
Abstract
RON is a tyrosine kinase receptor activated by the macrophage-stimulating protein (MSP) ligand that is overexpressed in human breast cancer. In humans, RON protein can be present in different isoforms, and the most studied isoform is represented by the short form of RON (sf-RON), which is generated by an alternative promoter located in intron 10 of the RON complementary DNA (cDNA). It plays an important role in breast cancer progression. Considering the many similarities between feline mammary carcinoma (FMC) and human breast cancer, the aim of this study was to investigate the expression of both RON and MSP in FMCs and to identify the presence of the sf-RON transcript. Tissue samples of spontaneous mammary tumors were collected from 60 queens (10 benign lesions, 50 carcinomas). All of the samples were tested for RON and MSP expression by immunohistochemistry; moreover, RNA was extracted from paraffin-embedded tissue samples, and the cDNA was tested by reverse transcription–polymerase chain reaction (RT-PCR) to identify the presence of sf-RON. Immunohistochemistry detected the expression of RON and MSP in 34 of 50 (68%) and 29 of 50 (58%) FMCs, respectively. RT-PCR revealed the presence of the short-form in 18 of 47 (38%) FMCs. This form originates, as in humans, from an alternative promoter (P2), and it codes for the proper feline short form (sf-RON). sf-RON expression was associated with poorly differentiated tumors and with a shorter disease-free (P < .05; hazard ratio [HR], 2.2) period and a shorter survival (P < .05; HR, 2.2). These results support FMC as a suitable model in comparative oncology and identify sf-RON expression as potential predictor of outcomes for this disease.
Since 1980, feline mammary carcinoma (FMC) has been recognized as a suitable animal model for studying human breast cancer since it shares histological, molecular, and clinical features with human breast carcinoma. 5,12,15,33
RON/stk proto-oncogene, a member of the MET family, is a tyrosine kinase receptor that is abnormally overexpressed in a variety of human cancers, including breast cancer. 21,30 The mouse homologue of human RON receptor, called stk, was identified in 1995, 16 while the feline homologue of the corresponding gene and protein, feline stk/RON, was first characterized in 2002. 11 Macrophage-stimulating protein (MSP) is RON’s specific ligand, and it is present both in tissue and in circulating blood. Downstream signaling targets of RON activity include PI3 K, Src, FAK, AKT, and MAPK, which can lead to proliferation, cell survival, cell motility, cell shape changes, and invasion in cancer. 9,10
Following alternative splicing, both the human RON and the homologous mouse stk genes can generate different isoforms of the receptor lacking the extracellular domain of the receptors but conserving its catalytic activity.
28,13
The 2 major transcripts are the full-length transcript that encodes the full-size RON receptor and the short form of RON (sf-RON), also called the truncated form. In humans, the full length of RON consists of 20 exons encoding a 180-kDa protein with 1400 amino acids while the sf-RON is generated by an alternative promoter (5′-UTR) located within intron 10 of messenger RNA (mRNA), regulating the transcription of sf-RON from the transcription initiation nucleotide represented by guanine
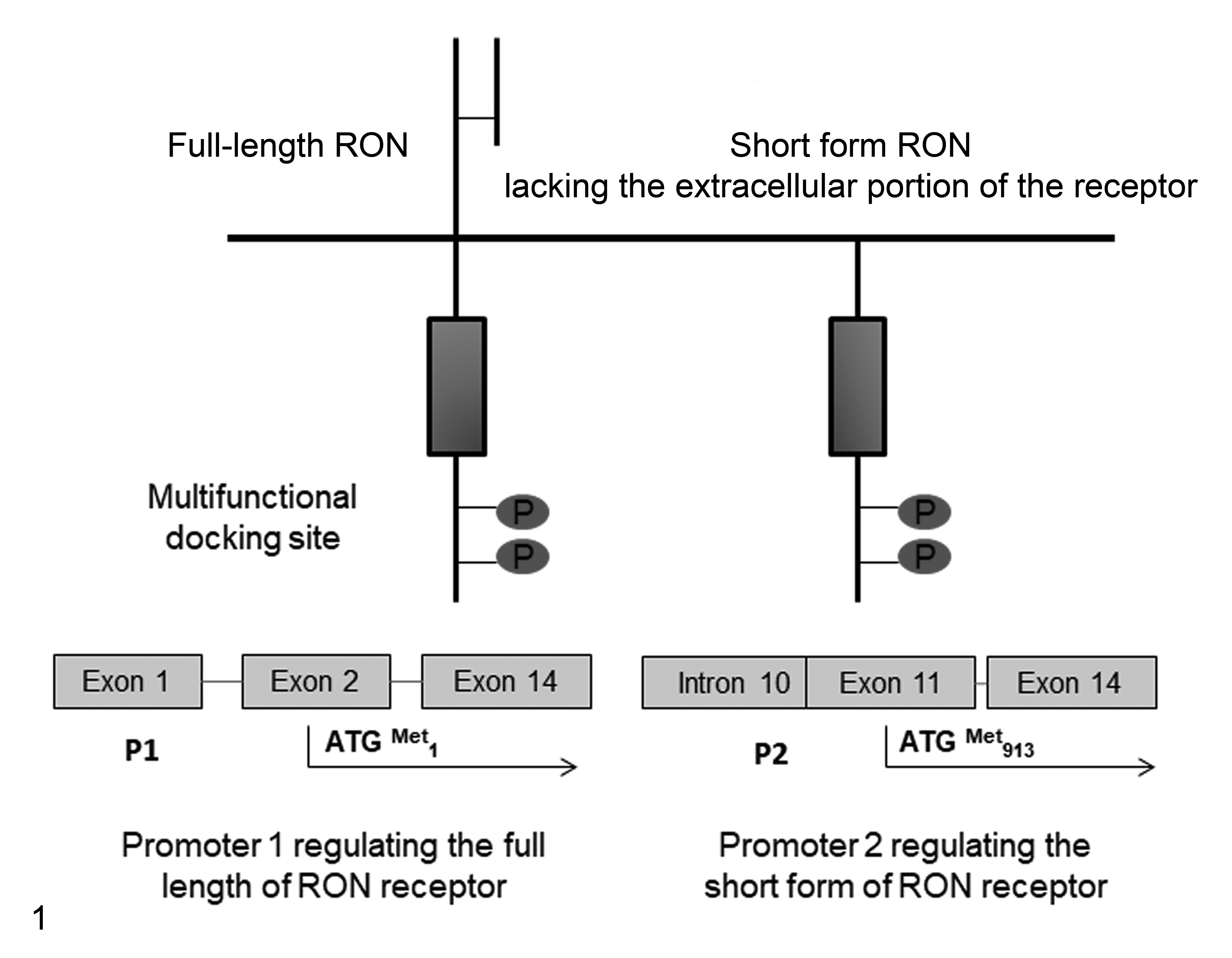
RON/stk gene structure and transcriptional regulation of the full-length and short forms lacking the extracellular domain of the receptor, respectively.
In humans, sf-RON was detected, at very low levels, in colon, skin, and lung tissue but not in bone marrow, bone marrow–derived cells, human muscle, or brain. 13,28 However, some reports have demonstrated the presence of sf-RON in cells derived from human breast cancer, lung cancer, pancreatic cancer, and leukemia. It has also been correlated with invasive capability in vitro, 3,4 and alternatively spliced RON transcripts have been reported in other types of cancers. 18 –20 In addition, it has been reported that sf-RON proteins are constitutively active since they are able to confer growth advantages to cells and have been involved in the epithelial-mesenchymal transition (EMT). 8
In breast cancer, the presence of sf-RON is correlated with a poor prognosis and tumor recurrence. 19 Moreover, the recent development of specific RON inhibitors in combination with PI3K inhibitors with demonstrated antitumor activity suggests an important role of sf-RON in terms of targeted therapies. 7,32
In consideration of the significant homologies between feline and human mammary tumors, the aim of this research was to investigate the expression of both isoforms of RON receptor and its ligand MSP in a large cohort of FMC tissues and cell lines and to identify the presence of the sf-RON transcript in FMC in relation to clinicopathologic findings.
Materials and Methods
Cell Lines
Six primary cell lines derived from FMC (FYCp, FKNp, FNNm, FMCp, FMCm, and P248) and spontaneously immortalized were tested in the in vitro experiments. 22,25,26,29 FYCp, FKNp, FNNm, FMCp, and FMCm were maintained in RPMI medium supplemented with 10% fetal bovine serum (FBS), 5000 IU/ml penicillin, 5 mg/ml streptomycin, and 2 mM L-glutamine; the P248 cell line was maintained in Dulbecco’s modified Eagle’s medium (DMEM) under the same conditions but with the addition of 10 mg/ml insulin to the medium. 23 The human GTL-16 gastric carcinoma cell line was used as a positive control in Western blot analysis and grown in DMEM as previously described. 31
Tissue Samples and Histological Examination
Tissue samples from spontaneous cases of feline mammary tumors and normal mammary tissues were collected from the 2009–2013 archives of the Diagnostic Laboratories of the Department of Veterinary Sciences of the University of Turin (Italy) and from the Department of Animal Pathology, University of Córdoba (Spain). Details of age, breed, ovariectomy status, and tumor size data were retrieved from the hospital database. Each animal was surgically treated with mastectomy (partial or total) and free surgical margins were histologically confirmed. This retrospective study was approved by the ethical committee of the Department of Veterinary Sciences of the University of Turin according to institutional guidelines.
Postoperative clinical, radiological, and echographic examinations at 6, 12, 18, and 24 months after surgery were performed to detect the presence of distant organ metastases or local relapse. Animals that died due to mammary carcinoma were submitted for postmortem examination to confirm the pathological diagnosis; subjects that died of nontumor-related causes (confirmed by necropsy) during the follow-up period were excluded from the study. The disease-free interval (DFI) was considered the number of days between surgery and tumor recurrence and/or evidence of metastatic disease, while overall survival (OS) was considered the period between surgery and death. Normal mammary tissues used as a control were collected from necropsy (after the owners’ consent). Samples were fixed in 4% neutral buffered formalin, paraffin embedded, sectioned at 4 μm, and stained with hematoxylin and eosin (HE). Samples were histologically examined and classified according to the World Health Organization (WHO) classification for tumors of domestic animals, 14 and surgical margins were evaluated to confirm that all tumor masses were completely surgically ablated. Carcinomas were graded according to Mills and colleagues. 24
Immunohistochemistry
Immunohistochemical (IHC) analysis was performed on 4-μm paraffin sections from all collected samples; immunocytochemistry was performed on all cultured cell pellets that were similarly fixed, embedded, and sectioned. After endogenous peroxidase activity blockade with 3% hydrogen peroxide for 20 minutes at room temperature, sections were subjected to high-temperature antigen retrieval by incubation with 98°C citric acid buffer (pH 6.0). Primary antihuman antibodies were used against RON β diluted 1:150, overnight incubation (C-20 rabbit polyclonal antibody, catalogue number sc-322; Santa Cruz Biotechnology, Dallas, TX) and against MSP diluted 1:50, overnight incubation (rabbit monoclonal antibody, catalogue number EPR6207; Abcam, Cambridge, UK). Antibodies were detected with avidin-biotin-peroxidase complexes using the Vectastain Elite ABC kit (Vector Laboratories, Burlingame, VT). Positive controls for RON and MSP were represented by lung and lymph node tissues, respectively. For the negative controls, positive control tissues were incubated with normal rabbit and mouse IgG (Santa Cruz Biotechnology). Moreover, to confirm the antibody specificity of MSP and RON antibodies, Western blot was assessed on protein lysates.
Immunostaining evaluation was performed independently by 2 observers (L.M. and S.I.); when there was disagreement, a consensus was obtained using a multihead microscope. Samples were scored semiquantitatively according to the intensity of membranous and/or cytoplasmic staining for both RON and MSP according to Ren and colleagues 27 (0 = no staining, 1+ = weak staining, 2+ = moderate staining, 3+ = strong staining). Samples with absent or weak immunostaining (score 0 and 1+) were defined as negative, and those with moderate to strong immunostaining (score 2+ and 3+) as positive. In the Supplemental Figures S1 to S8, examples of each score are shown.
Protein Extraction and Western Blot Analysis
Total proteins were extracted from 3 cases of feline mammary carcinomas (FMC1, FMC2, and FMC3) collected at the Veterinary Hospital of the Department of Veterinary Science of Turin and from the FYC cell line; normal feline spleen tissue from necroscopy was used as a negative control while protein lysate from GTL16 was used as a human positive control. Total proteins were extracted with fresh RIPA buffer solution (150 mM sodium chloride, 1.0% Triton X-100, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate, 50 mM Tris, pH 8.0) containing a protease inhibitor cocktail (Sigma-Aldrich, St Louis, MO). In total, 200 μg of total protein from each sample was separated on 9% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) gels, transferred onto Hybond-C Extra membranes (Amersham Biosciences, Bath, UK), and incubated overnight at 4°C with the same primary antibody against RON used in immunohistochemistry. After washing 3 times in phosphate-buffered saline (PBS)–Tween for 15 minutes and incubating with a horseradish peroxidase (HRP)–linked secondary antibody diluted 1:2000 in PBS-Tween 0.01%, the membranes were washed in PBS-Tween for 30 minutes and then incubated with an enhanced chemiluminescence reagent (Super Signal West Pico Rabbit IgG Detection Kit; Thermo Scientific, Waltham, MA). To confirm also the specificity of MSP antibody used in immunohistochemistry, total proteins extracted from feline cell lines (FMCp, FMCm, and FNN) were subjected to Western blot against MSP (dilution 1:1000) as previously described.
DNA Extraction and Sequencing of Genomic Feline RON Exons 10–14
Genomic DNA was extracted from normal feline lymphoid tissue (spleen) using a QIAamp DNA mini kit (Qiagen, Hilden, Germany). Polymerase chain reaction (PCR) was performed on 100 ng total genomic DNA using HotStart AmpliTAQ (Qiagen), and amplification of the genomic region of the feline RON corresponding to exons 8 to 14 was performed. Sequences of exonic primers P1 and P2 are shown in Supplemental Table S1 and were designed on the basis of the sequence of feline RON complementary DNA (cDNA) present in the gene bank (XP_003982293). The PCR protocol for all primers was as follows: 1 cycle at 95°C for 5 minutes, 30 cycles (95°C for 1 minute, 60°C for 1 minute, 72°C for 1 minute), and 1 cycle at 72°C for 7 minutes. After purification of specific amplified bands from agarose by the QIAquick gel extraction kit (Qiagen), direct sequencing of PCR products was performed using the BigDye Terminutesator V 3.1 Sequencing Kit and ABIPRISM 310 Genetic Analyzer (Applied Biosystems, Foster City, CA).
cDNA Preparation, PCR, and Sequencing of sf-RON
Total RNA was extracted from paraffin-embedded tissues of FMCs, benign lesions, and normal mammary glands using an RNA FFPE extraction kit (Qiagen) and from 6 FMC cell lines (FYCp, P248 m, FKNp, FNNm, FMCp, and FMCm) using the TRIzol method (Sigma-Aldrich). The quality of the RNA was evaluated using the Experion Automated Electrophoresis System (Bio-Rad, Hercules, CA), and samples with degraded RNA were excluded. cDNA was synthesized from 1 μg total RNA using the QuantiTect Reverse Transcription kit (Qiagen), including genomic DNA removal as described in the procedures of the kit. To examine the RON transcript, end-point reverse transcription–PCR (RT-PCR) was performed with primer P3 sense and P4 antisense (Suppl. Table S1) in a final volume of 20 μl using HotStart TAQ (Qiagen). The PCR conditions were as follows: 1 cycle at 95°C for 5 minutes, 30 cycles (95°C for 1 minute, 60°C for 1 minute, 72°C for 1 minute), and 1 cycle at 72°C for 7 minutes. Moreover, to exclude genomic DNA contamination, PCR of intronic sequences corresponding to introns 12 and 13 of the feline RON gene was performed on cDNA (data not shown).
Amplification products obtained from RT-PCR were extracted from the agarose by a QIAquick gel extraction kit (Qiagen) and directly sequenced using a BigDye Terminator V 3.1 Sequencing Kit and an ABI PRISM 310 Genetic Analyzer (Applied Biosystems). Amplification of feline GAPDH was used as the housekeeping gene. The presence of CpG islands was assessed, and we submitted the sequence to the online software available at http://dbcat.cgm.ntu.edu.tw/.
Statistical Analysis
IHC results, molecular data, and clinicopathologic findings were grouped into contingency tables and analyzed using Fisher’s exact test or χ2 test. Survival curves were computed using the Kaplan-Meier method, and tests for differences in survival, considering all known prognostic factors for FMC, were performed using the log-rank test. OS was defined as the number of days between surgery and death, while the DFI was defined as the number of days between surgery and tumor recurrence and/or evidence of metastasis. Censored data were considered as those cases that were still alive or that did not present with tumor recurrence or metastasis at the end of the monitoring period. Data were analyzed with MedCalc Statistical Software v.13.3 (MedCalc Software bvba, Ostend, Belgium); P < .05 was considered statistically significant.
Results
Clinical and Pathologic Data
Sixty queens with mammary lesions were enrolled in this study. Their mean age was 11.3 years (range, 8–17 years); 20 of 60 were spayed (before 2 years of age) and 40 of 60 were not. Forty of 60 were European Shorthair, 16 of 60 were European longhair, and 4 of 60 were Siamese breed. Histological examination resulted in 50 FMCs and 10 benign lesions (Suppl. Table S2). Four normal mammary tissues (from archives) were also evaluated as controls.
Regarding queens with a diagnosis of FMC, at the time of diagnosis, histologically confirmed lymph node metastases were present in 2 of 50 (4%) cases, and no clinically or radiographically observable distant organ metastases were present; 10 of 50 (20%) queens did not present with tumor recurrence while 40 of 50 (80%) relapsed between 2 and 23 months after surgery. The median OS recorded was 180 days (range, 45–730 days). Forty of 50 cases developed pulmonary metastases during the follow-up period and died of cancer-related causes as confirmed at postmortem examination. Ten censored cases were included in this study.
Immunohistochemistry
Immunoreactivity to MSP (Figs. 2, 4) and RON (Figs. 3, 5) proteins, if present, were observed in the cytoplasm of FMC cell lines and carcinomas. IgG control staining is shown in Supplemental Figure S9. Immunoreactive products were not observed in normal mammary glands (Suppl. Fig. S10).
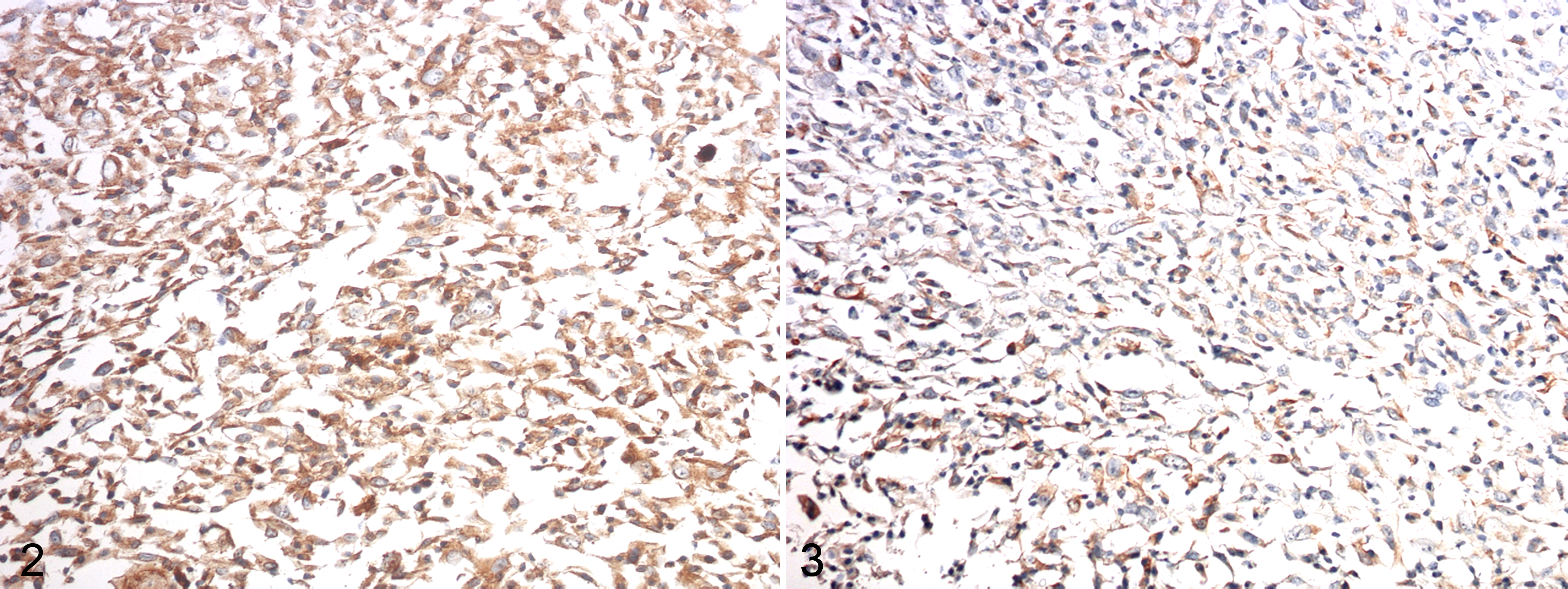
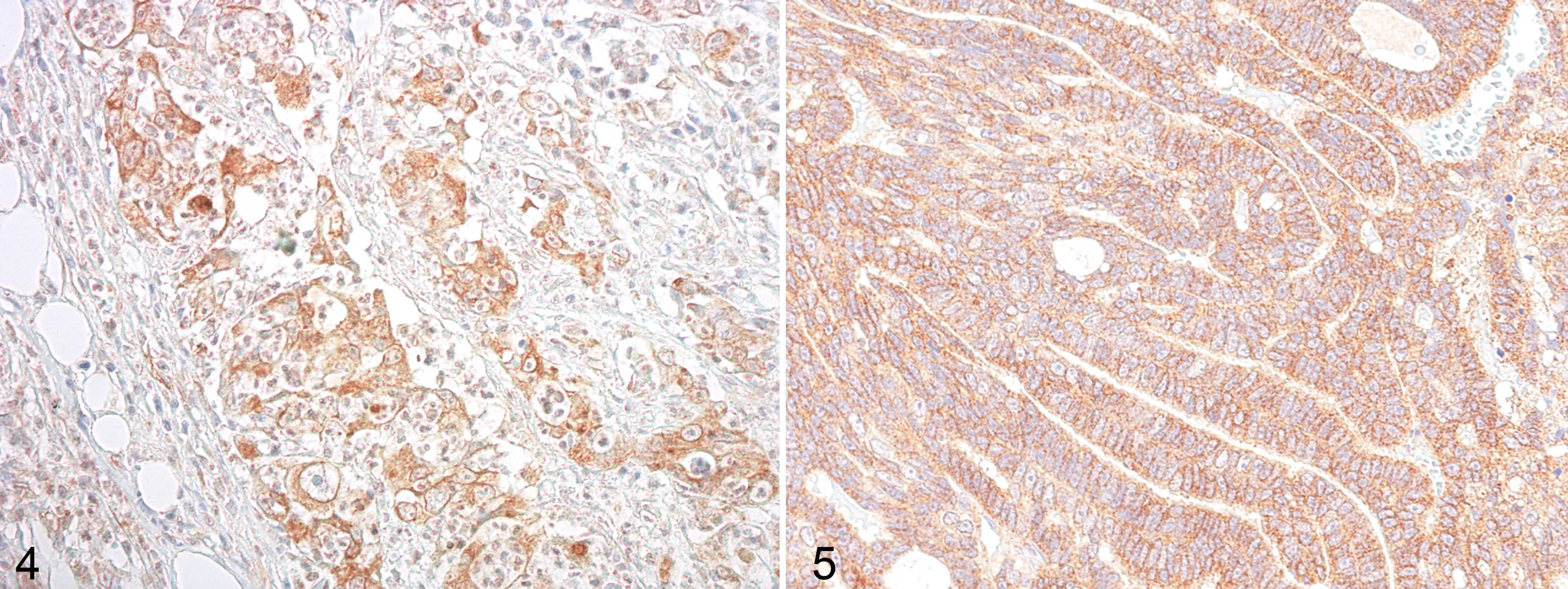
Regarding the expression of RON and MSP in benign lesions, 3 of 10 (2 adenomas and 1 fibroadenoma) were positive for RON, while 2 of 10 (1 adenoma and 1 fibroadenoma) were positive for MSP. In FMCs, RON was positive in 34 of 50 (68%) of cases, and MSP was positive in 29 of 50 (58%) of cases. A total of 52% of FMCs expressed both RON and MSP, and a statistically significant association was found between the expression of these 2 antibodies (P < .05).
The specificity of RON and MSP antibodies was confirmed by Western blot analysis (Suppl. Fig. S11).
No statistical correlation was found between the immunohistochemical results for RON and MSP in relation to the clinicopathologic findings in the 50 FMC cases (Table 1).
Relation Between Clinicopathologic Features and Immunohistochemical Results.
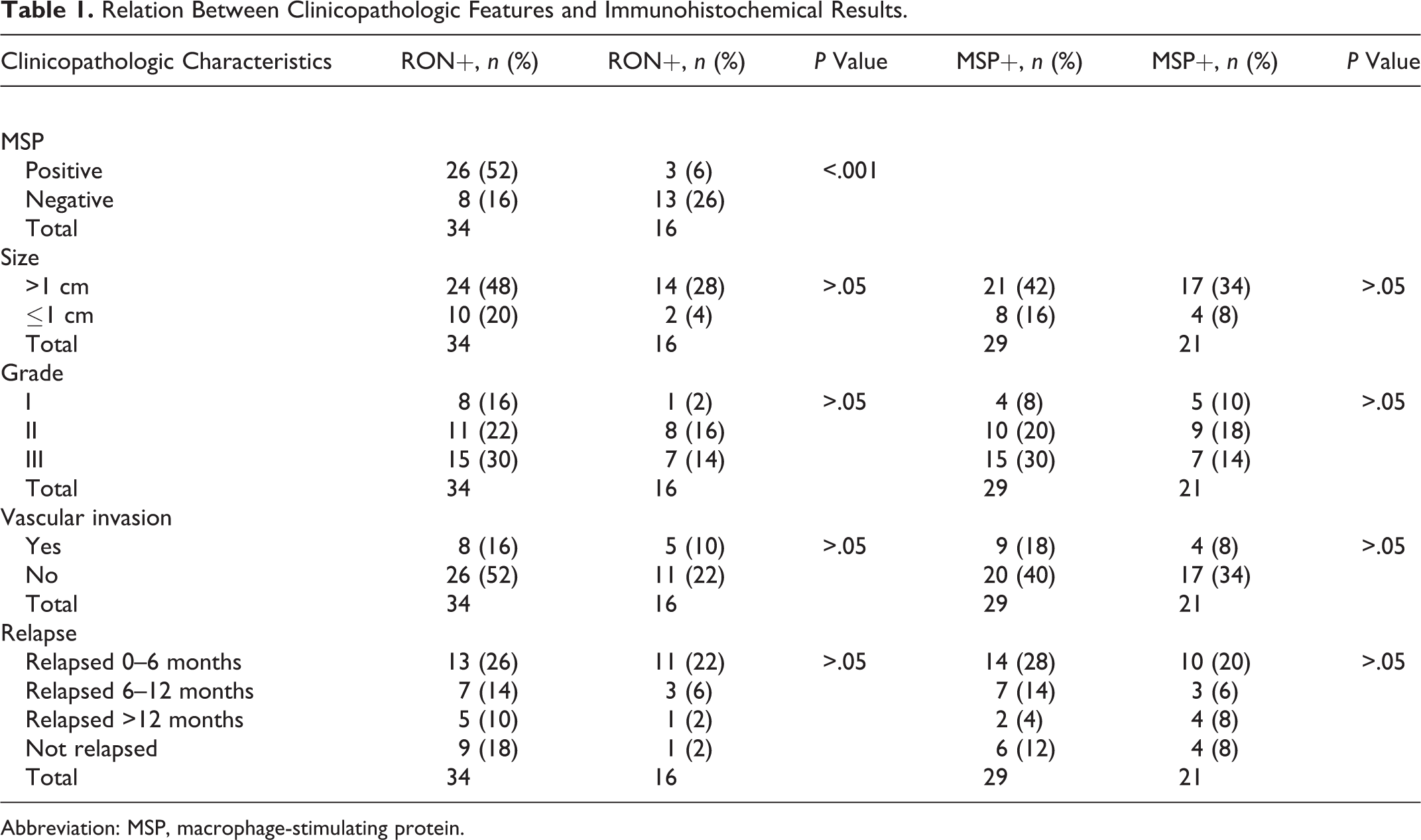
Abbreviation: MSP, macrophage-stimulating protein.
Genomic Sequence of Feline RON
Sequences of DNA coding for feline RON from exons 10 to 14 were obtained from normal spleen, and the sequences of the gene, mRNA, and CDS (coding sequence) with exon-intron junctions were submitted to GenBank with the accession number KT248534.
Identification of Feline sf-RON
To examine the presence of sf-RON in our samples, RT-PCR was performed with P1 and P2 primers designed to amplify exon 10 or exon 11 of the feline cDNA. PCR products obtained from the amplification showed 2 bands, the higher band containing intron 10 (size 94 bp) in the cDNA sequence and comigrating with normal feline DNA (size 350 bp), as well as a lower band containing the full-length feline RON but not containing intron 10 with a corresponding size of 256 bp. Both of the bands obtained were sequenced and revealed that the highest band corresponded to sf-RON matching at 100% with the genomic sequence previously submitted in Gene Bank (accession number KT248534). Figure 6 shows the rationale used to amplify sf-RON.
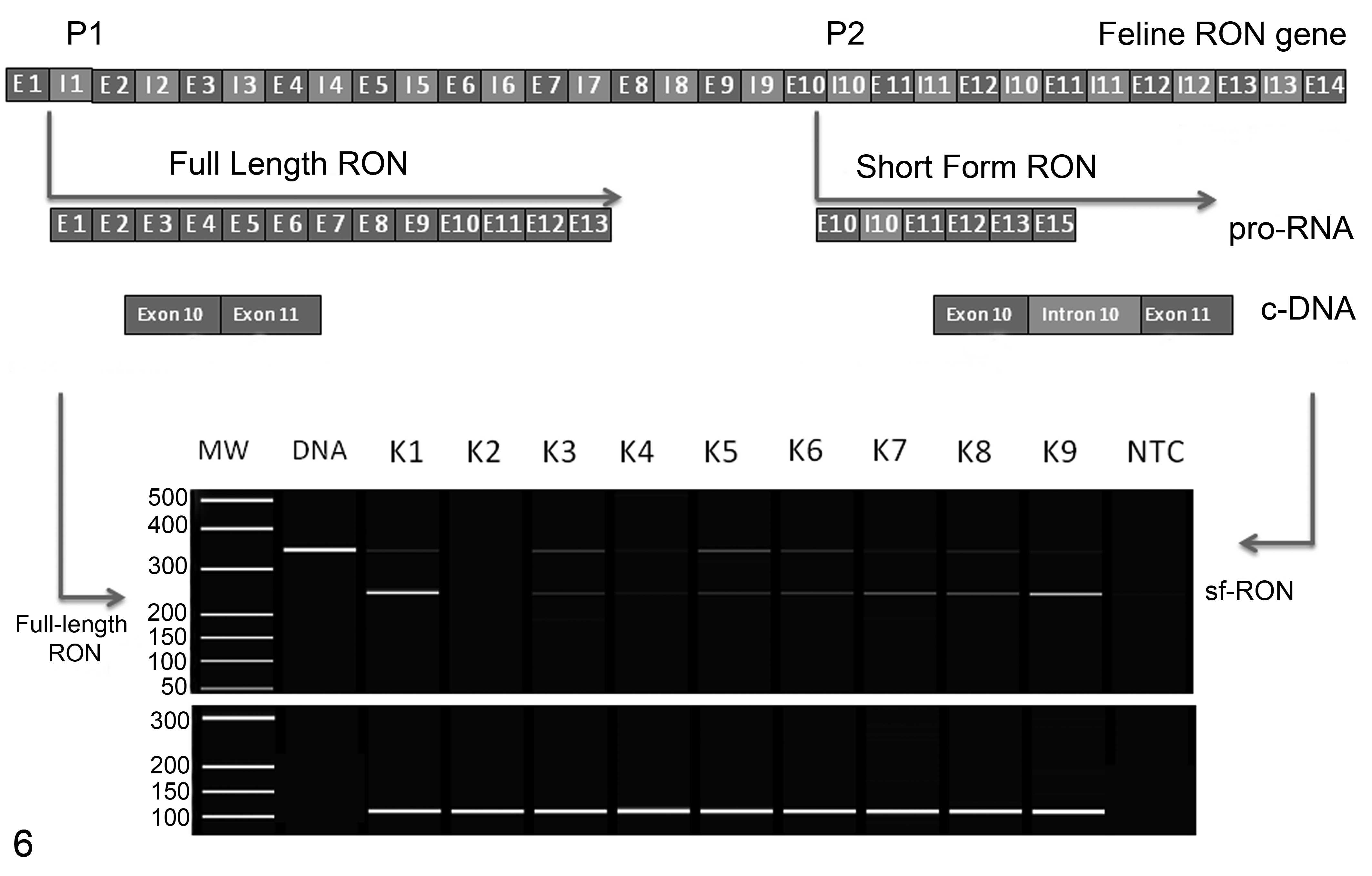
Regulation of the expression of full-length and short forms of feline RON gene receptor and reverse transcription–polymerase chain reaction products obtained from feline mammary carcinomas. The higher and lower bands correspond respectively to short and full-length forms of RON. K1 to K7 correspond to feline mammary carcinomas while NTC is the nontemplate control. GAPDH was used as the housekeeping gene control (lower panel).
Using this RT-PCR assay, sf-RON was detected in 18 of 47 (38.3%) FMCs (3/50 cases were excluded from analysis because the RNA was degraded) and in 4 of 6 cell lines. Benign tumors and normal mammary gland tissue did not express feline sf-RON. All bands obtained were automatically sequenced to confirm the presence of sf-RON in 18 of 47 samples. In Supplemental Figure S12, RT-PCR products obtained from cDNA templates from the cell lines are shown.
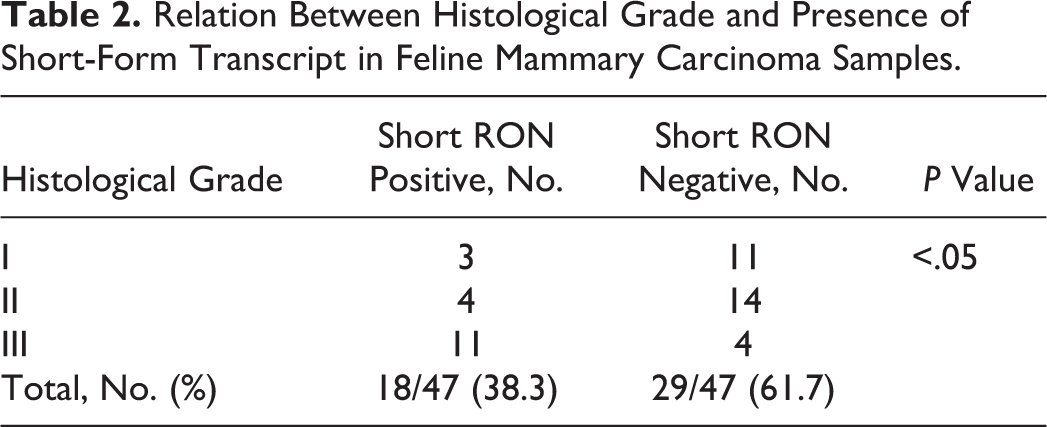
Statistical analysis revealed that queens with FMC that were positive for sf-RON had both a shorter DFI (median 180 days vs 635 days; hazard ratio [HR], 2.2; 95% confidence interval [CI], 1.04–4.6) and a shorter OS (median 365 days vs 755 days; HR, 2.2; 95% CI, 1.03–4.7) compared with queens with tumors negative for sf-RON (Figs. 7, 8). Moreover, FMC positivity for sf-RON was statistically associated with poorly differentiated histological grade (Table 2).
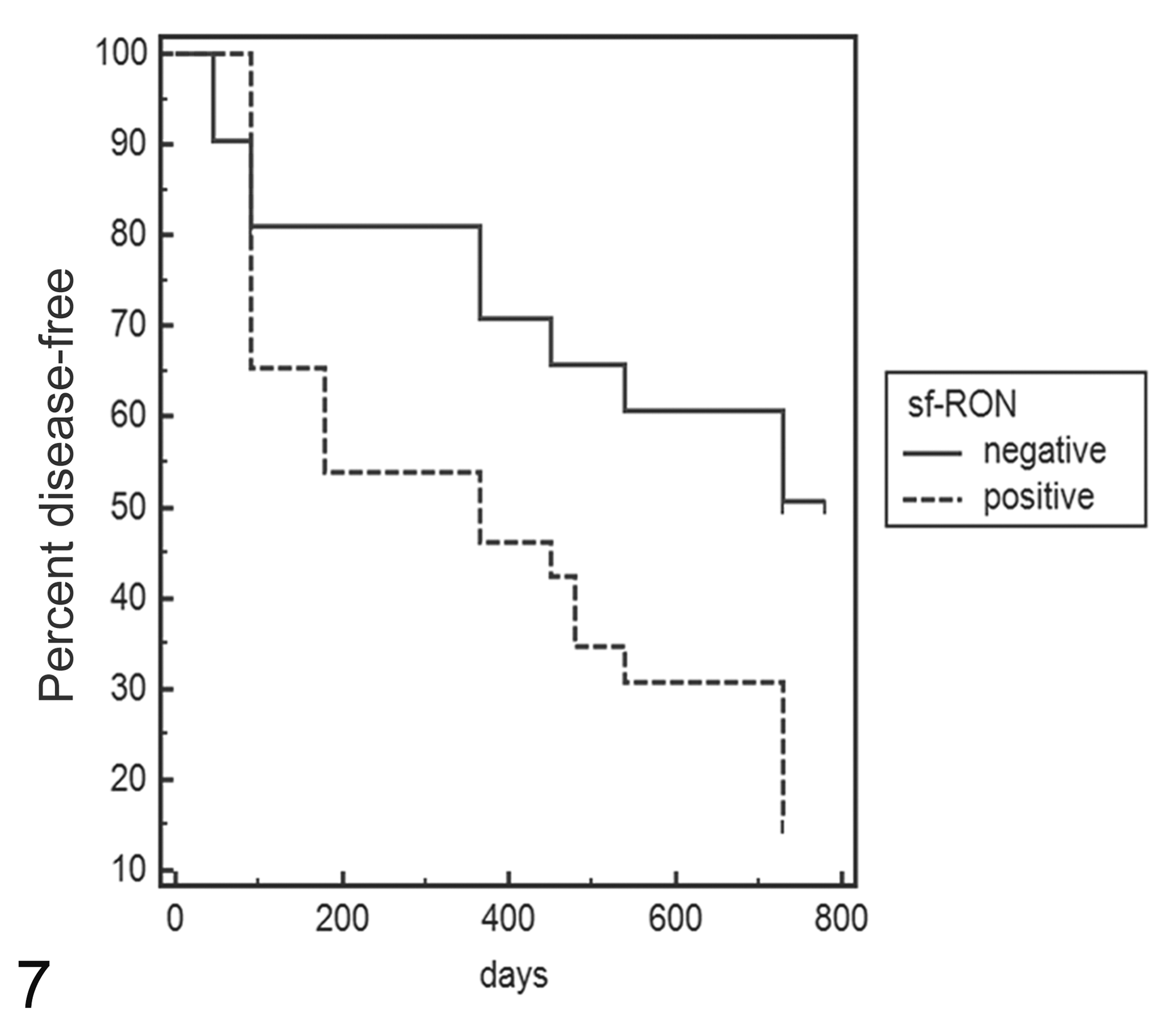
Kaplan-Meier survival curve of disease-free interval (DFI) in feline mammary carcinomas. Total mean DFI: 407 days; mean sf-RON–positive samples: 326 days; mean sf-RON–negative samples: 510 days (log-rank test: P < .05). The data are based on expression of sf-RON as determined by reverse transcription–polymerase chain reaction.
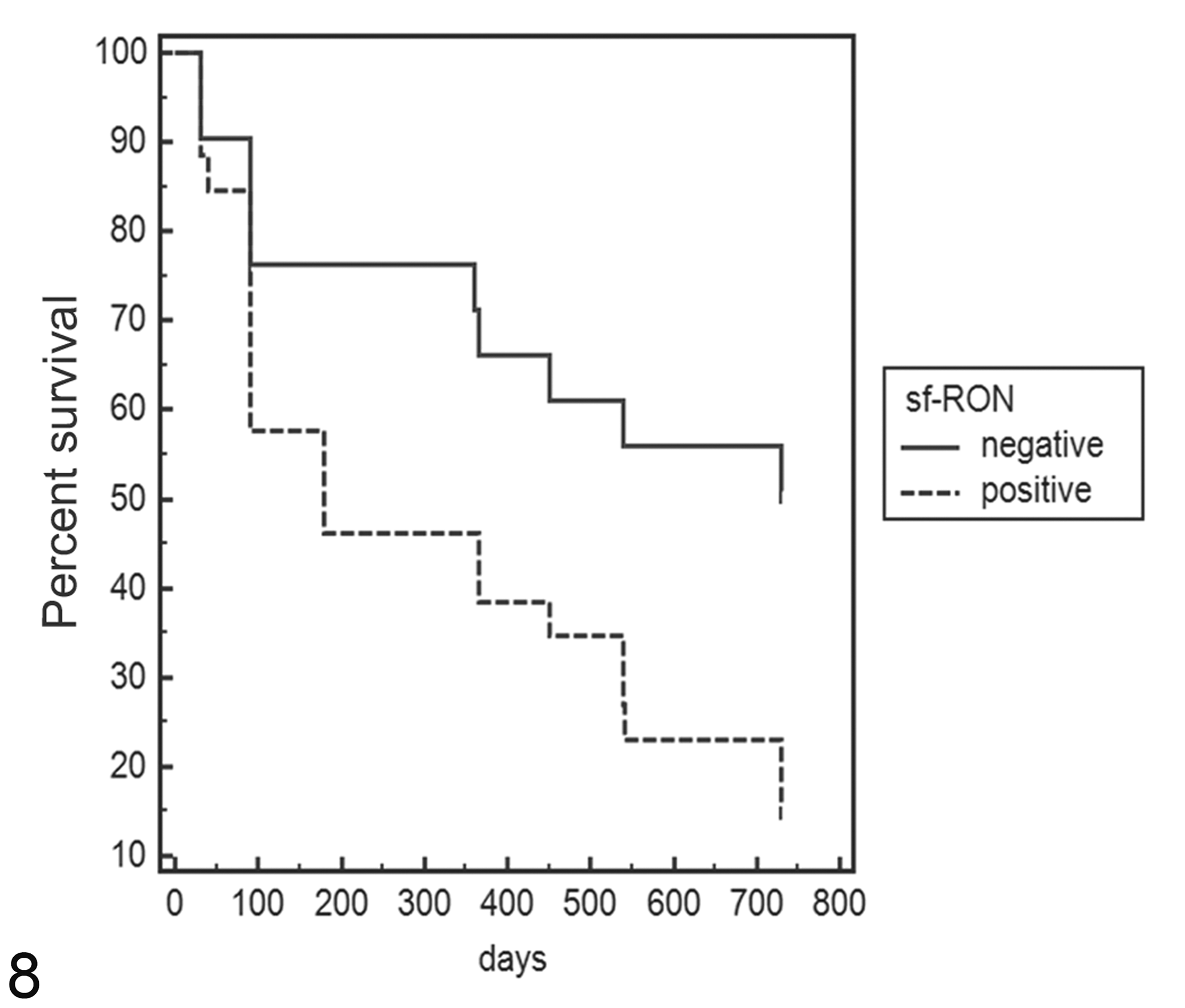
Kaplan-Meier survival curve of overall survival (OS) in feline mammary carcinomas. Total mean OS: 466 days; mean sf-RON–positive samples, 387 days; mean sf-RON–negative samples, 568 days (log-rank test: P < .05). The data are based on expression of sf-RON as determined by reverse transcription–polymerase chain reaction.
Relation Between Histological Grade and Presence of Short-Form Transcript in Feline Mammary Carcinoma Samples.
To demonstrate if intron 10 in felines also contained typical CpG island regions acting as alternative promoter regions, the entire sequence was analyzed using software available at http://dbcat.cgm.ntu.edu.tw/. From this analysis, we found that the region corresponding to intron 10 contained more than 66% of the CpG islands (Suppl. Fig. S13).
Protein Analysis
Feline mammary carcinoma samples (FMC1, FMC2, and FMC3) corresponding to poorly differentiated (FMC1 and FMC2) and moderately differentiated mammary carcinomas (FMC3) were collected. Protein analysis revealed that all FMCs examined and the FYC cell line expressed the precursor of RON at different levels, resulting in a protein of 180 kDa comigrating with the human control represented by the GTL16 cell line (Fig. 9). As shown, the β chain isoform in humans corresponds to 150 kDa, while in feline samples, it corresponds to 135 kDa, as previously demonstrated by the same authors, 11 and it was present in all analyzed FMCs at different levels.
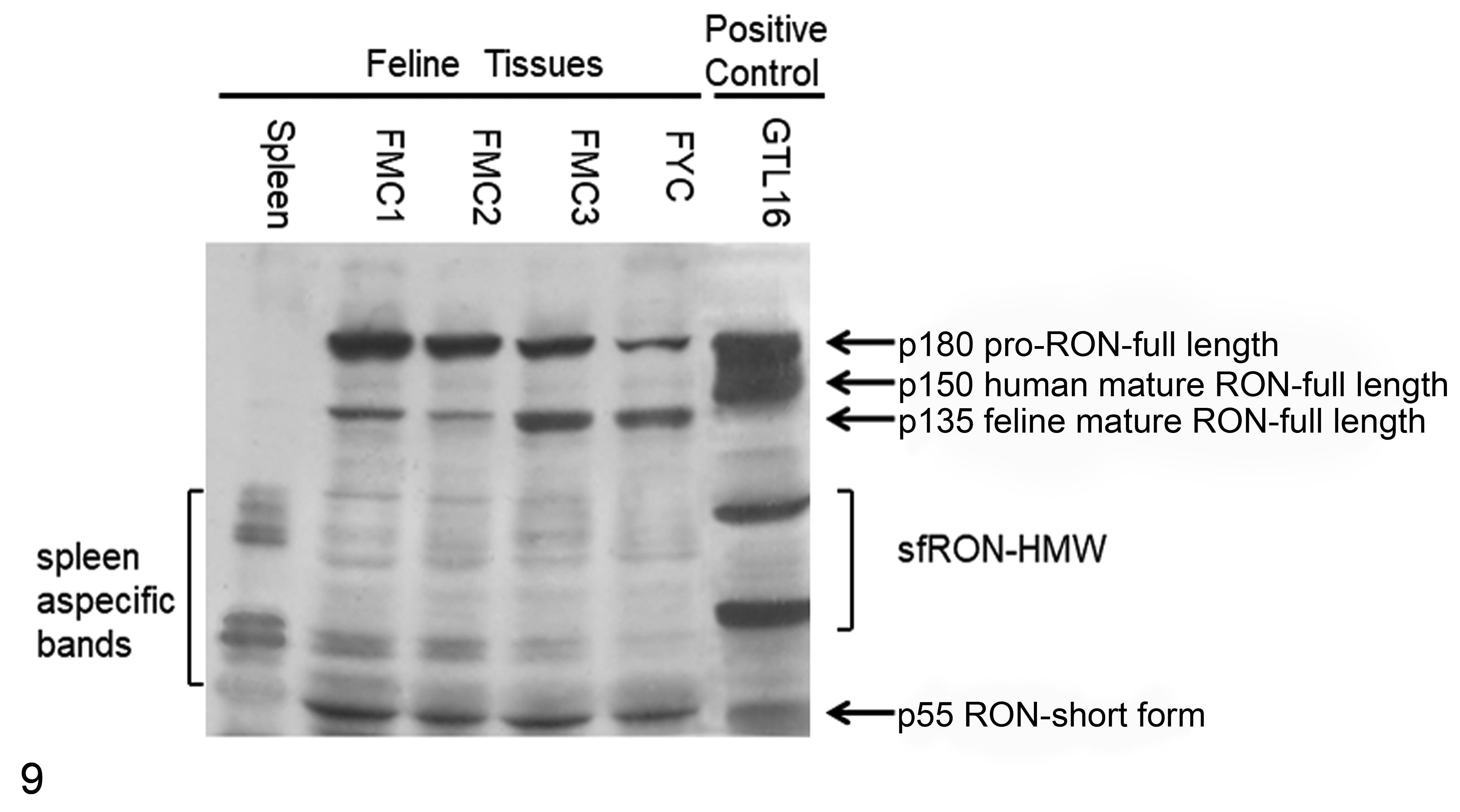
Western blot against RON of protein lysates from 3 feline mammary carcinomas (FMC1, FMC2, and FMC3), FYC cell line, feline spleen, and the human GTL16 cell line. fl-RON and sf-RON are present in all feline mammary carcinoma samples at different levels. Feline spleen represents the negative control and shows nonspecific bands. GTL16 represents the human positive control. HMW, high molecular weight.
Conversely, in all samples, a specific band corresponding to 55 kDa was detected, confirming the presence of the short form in feline samples and in the FYC cell line that expresses sf-RON, as also shown in RT-PCR (see Suppl. Fig. S12). Feline spleen gave negative findings for both isoforms (Fig. 9). The presence of additional protein bands migrating at 72 to 90 kDa should be referred to as sf-RON–high molecular weight (sf-RON–HMW) as already demonstrated in human breast cancer, 19,20 which is recognized by the same antibody.
Discussion
In this article, we provide evidence that the truncated form of RON kinase is expressed in feline mammary tumors and that its expression represents a prognostic factor in this type of tumor. The presence of the RON transcript in feline mammary tumors was first demonstrated in 2002 when the authors partially identified the gene coding for full-length RON protein. 11 In humans and mice, the gene coding for sf-RON is regulated by an internal promoter (P2) located in intron 101 as shown in Figure 1. To demonstrate if this isoform was also present in cats, the authors first determined the genomic sequence of feline RON from exon 8 to exon 14 and its intron/exon junctions. These data showed high homology with human, canine, and murine sequences and were submitted to the GenBank database. On the basis of the feline intron 10 sequence and adopting the molecular approach published by Bardella and colleagues, 3 the authors evaluated the presence of both sf-RON by end-point RT-PCR in cDNA from FMC cell lines and the RON protein from tissue samples from feline mammary tumors. The results obtained from RT-PCR on tissues demonstrated the presence of sf-RON in 38.3% of feline mammary carcinomas and in 4 of 6 feline mammary cell line transcripts. In all of the samples, the sequence of amplicons revealed the insertion of intron 10 (P2). Moreover, in benign lesions and normal mammary glands, sf-RON was not detected. These results were confirmed by Western blot analysis of FMC samples and the FYC cell line. In Figure 9, all samples, at different levels, showed the mature form of the feline RON migrating at 135 kDa, as previously demonstrated by the same authors, 11 while sf-RON was detectable at 55 kDa in feline FMCs, FYC cell lines, and human controls represented by GTL16; feline spleen was negative for both isoforms. In this experiment, the authors also detected protein products migrating at 72 to 95 kDa. These proteins, as demonstrated in human breast cancer, 19 can be referred to other isoforms called sf-RON–HMW, derived from further different splicing of the human RON gene that can generate protein products of approximately 72 to 95 kDa. 20 The sf-RON–HMW isoforms were identified in human breast cancer tissues using the same antibody by Liu et al, 19 and this suggests that the presence of these isoforms should be investigated in cats.
Overall, these data suggest that in cats as in humans, sf-RON originates from the alternative promoter (P2) located in intron 10. In particular, analyzing the nucleotide sequence “corresponding” to intron 10, we found a high percentage of possible CpG islands with a GC content of 66%, suggesting, as in humans, that RON gene transcription may be regulated by a promoter in the CpG islands region. Moreover, the presence of sf-RON was significantly associated with a poorly differentiated histological grade, with a shorter disease-free (DFI) period and a shorter survival (OS). Regarding this aspect, there are no data available as to the prognostic value of sf-RON in human patients with breast cancer. Otherwise, our results are according to literature where the presence of sf-RON confers aggressiveness of human breast cancer in in vitro and in vivo conditions, and it is able to convert slow-growing and nonmetastatic tumors into rapidly growing tumors and promotes EMTs via PI3K pathways, 19 confirming the importance of this protein in breast cancer progression, as already demonstrated in feline mammary carcinomas. 22,23
Immunohistochemical expression of the RON receptor and its specific ligand MSP was observed in 68% and 58% of FMCs, respectively, while 52% of cases coexpressed both proteins. However, no correlation was observed between immunohistochemical expression of the RON receptor or its ligand MSP with clinicopathologic and survival data. The antibody used in this study for the detection of RON protein (C-20) recognizes the COOH terminus of the protein (intracellular portion), which is present in both isoforms (full and short). Thus, IHC data could not differentiate between the 2 isoforms in tissues. These considerations could explain the lack of correlation between IHC expression of RON/MSP and the clinical outcomes compared with expression of sf-RON assessed by RT-PCR.
Our IHC data confirm our previous results in feline mammary tumors 11 and are similar to the human literature where RON is expressed in 50% of primary breast carcinomas but does not represent a prognostic factor. 21
No data are available in human breast cancer regarding the association between the presence of sf-RON and the clinical findings, although it is known that this isoform, through its interaction with PI3K, is able to promote metastasis. 19
The relevance of sf-RON in human cancer is of emerging interest due to the development of specific molecules able to inhibit RON receptor activity. 34 The potential use of molecules able to inhibit RON has been recently studied in cancers of the pancreas, lung, colon, and breast. 6,17,18,35 As previously discussed, FMC highly resembles human breast cancer, sharing histopathological, clinical, and molecular features. 12,15,33 In this study, we demonstrated that feline mammary tumors and their derived cell lines present with the truncated form of RON, and its presence is significantly associated with high grade of tumor and a worse prognosis.
As shown in Figures 7 and 8, after 100 days, there was no apparent effect of sf-RON on the DFI and OS, but its effect seems to appear in a later phase. This tendency suggests that in the early stages of tumor progression, sf-RON is not involved, but in the later stages, further additional pathways participate in tumor progression. These results support FMC as a suitable model in comparative oncology and identify sf-RON expression as having potential in predicting the outcomes of this disease.
Supplemental Material
Supplemental Material, DS1_VET_10.1177_0300985818806967 - Expression of the Short Form of RON/STK in Feline Mammary Carcinoma
Supplemental Material, DS1_VET_10.1177_0300985818806967 for Expression of the Short Form of RON/STK in Feline Mammary Carcinoma by Lorella Maniscalco, Silvia Guil-Luna, Selina Iussich, Francesca Gattino, Calogero Trupia, Yolanda Millan, Juana Martín de Las Mulas, Raquel Sanchez Cespedez, K. Saeki, Paolo Accornero, and Raffaella De Maria in Veterinary Pathology
Footnotes
Acknowledgements
We thank Alessandra Sereno, Rocchina Evangelista, and Gema Muñoz, School of Veterinary Medicine, University of Turin, Italy, for their technical support.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Financial support was partially supplied by grant AGL 2011-25553 from the Spanish Ministry of Research, Spain.
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.