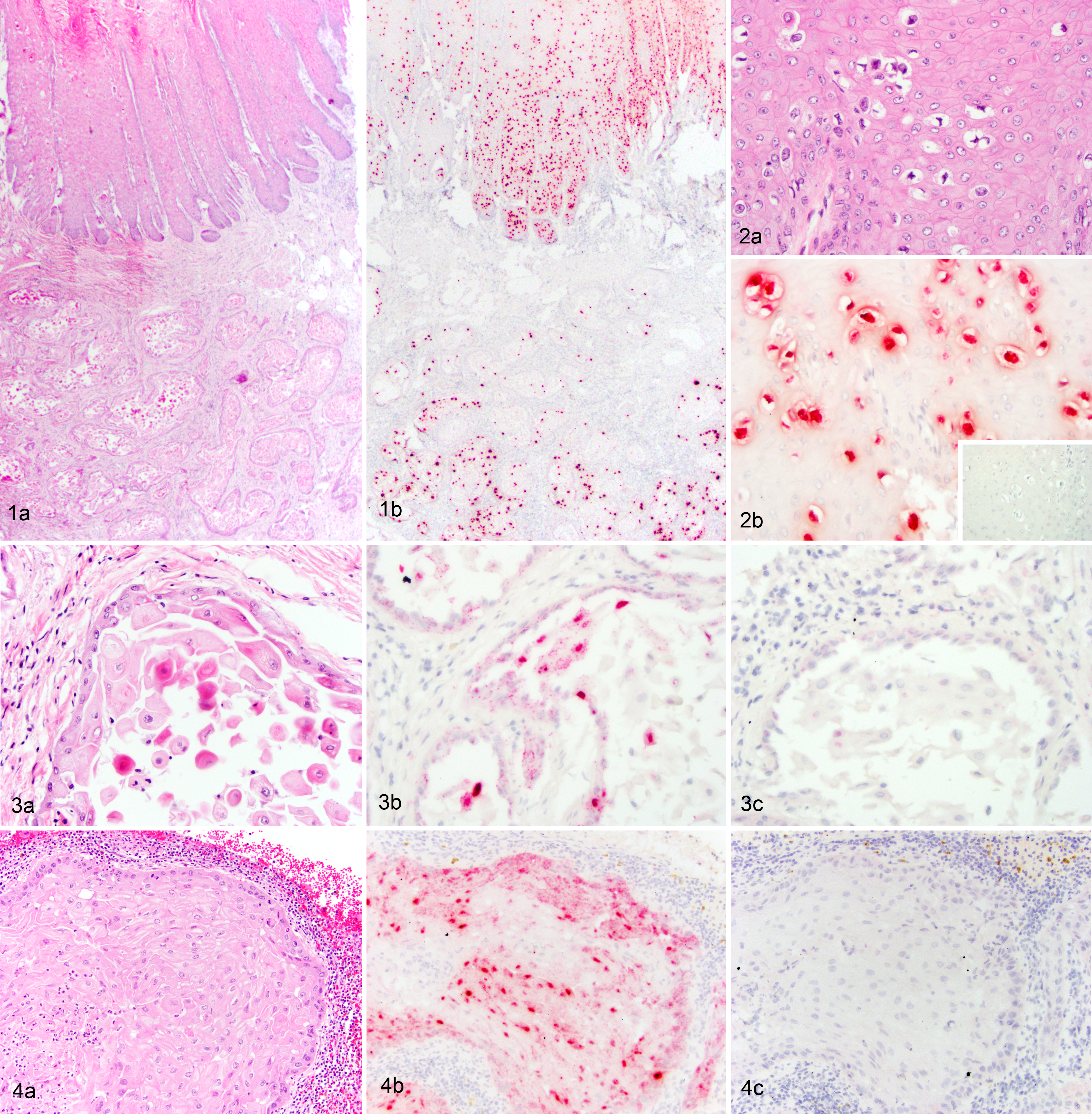
Invasive neoplastic cells contained variably intense dot-like hybridization signals within the nucleus and/or cytoplasm (Fig. 3A and B; Suppl. Figs. 2–5), occurring within 60%–100% of the neoplastic cells. No signals were seen above background staining levels using the negative control probe (Fig. 3C; Suppl. Figs. 2–5C). Metastatic lesions were present in 3 of the 5 positive cases; ISH signals for EcPV-2 were also detected in metastatic lesions and characterized by variably intense dot-like hybridization signals within tumor cell nuclei and/or cytoplasm (Fig. 4). Six gastric SCCs had no hybridization signals for EcPV-2 nucleic acid (Table 1). All non-SCC gastric samples were negative (Supp. Table 1).
All ISH-positive gastric SCC samples were also PCR-positive for EcPV-2. Two EcPV-2 PCR-positive gastric SCCs, however, were ISH-negative. While the cause for this discrepancy is unknown, it may reflect a difference in sensitivity between the 2 assays. Regardless, there were still several gastric SCC cases that were EcPV-2-negative. One possibility is that there is more than one mechanism for oncogenesis in gastric SCC, as has been suggested for genital SCC. 10 Alternatively, it is possible that EcPV-2 is an essential initiating oncogenic event but not required in later steps of malignant progression (“hit-and-run” hypothesis). This hypothesis has been suggested for development of human cutaneous SCCs. 2 In this scenario, EcPV-2 could be present within some tumors but lost within others.
While 30%–60% of human oropharyngeal SCCs are causally associated with HPV, human gastric SCCs are rare with an unknown etiopathogenesis. 1,5 There is no reported association with HPV. Suggested hypotheses for oncogenesis include tumors arising from regions of squamous metaplasia, transformation of a stem cell, or squamous differentiation within an adenocarcinoma. 1 We identified EcPV-2 nucleic acid within 45% of equine gastric SCC, which is closer to the percentage identified in human oropharyngeal SCC than gastric SCC. This difference could possibly be attributed to the anatomic differences between the human and equine stomach. While the human stomach is lined entirely by glandular epithelium, the equine stomach is composed of both a glandular and a nonglandular squamous portion; it is this squamous portion that gives rise to the gastric SCCs. 9 Given this distinction, it is possible the squamous portion of the equine stomach is more like an extension of the oropharyngeal macro- and micro-environments and thus susceptible to diseases more commonly seen within the oropharyngeal region. Provided that, however, EcPV-2 has only been detected within 3/20 (15%) and 4/15 (27%) oropharyngeal SCCs. 3,7 This lower detection rate within equine oropharyngeal SCC compared with gastric SCC could reflect the small sample size in these studies, differences in methodology to detect EcPV-2, differences in prevalence of EcPV-2, or even differences within oncogenic potential of EcPV-2 at these different sites.
In conclusion, this study provides support for a potential association between EcPV-2 and a subset of equine gastric SCC. Further studies are necessary to determine the role of EcPV-2 in development of gastric SCC.
Supplemental Material
Supplemental Material, Combined_supplemental_materials-Alloway_et_al - A Subset of Equine Gastric Squamous Cell Carcinomas Is Associated With Equus Caballus Papillomavirus–2 Infection
Supplemental Material, Combined_supplemental_materials-Alloway_et_al for A Subset of Equine Gastric Squamous Cell Carcinomas Is Associated With Equus Caballus Papillomavirus–2 Infection by Elizabeth Alloway, Keith Linder, Susan May, Trevor Rose, Josepha DeLay, Susan Bender, Alison Tucker and Jennifer Luff in Veterinary Pathology
Footnotes
Acknowledgements
We thank the NC State CVM and Rollins Histopathology Laboratories for their technical expertise, as well as Dr Murray Hazlett, AHL, University of Guelph, for his contribution to data searches.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Research reported in this publication was supported by the Office of Research Infrastructure Programs of the NIH under award number K01 OD023219-03.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.