
Abstract
Idiopathic chronic diarrhea (ICD) is a common ailment affecting captive rhesus macaques (Macaca mulatta). ICD cases are characterized by diarrhea in the absence of commonly identified diarrheal pathogens and multiple recurrences even after supportive therapy. Histologically, the disease is characterized by lymphoplasmacytic colitis. We identified 35 rhesus macaques euthanized for ICD during a 7-month period and described demographic, clinical, histologic, and immunologic commonalities. We found a trend of historic Campylobacter spp. and trichomonad infections. Furthermore, rhesus macaques with ICD demonstrated loss of normal colonic adherent bacterium, identified in this study as Helicobacter macacae; increased abundance of Pentatrichomonas hominis; and increased frequency of colonic serotonin-positive enterochromaffin cells. Interestingly, colonic and ileal T-helper cells of animals with ICD manifested decreased capacity for expression of certain cytokines, in particular interleukin (IL)–4 and IL-13. These data further describe a common ailment and suggest new avenues to identify complex interactions involved in the etiology of recurring diarrhea in young rhesus macaques.
Keywords
Chronic diarrhea is a major cause of morbidity of rhesus macaques at the California National Primate Research Center (CNPRC) and in other captive macaque populations. 17,22,23,37,41 Infectious causes of diarrhea, particularly bacterial enterocolitis associated with Campylobacter infection, are common in the outdoor, group-housed population at the CNPRC. However, up to 25% of rhesus macaques develop an apparently noninfectious diarrhea syndrome referred to as idiopathic chronic diarrhea (ICD), the pathogenesis of which remains obscure. 4 This syndrome is a diagnosis of exclusion, based on failure to identify typical diarrheagenic agents and persistence or recurrence of clinical signs despite supportive therapy. Affected animals typically suffer a protracted course of diarrhea and repeated hospitalization before ultimately requiring euthanasia for the welfare of the animal. They are more frequently submitted to the CNPRC necropsy service during the winter and spring. These seasons comprise the rainy season, but it is not known if ICD is causally related to the weather, particularly as weaning also occurs in these seasons. There is a pressing need to understand the pathophysiology of ICD, determine how the disease may affect use of rhesus macaques in biomedical research, and develop biomarkers that identify at-risk animals earlier in the disease process.
Lymphoplasmacytic colitis is the most frequent microscopic finding in rhesus macaques with ICD. 4 This is a nonspecific lesion that is similar to microscopic colitis of humans and may also be seen in a subset of cases of diarrhea-predominant irritable bowel syndrome (D-IBS). 4,21,45 Campylobacter and other gastrointestinal pathogens of humans that are common to rhesus macaques may be associated with postinfectious syndromes including irritable bowel syndrome (IBS) and, perhaps, microscopic colitis. 28,34,35,42,45,49 The pathogenesis of these syndromes is poorly understood, but there may be some overlap, and both have been associated with ongoing, predominantly Th1-mediated immune activation and alterations in the density of intestinal enteroendocrine cells.* In particular, increased density of colonic serotonin-positive enterochromaffin cells (ECs), a subset of enteroendocrine cells, has been identified in both microscopic colitis and postinfectious IBS (PI-IBS). Altered serotonin signaling may result in hypermotility, hypersensitivity, and increased permeability of the gut as seen with PI-IBS. 14,15,32
In this study, we sought to more fully describe the history of infectious causes of diarrhea in animals that subsequently develop ICD and to investigate the presence of histologic and immunologic changes consistent with PI-IBS. Our results suggest that, similar to PI-IBS, ICD is associated with a history of Campylobacter infection, dysbiosis, and hyperplasia of enteroendocrine cells. However, unlike PI-IBS, cytokine expression by gut-resident CD4+ T cells was reduced in animals with ICD.
Materials and Methods
Animals
The CNPRC is an AAALAC (The Association for Assessment and Accreditation of Laboratory Animal Care)-accredited facility where animals are cared for in accordance with the USDA Animal Welfare Act and regulations and the Guide for the Care and Use of Laboratory Animals. 1,2,24 Established policies of the Institutional Animal Care and Use Committee (IACUC) of the University of California, Davis were followed. Animals were housed at the CNPRC either in outdoor corrals or indoors. All rhesus macaques that were submitted to necropsy over a period of 7 months with the primary problem of chronic diarrhea were investigated. Only rhesus macaques of Indian origin were assessed in this study. Animals were considered to have idiopathic chronic diarrhea based on the following criteria: greater than 45 days of recorded diarrhea and/or greater than 3 hospitalizations for treatment of diarrhea in the previous 6 months, negative culture and stool examination tests for bacterial and parasitic pathogens in the previous 6 months, no etiology for diarrhea identified at the time of necropsy by intestinal cultures and Giardia/Cryptosporidium immunofluorescent assay (IFA), and no etiology for diarrhea identified during microscopic analysis of tissues. Eleven rhesus macaques of Indian origin that were similarly housed and fed were assigned as controls. These rhesus macaques had no history of diarrhea in the previous year, were negative for bacterial and parasitic causes of diarrhea, and exhibited no gross or microscopic evidence of colitis. The control animals were submitted for necropsy for reasons unrelated to gastrointestinal disease or were control animals in other studies (Suppl. Table S1).
Necropsy
All animals underwent complete necropsies. Cultures of small intestinal contents, large intestinal contents, and bile for Campylobacter spp., Shigella flexneri, Yersinia species, and Escherichia spp. were carried out, as well as fecal examination by direct microscopy and IFA for Giardia and Cryptosporidium. The intestinal tract and any gross lesions identified at the time of necropsy were examined microscopically. Routine sections included duodenum (immediately distal to the stomach), mid-jejunum, terminal ileum, ileocecal junction, ileocecal lymph node, proximal colon, mid-colon, distal colon, and rectum. Tissues were fixed in 10% neutral buffered formalin for 3 days prior to being routinely processed, embedded in paraffin, sectioned at 5 μm, and stained with hematoxylin and eosin. Samples of proximal and distal colon were also fresh frozen in optimal cutting temperature (OCT) gel and stored at –80°C until use.
Semiquantitative Microscopic Assessment of Colitis, Epithelial Adherent Bacteria, and Trichomonad Burden
A single section of each segment of large intestine (cecum, proximal colon, mid-colon, and distal colon) was graded for the following features: lamina propria infiltration, acute inflammation, surface epithelium damage, surface epithelium tufting, crypt proliferation, goblet cell density, and intraepithelial lymphocytes (see Suppl. Table S2). Epithelial adherent bacteria were graded as follows: grade 1, no bacteria in the section; grade 2, bacteria present in patches on <50% of epithelial surface; and grade 3, bacteria covering the majority of the epithelial surface in the section. Trichomonad burden was graded as follows: grade 1, no trichomonads in the section; grade 2, small numbers of trichomonads in the lumen and superficial crypts; grade 3, moderate numbers of trichomonads in the lumen and multifocally extending deep into crypts; and grade 4, large numbers in the lumen and/or expanding into many crypts. The median of the score for each individual section of colon was taken as the individual animal score for each feature.
Immunohistochemistry
Sections (5 μm thickness) of small intestine (duodenum and jejunum) or large intestine (mid-colon and distal colon) were deparaffinized in xylene and hydrated through a series of ethanol and water washes. Antigen retrieval was achieved by heating to 100°C in Dako pH 6 antigen retrieval buffer (Agilent, Santa Clara, CA) for 30 minutes. Following blocking with 10% goat serum in Tris-buffered saline (TBS) for 1 hour, primary and secondary antibodies were applied (Suppl. Table S3). Detection was achieved using diaminobenzoate (DAB) chromogen (Agilent), and slides were counterstained with Mayer’s hematoxylin.
The density of epithelial CD3+ T cells (IEL) was assessed by counting the mean number of reactive cells per 100 enterocytes, over a total of 500 enterocytes and subsequently assigning a grade between 1 and 4 representing the quartiles of the results as follows: grade 1, <22 IELs/100 enterocytes; grade 2, 22 to 31 IELs/100 enterocytes; grade 3, 32 to 41 IELs/100 enterocytes; and grade 4, >41 IELs/100 enterocytes. The densities of enteroendocrine cells and enterochromaffin cells (chromogranin A and serotonin reactive, respectively) were assessed by counting the mean number of reactive cells per 600× field and subsequently assigning a grade between 1 and 4 representing the quartiles of the results. For chromogranin A: grade 1, <10/600× field; grade 2, 10 to <13/600× field; grade 3, 13 to <15/600× field; and grade 4, >15/600× field. For serotonin: grade 1, <8/600× field; grade 2, 8 to <10/600× field; grade 3, 10 to 12/600× field; and grade 4, >12/600× field. Counts were converted to categorical data because accurate counts cannot be achieved in hematoxylin and eosin sections without using a stereological approach.
Laser Capture Microscopy
Cryostat sections (10 μm thickness) of OCT-frozen proximal colon from 3 control animals were mounted on PEN-Membrane slides (Leica). Dissection of the epithelial layer and adherent bacteria was carried out using a Leica LMD6000 microscope and Leica Laser Microdissection V 6.5 software. Tissue segments were stored in RX buffer (Qiagen, Valencia, CA) on dry ice and transferred to –20°C prior to DNA extraction.
Extraction of Protozoal and Bacterial DNA and Sequencing
DNA was extracted from 10-μm sections of formalin-fixed, paraffin-embedded (FFPE) tissues by the method of Coura et al 11 and Gillio-Tos et al. 19 Conventional polymerase chain reaction (PCR) was performed using primers TRF1 (5′-TGCTTCAGTTCAGCGGGTCTTCC) and TRF2 (5′- CGGTAGGTGAACCTGCCGTTGG) that are specific to the 5.8S ribosomal RNA (rRNA) gene segment of trichomonads, by the method of Hayes et al. 20 DNA was extracted from laser-captured cells using a Qiagen DNeasy kit per the manufacturer’s instructions (Qiagen). Conventional PCR was performed using the bacteria-wide 16S rRNA primers E334F (5′-CCAGACTCCTACGGGAGGCAGC) and E939R (5′- CTTGTGCGGGCCCCCGTCAATTC) by the method of Baker et al. 6 A blank-reagent negative control was included in all DNA extractions and PCR reactions. All amplicon products from FFPE and laser capture cells were submitted to the University of California Davis UCDNA Sequencing facility (http://dnaseq.ucdavis.edu/) for bidirectional sequencing. Sequences were analyzed with Sequencher 5.4.1.
Isolation of Peripheral Blood Mononuclear Cells and Lamina Propria Mononuclear Cells and Flow Cytometry Analysis
Peripheral blood mononuclear cells (PBMCs) and lamina propria mononuclear cells (LPMCs) were isolated from a subset of 10 animals with ICD and 8 controls. Heparinized whole blood was collected at necropsy, and PBMCs were extracted using lymphocyte separation medium (MP Biomedicals, Burlingame, CA) using standard techniques. LPMCs were extracted from 2-cm-long loops of terminal ileum and mid-colon. Sections of gut were immediately rinsed in sterile phosphate-buffered saline (PBS), minced with scissors, and shaken with 50 ml of cell release medium (AIM-V medium, Life Technologies, Carlsbad, CA; supplemented with 5% v/v heat-inactivated fetal bovine serum, 6.25 mM HEPES buffer, 0.1 mg/ml DNAse, 0.5 mg/ml collagenase, penicillin, and streptomycin) for 2 hours at 37°C. Solids were removed by straining followed by centrifugation at 1800 rpm, and LPMCs were isolated from the supernatant using a Percoll gradient. PBMCs and LPMCs were suspended in 5% DMSO in heat-inactivated fetal bovine serum and stored in liquid nitrogen in aliquots of 5 million cells until use.
Flow Cytometry
The following antibodies from BD Biosciences (San Jose, CA) were used: anti-CD3 BV786 (SP34-2), anti-CD4 BV650 (L200), anti-CD8 PerCPCy5.5 (SK1), anti-CD3 Alexa 700 (SP34-2), anti-CD20 FITC, anti–interleukin (IL) 2 APC (MQ1-17H12), anti–tumor necrosis factor α (TNFα) Alexa 700 (Mab11), anti–interferon γ (IFNγ) PE-Cy7 (B27), and anti–IL-4 BV421 (8D4-8). In addition, the following antibodies from other manufacturers were used: anti–IL-17A AF488 (Bio64DEC17; ebioscience, San Diego, CA), anti-FoxP3 Pacific Blue (206D; Biolegend, San Diego, CA), and anti–IL-13 PE (JES10-5A2.2; Miltenyi, Auburn, CA). For the T-regulatory cell assay, frozen cells were thawed and resuspended into V-bottom plates in a brilliant violet staining buffer (BD Biosciences) with extracellular antibodies for 20 minutes at 4°C in the dark. Cells were washed with PBS containing 2% fetal bovine serum (FBS), then permeabilized with FOXP3 Fix/perm buffer (Biolegend) according to the manufacturer’s instructions. These cells were then suspended in brilliant violet staining buffer with anti-FoxP3 and anti-CD3, then incubated in the dark at room temperature for 30 minutes. Cells were then washed in PBS with 2% FBS and resuspended in 0.5% paraformaldehyde (PFA) in PBS/FBS and held overnight at 4°C.
For the cytokine production assay, cells were initially stimulated for 16 hours with 50 ng/ml phorbol myristate acetate and 1 μg/ml ionomycin in the presence of 5 μg/ml GolgiPlug (BD Biosciences). Cells were then stained in PBS containing 2% FBS with anti-CD4 and anti-CD8 (clones and colors given above). Intracellular staining employed Cytofix/Cytoperm from BD Biosciences following the manufacturer’s instructions. Cells were resuspended in 50 μl brilliant violet staining buffer containing anti-CD3, anti–IL-2, anti-TNFα, anti-IFNγ, and anti–IL-4, then incubated for 30 minutes in the dark at 4°C. Cells were washed in BD Perm/Wash Buffer, washed again in PBS with 2% FBS and 0.5% PFA, and held overnight at 4°C. Flow cytometry data were collected using a BD Fortessa.
Statistical Analysis
Comparison of histologic features between animals with ICD and controls was carried out in Prism 6 (GraphPad Software, San Diego, CA) using nonparametric statistics. Flow cytometry data were gated and analyzed with FlowJo, and then comparisons between animals with ICD and controls were carried out in the R programming language.
Results
ICD Is a Disease of Outdoor-Housed, Juvenile Rhesus Macaques
During the 7 months of this study, a total of 95 animals were submitted to necropsy because of intractable diarrhea, which is consistent with the frequency of such cases in the previous 5 years. Of the 95 animals submitted to necropsy due to intractable diarrhea, 35 met the criteria necessary to diagnose ICD. Sixty percent of these animals were females, but when normalized for sex distribution in the colony, the sexes were equally represented (Fig. 1). Most animals affected by ICD were outdoor-housed and juveniles under the age of 5 years at the time of necropsy (Figs. 2, 3). The 60 animals submitted to necropsy but not meeting the criteria for ICD were similar in age and history and were excluded primarily due to having recorded diarrhea of insufficient duration or, rarely, being pathogen positive at the time of necropsy (13/60).
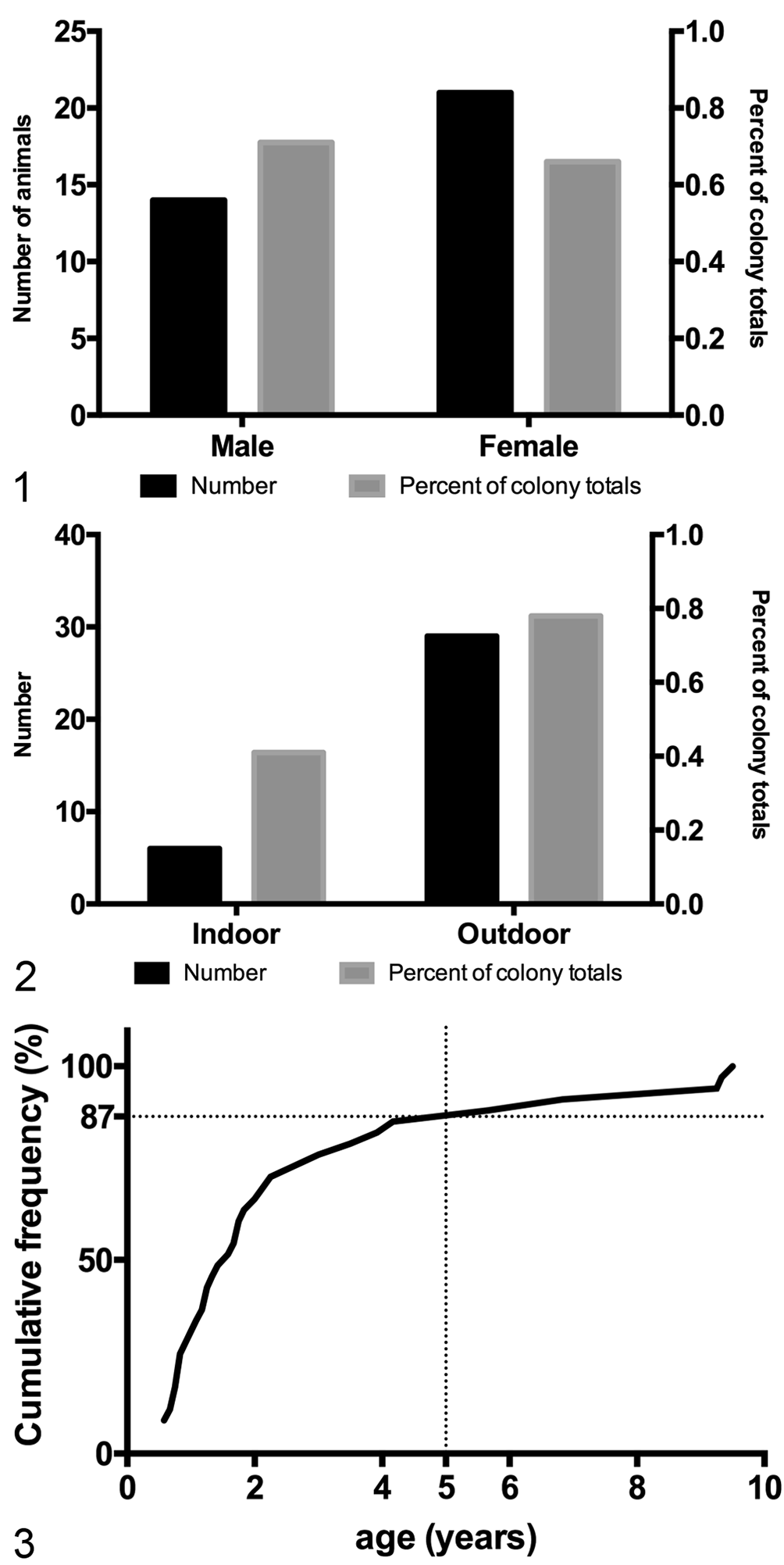
Demographics of idiopathic chronic diarrhea (ICD). Figure 1. Counts and frequencies of animals with ICD, by sex. Counts are shown with black bars measured against the left vertical axis. Frequencies are shown with gray bars measured against the right vertical axis (percentage of total colony population). Both sexes were equally represented among the total colony population (gray bars). Colony census for year of study: 1965 males and 3204 females. Figure 2. Counts and frequencies of animals with ICD, by housing location. Affected animals were more likely to be outdoor-housed (gray bars). Colony census for year of study: 1467 indoor-housed and 3702 outdoor-housed. Figure 3. A plot of cumulative frequency shows that 87% of all cases occur before 5 years of age (dotted lines).
Juvenile rhesus macaques are rarely submitted to necropsy for reasons other than gastrointestinal disease. As a result, assigning age-matched control animals was challenging, and the median age of controls was greater than that of those with ICD (Suppl. Table S1).
Historic Campylobacter and Trichomonad Infections Are Common in Animals With ICD
Rectal cultures and direct fecal examinations were carried out during each episode of diarrhea suffered by animals with ICD that were assigned to this study (median number of episodes: 4; range, 1–9). Isolation of Campylobacter spp. during 1 or more of these episodes was far more frequent than isolation of other bacterial species (Fig. 4). Of note, bacteria and parasites that are considered primary pathogens of the gastrointestinal tract (ie, Campylobacter jejuni, Yersinia spp., Shigella flexneri, Giardia spp., Cryptosporidium spp., and Trichuris trichiura) were infrequently isolated from animals that progressed to ICD. In contrast, bacteria and parasites that are typically considered commensal, such as Campylobacter coli, Campylobacter lari, and trichomonads, were frequently isolated in high numbers from animals that progressed to ICD. A direct comparison to the control cohort is impossible as fecal cultures and parasite exams are only regularly taken from animals with diarrhea. However, at the time of necropsy, only 9% (1/11) of control animals had Campylobacter (in low numbers) and 0% were positive for trichomonads by fecal examination, in comparison to 40% Campylobacter spp. (14/35) and 26% trichomonads (9/35) in ICD animals (Fig. 5). Of 35 animals with ICD, 16 received antibiotics (doxycycline, metronidazole, and/or tylosin) within a month prior to necropsy. Positive bacterial cultures and the identification of trichomonads were less frequent in these 16 animals (Fig. 6); thus, the prevalence of Campylobacter spp. and trichomonad infections in ICD cases may be underestimated.
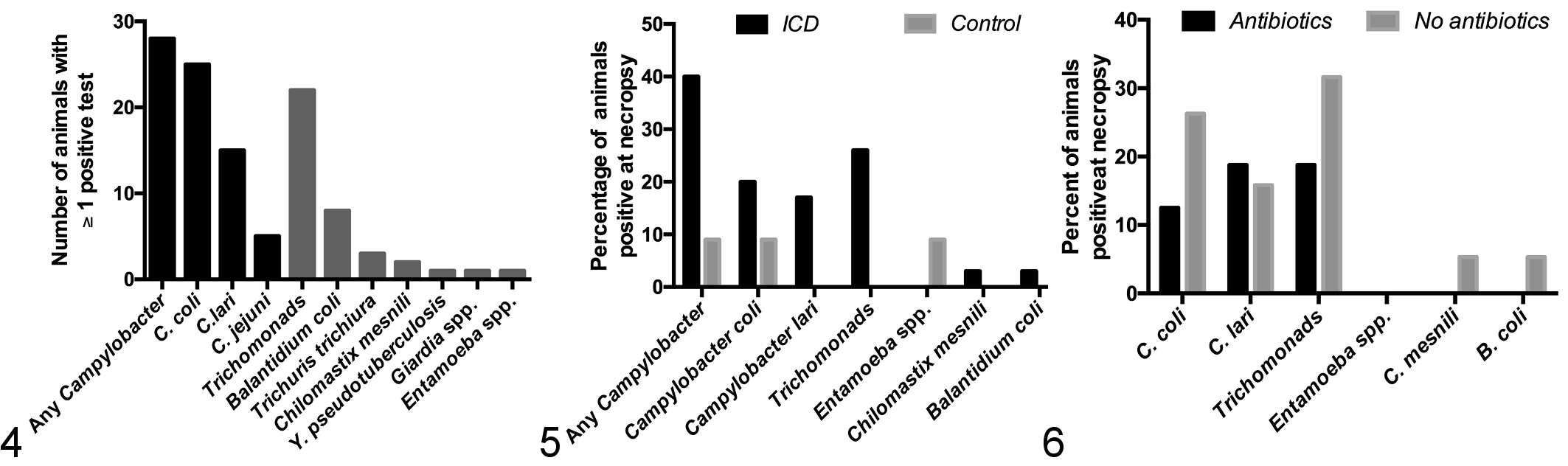
Animals with idiopathic chronic diarrhea (ICD) have a history of Campylobacter infections. Figure 4. Twenty-eight of 35 animals with ICD had at least 1 historic bout of diarrhea that was culture positive for Campylobacter spp., predominantly Campylobacter coli (black bars). Identification of trichomonads on fecal examination during bouts of diarrhea was also common (gray bars). Figure 5. Campylobacter spp. and trichomonads were more likely to be identified in animals with ICD than in controls. Figure 6. Antibiotic treatment within a month prior to necropsy of animals with ICD was associated with a slightly reduced incidence of Campylobacter spp. and trichomonad infections (black bars compared to gray).
Of the 13 animals with diarrhea that were excluded due to pathogen detection at necropsy, the most common isolate was C. jejuni (7/13). Two animals were culture positive for S. flexneri, 2 for T. trichiura, and a single animal for Giardia spp. One animal with no history of diarrhea could not be assigned as a control because it cultured positive for S. flexneri at the time of necropsy.
ICD Colitis Is Associated With Loss of Normal Colonic Epithelial Colonization With Helicobacter macacae and Overgrowth of Trichomonads
Macroscopic findings in animals with ICD were consistent with previous reports and consisted of a distended colon containing watery, nonbloody fecal material. The colonic mucosa was mildly thickened and nonulcerated, and local lymph nodes were enlarged. Microscopic, predominantly lymphoplasmacytic, inflammation was present at all levels of the large intestine examined (Fig. 7). Histologic evidence of acute inflammation was variably present in the form of crypt abscesses and/or mucus plugs with cellular debris and mucosal neutrophil infiltration (Fig. 8). In addition, mucosal hypertrophy, loss of goblet cells, increased epithelial lymphocytes, and superficial attenuation and/or tufting of mucosal epithelium were present to varying degrees in different individuals (Figs. 9, 10). Paneth cell metaplasia in the colon was rarely present, and fibrosis was not a feature of ICD in the examined animals.
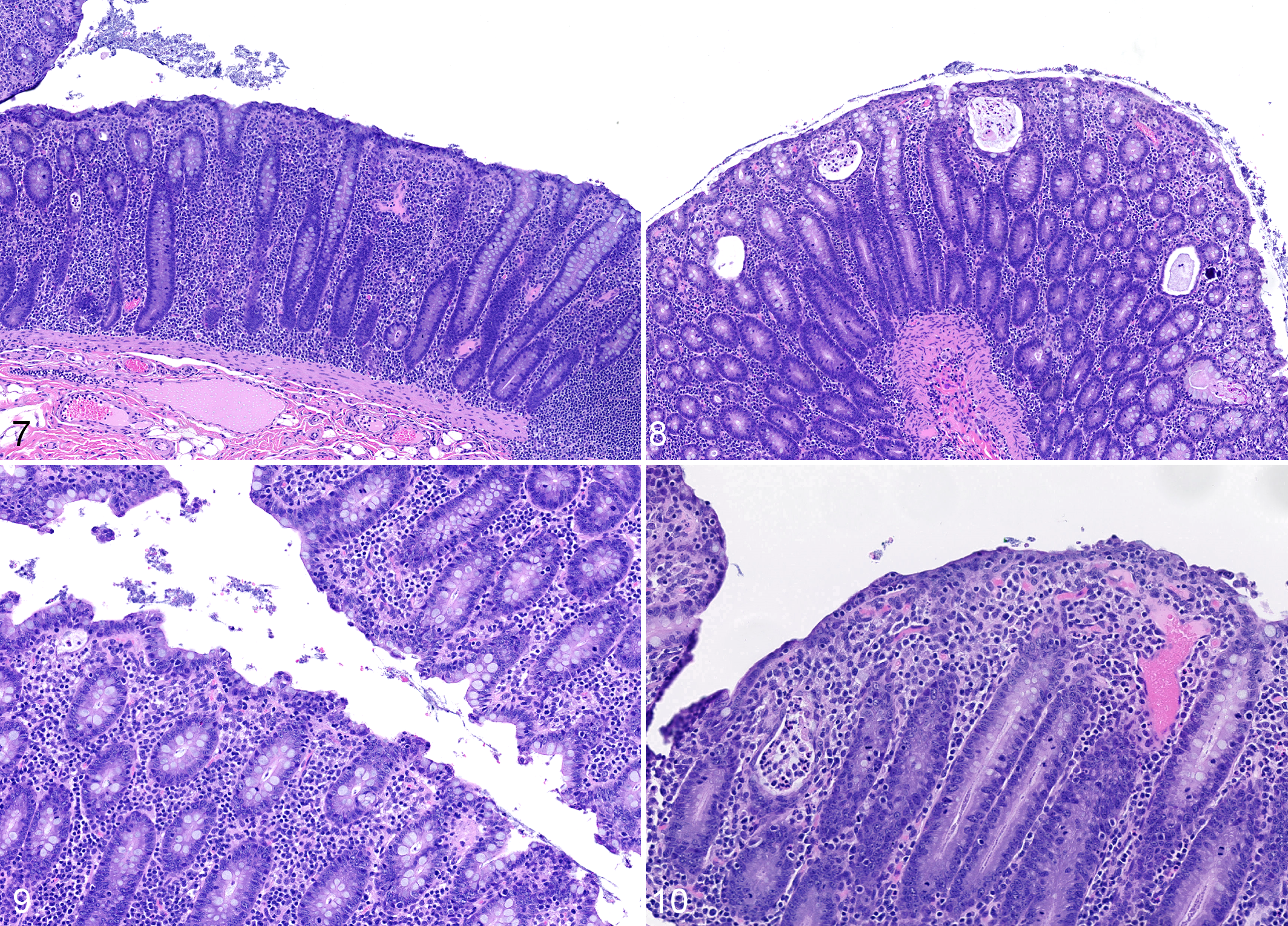
Idiopathic chronic diarrhea (ICD), colon, rhesus macaque. Figure 7. The lamina propria is expanded by an infiltrate of lymphocytes and plasma cells that separate and raise crypts. Crypts are elongated. Hematoxylin and eosin (HE). Figure 8. Crypts are expanded and filled with cellular debris, mucus, and/or neutrophils. HE. Figure 9. Superficial epithelium is multifocally tufted, interpreted as increased turnover and sloughing of superficial epithelial cells. There are increased numbers of intraepithelial lymphocytes in the superficial mucosa. HE. Figure 10. Superficial mucosa is multifocally attenuated with low cuboidal to squamous epithelial cells, and there are reduced numbers of goblet cells. There are superficial infiltrates of lymphocytes, plasma cells, and macrophages.
We noted a conspicuous absence in the ICD cohort of normal colonic epithelium-adherent bacteria, termed the blue brush border (Figs. 11, 12). We also observed greater abundance of microscopically evident trichomonad-like parasites (Figs. 13, 14), which frequently extended into colonic crypts. In contrast to the fecal examination results presented in Fig. 5, microscopically increased burdens of trichomonads were associated with antibiotic therapy in the month prior to necropsy (Fig. 15). Complete grading data for all histopathology and immunohistochemistry end points are available in Supplemental Table S4.
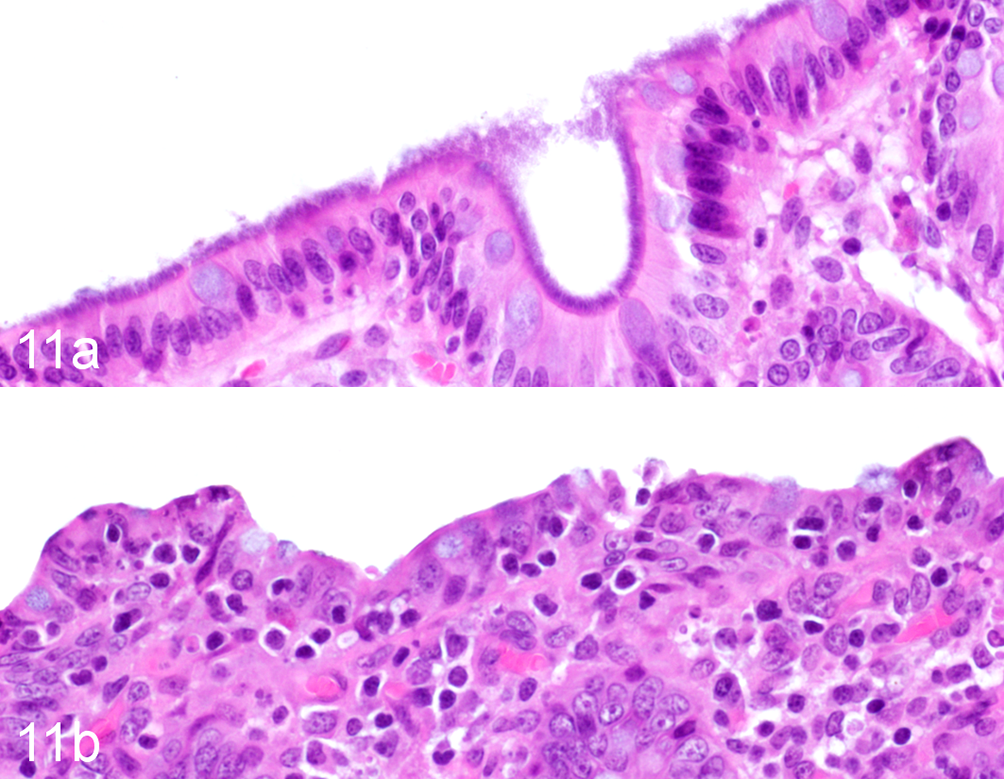
Idiopathic chronic diarrhea (ICD) is associated with decreased epithelial adherent bacteria. (a) Colon, rhesus macaque, control animal. The superficial mucosa of the normal rhesus macaque colon is densely colonized by bacteria that were identified as a pure population of Helicobacter macacae. Hematoxylin and eosin (HE). (b) ICD, colon, rhesus macaque. Bacteria are largely absent on the superficial mucosa. HE.
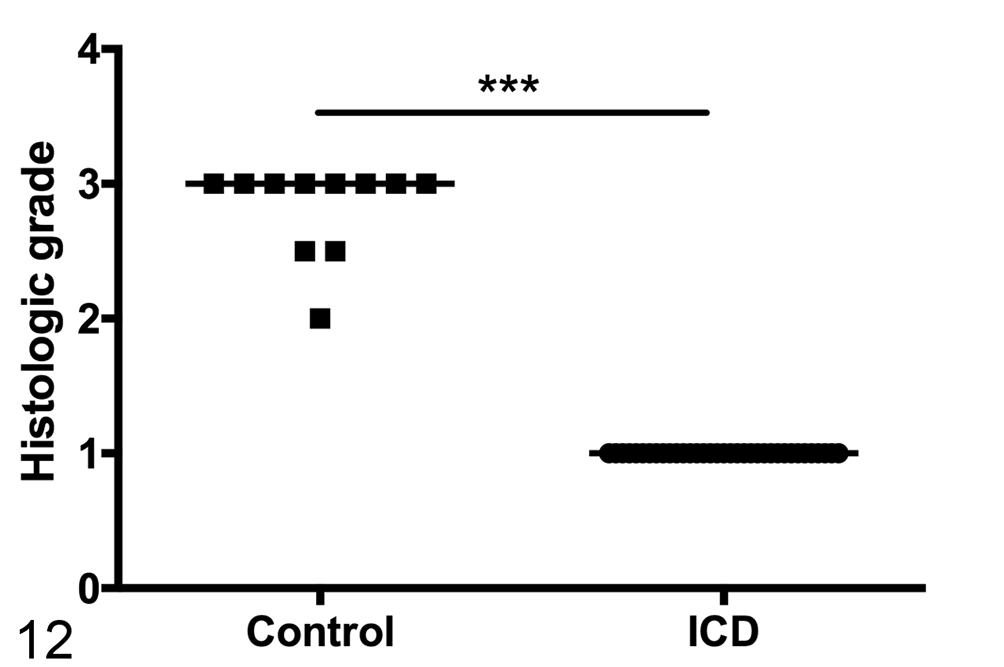
Semiquantitative assessment of superficial epithelial adherent bacteria shows a significant decrease in animals with ICD. Line represents median score. Mann-Whitney test, ***P < .0001.
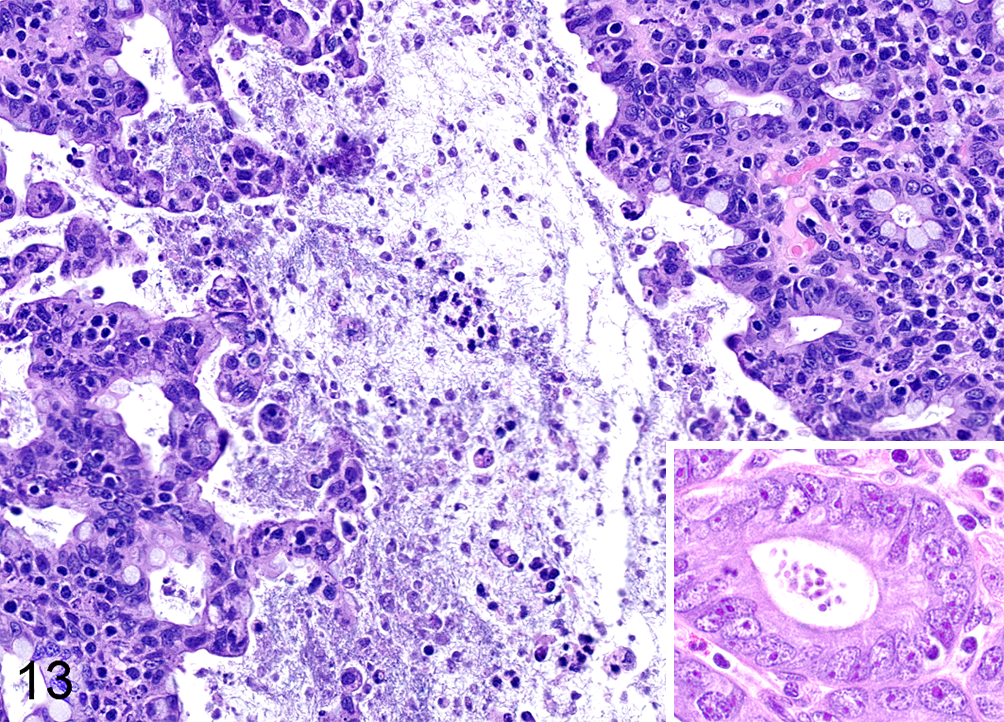
Idiopathic chronic diarrhea (ICD) is associated with an increased burden of trichomonads. Numerous trichomonad parasites are present within the lumen of the colon and extending into the superficial crypts. Note the marked tufting of the superficial mucosa. Inset: Multifocally protozoa extend deep into and distend crypts. Hematoxylin and eosin.
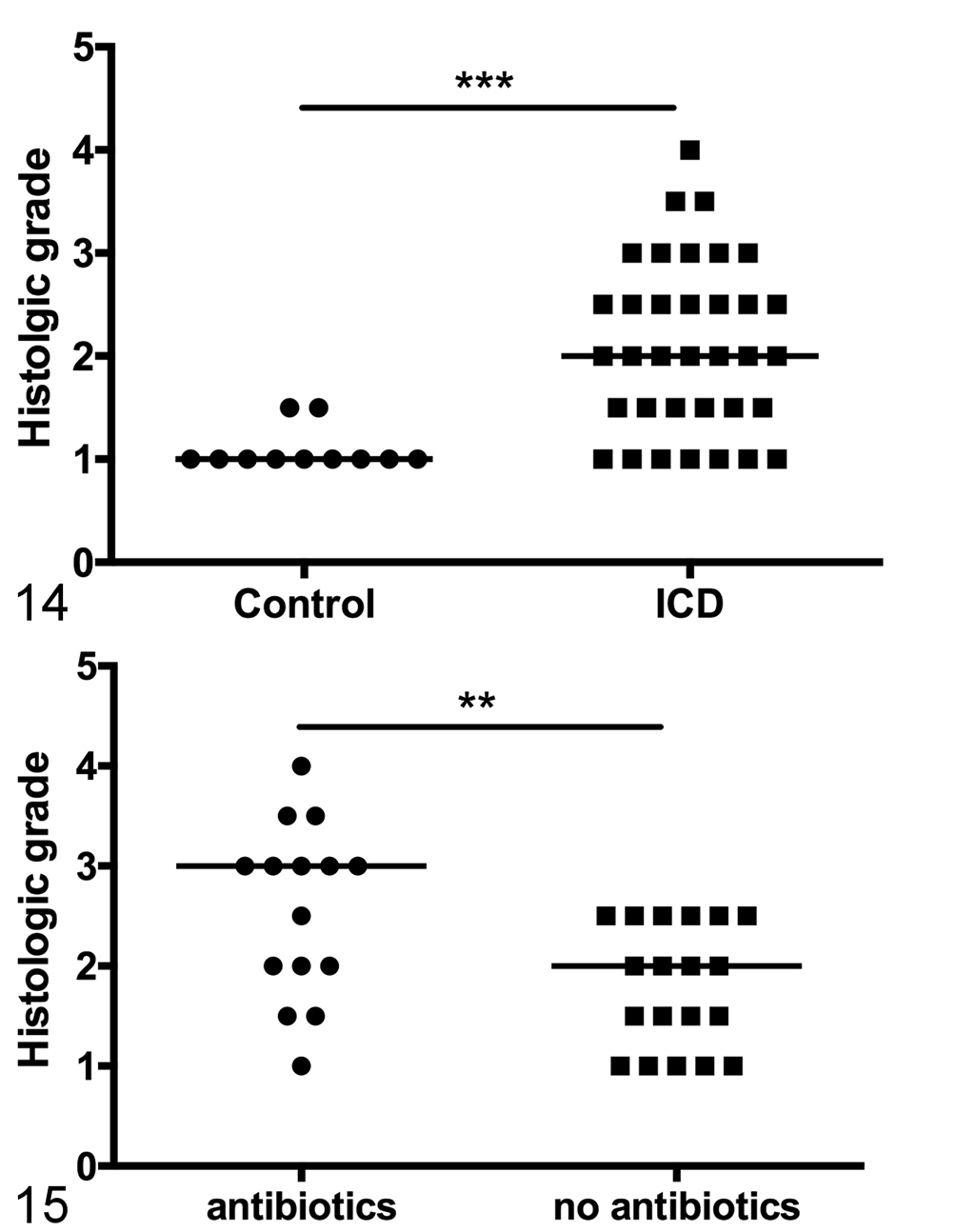
Idiopathic chronic diarrhea (ICD) is associated with an increased burden of trichomonads. Figure 14. Semiquantitative assessment of trichomonad burden shows a significant increase in animals with ICD. Line represents median score. Mann-Whitney test, ***P < .0001. Figure 15. Among ICD animals, antibiotics in the month prior to necropsy are associated with increased presence of trichomonads. Line represents median score. Mann-Whitney test, **P < .0073.
PCR-based sequencing of the 16S rRNA gene of colonic epithelium-adherent bacteria, via laser capture microscopy of the colonic epithelial layer of 3 control animals, revealed the bacteria (866 bp; GenBank KT203770) to be a pure population 100% identical to Helicobacter macacae (GenBank KC894696). Analysis of the trichomonad-like parasite population showed that the trichomonads (338 bp; GenBank KT203771) were a mixed population composed primarily of parasites 98% identical to Pentatrichomonas hominis (Genbank AF156964).
ICD Is Associated With Greater Frequency of Enterochromaffin Cells in the Colonic Epithelium and T-Cells in the Small Intestine
Given the prevalence of Campylobacter infections both within the colony and in the ICD cohort, we sought to assess the frequency of enteroendocrine cells in the inflamed colon. We identified a significant increase in serotonin-reactive enterochromaffin cells in the mid-colon of animals with ICD compared to controls (Figs. 16–18). Similarly, the number of chromogranin A–reactive enteroendocrine cells was increased in ICD animals (Fig. 19). There was a greater frequency of both enteroendocrine and enterochromaffin cells in the distal colon/rectum than the mid-colon, but there were no differences between ICD and control animals in this region.
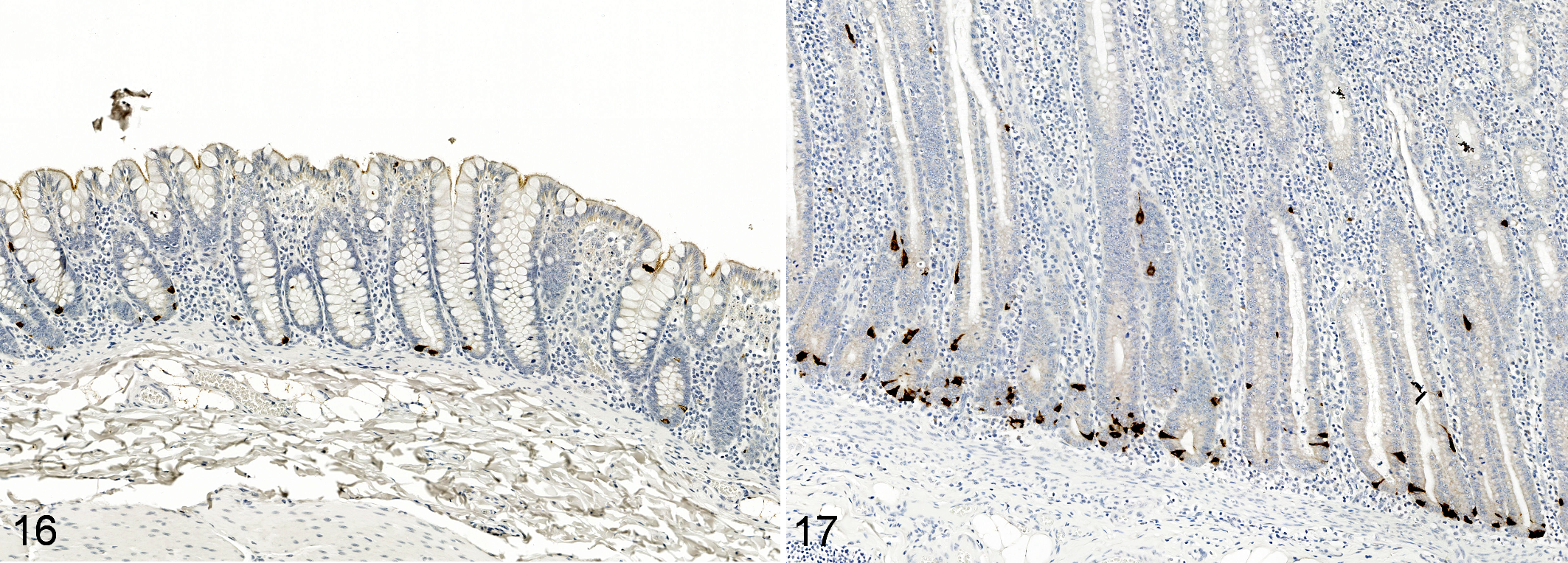
Idiopathic chronic diarrhea (ICD) is associated with increased numbers of enterochromaffin cells in the mid-colon. Immunohistochemistry for serotonin. Figure 16. Colon, rhesus macaque, control animal. Enterochromaffin cells are sparse in the uninflamed colon with typically no more than 2 per crypt. Figure 17. ICD, mid-colon, rhesus macaque. There are numerous enterochromaffin cells per crypt.
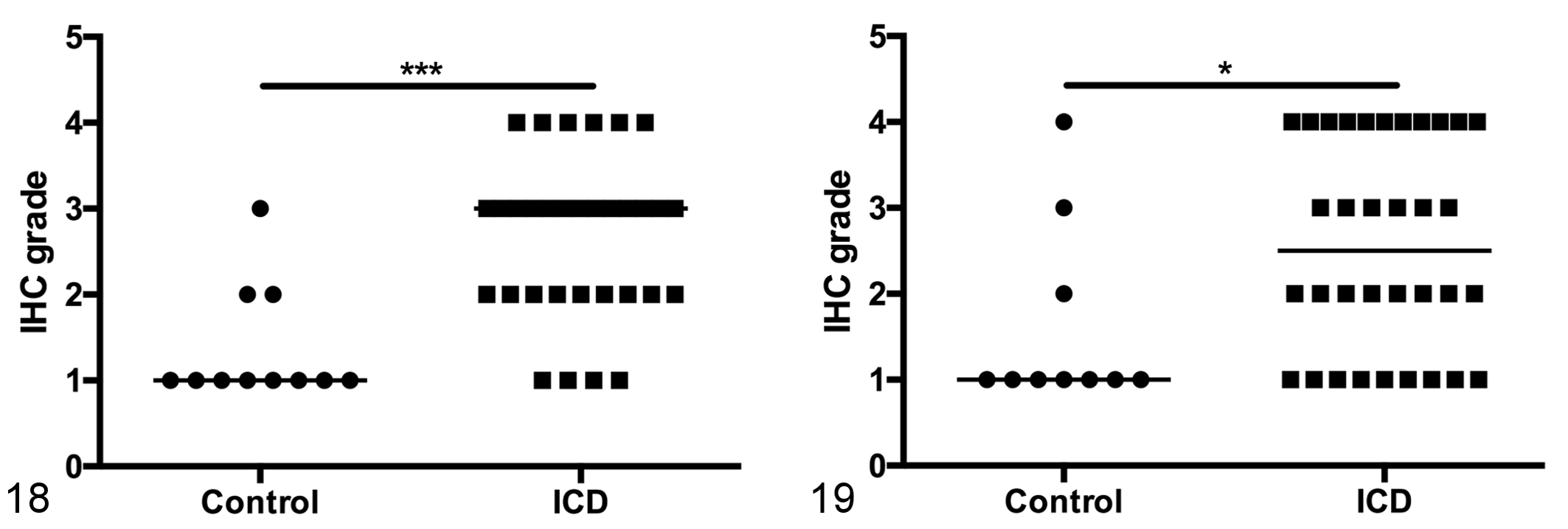
Idiopathic chronic diarrhea (ICD) is associated with increased numbers of enterochromaffin cells in the mid-colon. Figure 18. Semiquantitative assessment of enterochromaffin cells (serotonin-reactive) in the mid-colon shows a significant increase in animals with ICD. Line represents median score. Mann-Whitney test, ***P = .0002. Figure 19. Semiquantitative assessment of enteroendocrine cells (chromogranin A-reactive) in the mid-colon shows a significant increase in animals with ICD. Line represents median score. Mann-Whitney test, *P = .0233. IHC, immunohistochemistry.
Small intestinal disease has been noted in humans with IBS and those previously infected with Campylobacter, 39,46,52 but previous descriptions of ICD-associated gastrointestinal pathology have been restricted to the lymphoplasmacytic colitis described above. 4,8 In our cohort of animals with ICD, we noticed an increased frequency of CD3+ intraepithelial lymphocytes in the small intestine (jejunum) in addition to the previously described colitis (Figs. 20–22). Similar increases in epithelial lymphocytes may have been present in the duodenum, but preservation of duodenal mucosal epithelium was often poor, hampering the assessment of this finding. Mild lymphoplasmacytic inflammation of the distal ileum, adjacent to the ileocecal junction, was variably present in animals with ICD.
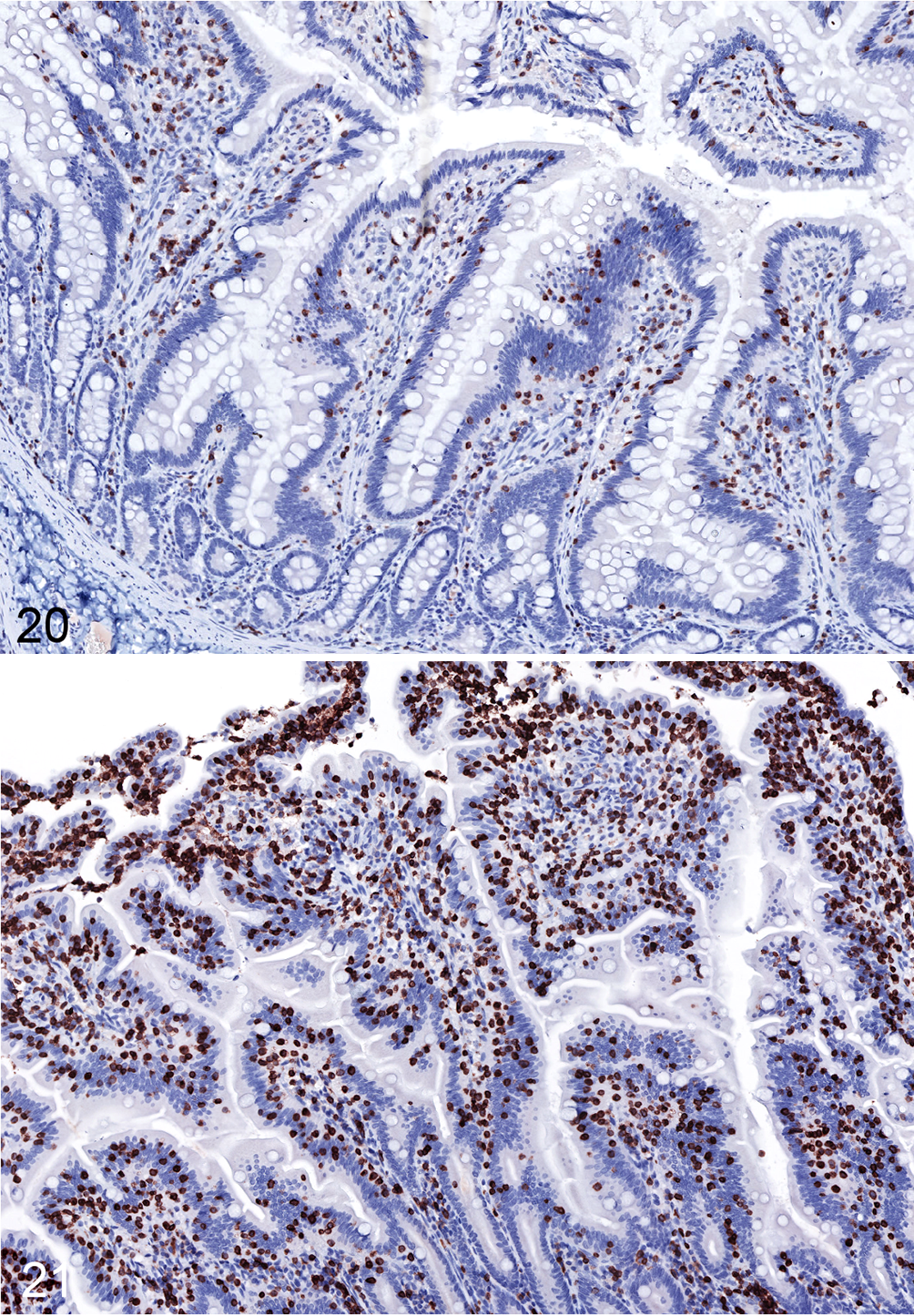
Idiopathic chronic diarrhea (ICD) is associated with increased numbers of intraepithelial T cells in the jejunum. Figure 20. Jejunum, rhesus macaque, control animal. Low numbers of intraepithelial T cells are present. Immunohistochemistry for CD3. Figure 21. ICD, jejunum, rhesus macaque. There are many intraepithelial lymphocytes that frequently form clusters. Immunohistochemistry for CD3.
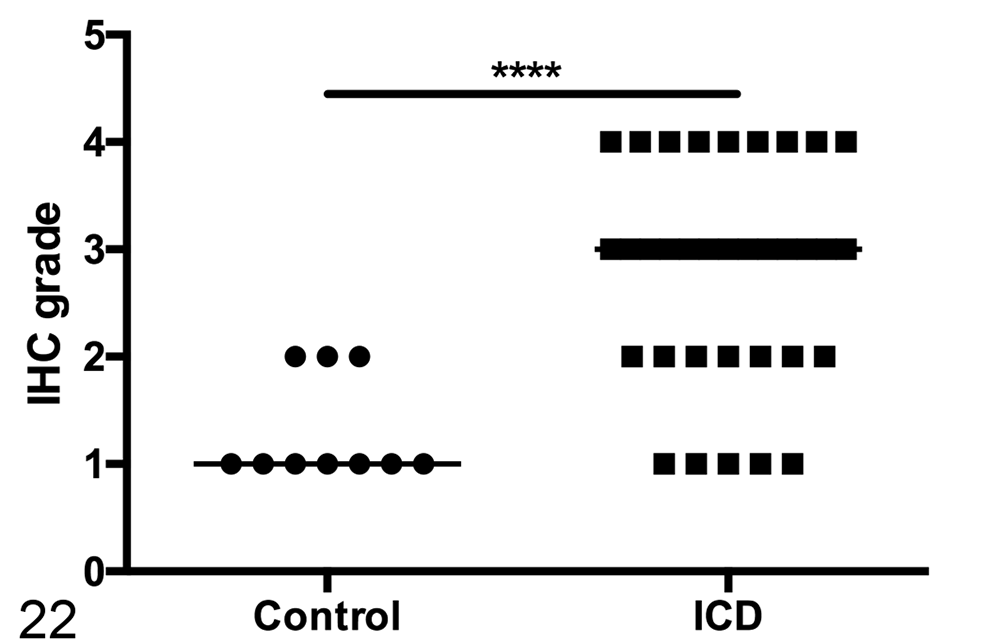
Semiquantitative assessment of intraepithelial T cells in the jejunum shows a significant increase in animals with idiopathic chronic diarrhea (ICD). Line represents median score. Mann-Whitney test, ****P < .0001. IHC, immunohistochemistry.
ICD Is Associated With a Decrease in Colonic Th2 Cells
To determine if the microscopically evident inflammation and apparent altered microbiota of ICD animals (prevalence of trichomonads and absence of Helicobacter along the epithelial surfaces) were associated with altered immune function, we tested the capacity for production of various cytokines by CD3+ T cells from animals in a subset of the cohort. Isolated PBMCs and ileal LPMCs from the 2 groups showed similar frequency of cells expressing the T-cell proinflammatory cytokines, IFN-gamma, TNF-alpha, and IL-17, though there was unexpectedly lesser frequency among colonic LPMCs from ICD animals (Figs. 23 and 24 in ileum and colon, respectively). Expression of the Th2-associated cytokine IL-4, however, was virtually undetectable among ileal and colonic CD4+ T cells of the ICD cohort (Figs. 25–28). IL-13 expression was also less frequent among ileal but not colonic cells from ICD animals (Figs. 25, 26). This effect was exclusive to CD4+ T-helper cells; capacity for cytokine production by CD3+CD8+ T cells was similar in the ICD and control cohorts (data not shown).
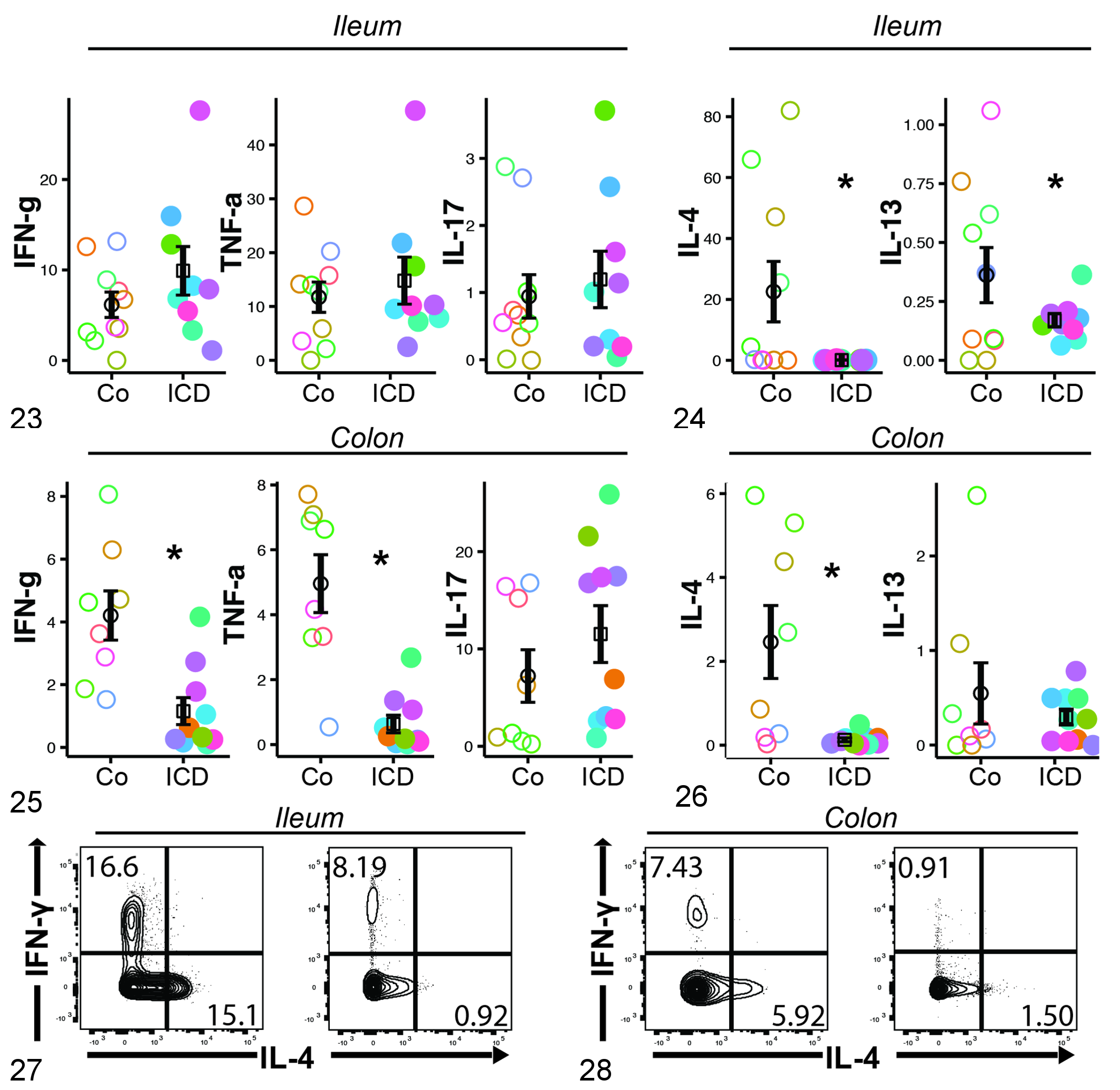
Idiopathic chronic diarrhea (ICD) animals have decreased representation of interleukin 4 (IL-4)–expressing and IL-13–expressing CD4+ T cells in the ileum and colon. Data shown represent cytokine production among CD4+ lamina propria lymphocytes from the ileum (Figs. 23, 25, and 27) or colon (Figs. 24, 26, and 28) of rhesus macaques with ICD vs control animals. Figures 23 and 24. Expression of proinflammatory cytokines in the ileum and colon. Figures 25 and 26. Expression of Th2 cytokines by T cells in the ileum and colon. Figures 27 and 28. Representative flow cytograms demonstrating greater potential for IL-4 and interferon γ (IFNγ) expression among ileal (Fig. 27) and colonic (Fig. 28) T cells of healthy controls (left) than animals with ICD (right). Colors represent individual animals. Student t test, *P < .05.
The decrease in potential for expression of proinflammatory cytokines suggested a possible expansion of regulatory T cells; however, there was no significant difference in CD4+FoxP3+ populations between ICD animals and healthy controls (Suppl. Fig. S1). Complete data for flow cytometry end points are available in Supplemental Tables S5 (colon) and S6 (ileum).
Discussion
ICD is prevalent in captive rhesus macaque populations that serve as vital immunologic models of disease, and yet studies investigating the condition’s immunopathology are scarce. In this study, we sought to more fully describe the demographics of animals affected with ICD, evaluate differences seen in peripheral and gut T-cell biology, and, given the prevalence of Campylobacter infections at the CNPRC, investigate histologic similarities between ICD and postinfectious syndromes of humans.
The lesions of ICD have previously been reported to be restricted to the proximal colon, 4 but in our cohort, the entire large intestine was affected, and more frequent lymphocytes in the small intestine were observed. Animals in the present study had severe diarrhea requiring euthanasia, unlike the stable animals described by Ardeshir et al, 4 which may in part explain the more extensive lesions.
Campylobacter infections are becoming more common in the United States and may be associated with chronic gastrointestinal sequelae. 39 The most frequent sequela is diarrhea-predominant IBS, which may affect up to 36% of those with acute campylobacteriosis, although the reported frequency is extremely variable. 28,39,40,43 The pathophysiology of PI-IBS is not well understood, in part due to a lack of appropriate animal models, but is thought to involve continued, low-grade mucosal immune activation, alterations in EC density and signaling, and increased gut permeability. 14,39,44,46 A history of Campylobacter infections was common in our cohort of animals with ICD and is in fact common throughout the colony: an ongoing prospective study suggests approximately 50% prevalence of subclinical Campylobacter infections in healthy, outdoor-housed rhesus macaques. Our data show that animals with ICD also have an increased density of enterochromaffin cells in the colon, as well as intraepithelial lymphocytes in the small intestine, similar to findings in PI-IBS and potentially as a result of Campylobacter infection. Serotonin is released following mechanical or chemical stimulation of enterochromaffin cells and signals through receptors on multiple cells, including afferent nerve fibers and enterocytes, to induce motility, secretion of bicarbonate and chloride, and perception of pain. 32 In addition, serotonin may contribute to mucosal inflammation through serotonin receptors on many different leukocytes. 5 While increased density of enterochromaffin cells is not specific to PI-IBS, these effects are thought to result in many of the symptoms; for this reason, serotonin receptor antagonists were developed for PI-IBS treatment. 38 While long-term treatment of rhesus macaques with serotonin antagonists is not practical, investigation of these therapeutics in affected animals would help to clarify the role of serotonin signaling in this disease.
Alterations in the intestinal bacterial microbiota have been associated with many chronic gastrointestinal diseases, including ICD, PI-IBS, and microscopic colitis of humans. 18,26,33 In our cohort of animals with ICD, 40% were culture positive for Campylobacter spp. at the time of necropsy despite multiple rounds of treatment. Furthermore, nonbiased assessment of the colonic bacterial community in rhesus macaques with colitis suggested that the Campylobacteraceae were overrepresented compared to healthy animals even when culture negative. 33 Abundant colon-adherent spirochaetes, likely H. macacae identified herein, have previously been noted in healthy rhesus macaques but may have been miscategorized as Brachyspira spp. 12,33 The lack of epithelial adherent H. macacae in animals with ICD may be an indication of the overall dysbiosis associated with ICD or previous antibiotic exposure or may be secondary to the increased turnover of surface epithelial cells associated with mucosal hyperplasia. 8,33 However, given the close apposition of these bacteria to colonic epithelial cells, it is possible they exert an important influence on the mucosal immune system, similar to the role of segmented filamentous bacteria in the development of Th17 cells in mice, 25 and lack of these bacteria may be directly related to ICD disease in affected animals. Colonization of the colon by Helicobacter spp. thus far appears to be a rhesus macaque–specific phenomenon. Alterations in the frequency of either spirochaetes or Campylobacter spp. have not been associated with PI-IBS or microscopic colitis in humans. 18,26,33
Alterations of the commensal eukaryotic community of the colon associated with gastrointestinal disease have not been extensively investigated. We observed an increased frequency of trichomonad parasites in animals with ICD, predominantly Pentatrichomonas hominis. P. hominis is generally accepted to be nonpathogenic in macaques and humans. 30,50 However, there are sparse reports of trichomonads resulting in gastrointestinal disease in immune-suppressed rhesus macaques and in other species. 9,51,54 While increased trichomonads may simply be indicative of the dysbiosis associated with ICD, further investigation of P. hominis as a potential pathogen is warranted. Our results provide conflicting evidence of the effect of antibiotics on trichomonad burden: by fecal examinations, antibiotics appeared to lower the burden, while by microscopy, the burden was increased. These findings are likely explained by the fact that trichomonads were often closely associated with the mucosa, extending into glands, and therefore were less likely to be identified in feces.
Our assessment of T-cell cytokines in ICD animals yielded surprising results. Underrepresentation of IL-4–expressing CD4+ T cells in the ileum and colon of rhesus macaques affected by ICD suggests either (1) failure to develop these cells, perhaps due to a preexisting abnormality of the gut microbiota, or (2) disease-induced repression or shift away from the classical Th2 phenotype. Simultaneous repression of the capacity for IFNγ production (Fig. 25) suggests a disease-induced process that results in generally lesser development or maintenance of differentiated effector T cells. This finding is reminiscent of the decreased cytokine production noted in bottle-fed infant rhesus macaques compared to breastfed infant rhesus macaques, especially because bottle feeding is a known risk factor for ICD development. 3 Previous studies that assessed T-cell function in rhesus macaques with ICD demonstrated both increased activation and a shift toward Th1 cytokine production. 8,41 Although neither was identified within our study, the decrease of cells producing Th2 cytokines does not conflict with those results. Broadhurst and colleagues 8 found that an increased level of Th2 cytokines, including IL-4, following exposure of ICD animals to T. trichiura larvae was associated with improvement of clinical signs; however, it was not clear if the underlying problem was attenuated Th2 signaling or elevated Th1 cytokines. Results in humans are similarly nebulous; studies assessing cytokine concentrations in the mucosa of humans with PI-IBS or microscopic colitis are few, and the results in IBS are conflicting. 7 Most studies support increased concentrations of Th1 cytokines. 7,10 Some studies of chronic colitis in children have shown decreased production of IL-4, and IL-4 gene therapy has proven effective in treating induced colitis in a murine model. 27,53 A 2015 study also suggested that patients with PI-IBS have subnormal production of IL-13. 48 Whether the altered function of mucosal+ CD4 T cells is a primary deficit in ICD or is secondary to either alterations in the microbiota or prolonged inflammation will require further investigation.
In conclusion, ICD of rhesus macaques is associated with a history of Campylobacter infection, colonic dysbiosis characterized by loss of H. macacae colonization and increased numbers of trichomonads, increased small intestinal epithelial lymphocytes, and increased colonic enterochromaffin cells. Despite the clearly inflammatory nature of ICD colitis, the frequency of mucosal CD4+ T cells expressing cytokines was, in general, decreased, particularly Th2 cytokines. These data suggest that several organisms, C. coli, P. hominis, and H. macacae, should be investigated further for their pathogenic or protective role in ICD and contribution to alteration of the mucosal immune system. Furthermore, while ICD is clearly an imperfect model for PI-IBS, enterochromaffin dysfunction is a feature of both diseases.
Supplemental Material
Supplemental Material, DS1-3_VET_10.1177_0300985818780449 - Idiopathic Colitis in Rhesus Macaques Is Associated With Dysbiosis, Abundant Enterochromaffin Cells and Altered T-Cell Cytokine Expression
Supplemental Material, DS1-3_VET_10.1177_0300985818780449 for Idiopathic Colitis in Rhesus Macaques Is Associated With Dysbiosis, Abundant Enterochromaffin Cells and Altered T-Cell Cytokine Expression by Steven T. Laing, David Merriam, Barbara C. Shock, Sarah Mills, Abbie Spinner, Rachel Reader, and Dennis J. Hartigan-O’Connor in Veterinary Pathology
Supplemental Material
Supplemental Material, DS4_VET_10.1177_0300985818780449 - Idiopathic Colitis in Rhesus Macaques Is Associated With Dysbiosis, Abundant Enterochromaffin Cells and Altered T-Cell Cytokine Expression
Supplemental Material, DS4_VET_10.1177_0300985818780449 for Idiopathic Colitis in Rhesus Macaques Is Associated With Dysbiosis, Abundant Enterochromaffin Cells and Altered T-Cell Cytokine Expression by Steven T. Laing, David Merriam, Barbara C. Shock, Sarah Mills, Abbie Spinner, Rachel Reader, and Dennis J. Hartigan-O’Connor in Veterinary Pathology
Supplemental Material
Supplemental Material, DS5_VET_10.1177_0300985818780449 - Idiopathic Colitis in Rhesus Macaques Is Associated With Dysbiosis, Abundant Enterochromaffin Cells and Altered T-Cell Cytokine Expression
Supplemental Material, DS5_VET_10.1177_0300985818780449 for Idiopathic Colitis in Rhesus Macaques Is Associated With Dysbiosis, Abundant Enterochromaffin Cells and Altered T-Cell Cytokine Expression by Steven T. Laing, David Merriam, Barbara C. Shock, Sarah Mills, Abbie Spinner, Rachel Reader, and Dennis J. Hartigan-O’Connor in Veterinary Pathology
Supplemental Material
Supplemental Material, DS6_VET_10.1177_0300985818780449 - Idiopathic Colitis in Rhesus Macaques Is Associated With Dysbiosis, Abundant Enterochromaffin Cells and Altered T-Cell Cytokine Expression
Supplemental Material, DS6_VET_10.1177_0300985818780449 for Idiopathic Colitis in Rhesus Macaques Is Associated With Dysbiosis, Abundant Enterochromaffin Cells and Altered T-Cell Cytokine Expression by Steven T. Laing, David Merriam, Barbara C. Shock, Sarah Mills, Abbie Spinner, Rachel Reader, and Dennis J. Hartigan-O’Connor in Veterinary Pathology
Footnotes
Acknowledgments
Laser capture microscopy was conducted using the CAMI core facility at the UC Davis Center for Health and the Environment.
Our thanks are extended to Anne Gibbons, Mark Allen, and Sona Santos, who carried out many of the necropsies for the animals investigated in this study.
Immune function studies were carried out in the Flow Cytometry Core at the California National Primate Research Center.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Office of the Director of the National Institutes of Health under grant P51 OD011107.
Supplementary material for this article is available online.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.