
Abstract
The epithelial-mesenchymal transition (EMT) is a dynamic process linked to metastasis in many tumor types, including mammary tumors. In this study, we evaluated E-cadherin and vimentin immunolocalization in primary canine mammary carcinomas (20 cases) and their respective metastases, as well as their relationship with the core regulators SNAIL/SLUG. To assess the number of cells undergoing the process of EMT, we quantitated double-positive (E-cadherin+/vimentin+) cells using immunofluorescence, via cell counting and image analysis. In addition, SNAIL/SLUG expression was evaluated by established immunohistochemical methods. Primary tumors had significantly more E-cadherin+/vimentin+ co-expression than their paired respective lymph node or distant metastasis, respectively. Furthermore, the percentage of E-cadherin+/vimentin+ cells in grade II and III carcinomas was significantly higher than in grade I tumors. Primary tumors had significantly higher SNAIL/SLUG expression when analyzed based on the percentage of positive cells compared with their respective distant metastases in pairwise comparisons. An inverse correlation was noted between SNAIL/SLUG immunoreactivity and percentage of E-cadherin+/vimentin+ immunopositive cells in primary tumor samples when SNAIL/SLUG immunoreactivity was grouped into 2 categories (high versus low) based on percentage-positive staining. These results show a positive correlation between E-cadherin+/vimentin+ cells and higher tumor grade, establish differences between primary tumor and their respective metastases, and provide further support that EMT plays a critical role in the metastasis of canine mammary carcinoma. Furthermore, these data suggest that modulation of this process could provide greater therapeutic control and provide support for further research to determine if E-cadherin+/vimentin+ co-immunoreactivity imparts predictive value in the clinical outcome of patients with canine mammary carcinomas.
Keywords
Mammary tumors are the most common neoplasm affecting sexually intact female dogs and thus represent a serious health problem in dogs all over the world.* The etiology of mammary tumors is multifactorial and includes genetic, environmental, nutritional, and hormonal factors. 1,33,50,51,54,57,59,61 Fifty percent of these tumors are malignant, with carcinomas of various subtypes being the most prevalent. 18,43,60 Highly variable clinical outcomes occur in dogs with mammary carcinomas and are influenced by such factors as tumor histological type, grade, and clinical stage. Dogs with stage I (<3 cm) and grade I primary tumors typically have a favorable long-term survival when treated with surgery alone. 61 However, metastasis remains a frequent cause of cancer-related death in dogs diagnosed with advanced-stage disease or higher-grade tumors. 9,20,31,32,61 Therefore, similar to women with breast cancer, predicting, preventing, and treating metastasis represents the most important obstacle to improve outcomes in dogs with clinically aggressive neoplastic subtypes. To do so requires an improved understanding of the complex biological and molecular changes that occur in the malignant epithelial cells during the transition to a metastatic phenotype. Despite progress, the process of metastasis, and specifically the interactions between tumor cells and their microenvironment that drives and/or facilitates metastasis, is incompletely understood. 31
The epithelial-mesenchymal transition (EMT) has been identified as a major player in tumor metastasis and progression. EMT is a dynamic process that occurs during epithelial tumor development and invasion, which has been associated with aggressive clinical behavior. 36 During EMT, epithelial cells undergo multiple biochemical changes to acquire a mesenchymal-like phenotype characterized by enhanced migratory capacity, invasiveness, and increased production of extracellular matrix components. In addition, during this process, epithelial cells may develop stem cell characteristics, such as increased resistance to apoptosis, anoikis, and immunosuppression. 13,24,27,28
EMT has been associated with the metastatic process in many different types of epithelial malignancies, including mammary carcinomas. 3,13,64,67 Accumulating evidence also supports a phenotypic plasticity of tumor cells that allows a reverse process, specifically the mesenchymal-epithelial transition (MET), to occur when metastatic cells colonize a distant secondary site. 38 MET is more challenging to document in patients with spontaneously occurring tumors, however, because of limitations in access to both primary tumors and respective metastatic lesions.
Nevertheless, certain features associated with EMT have been well documented, in both research models and naturally-occurring mammary cancer in humans and dogs, and support a crucial role for EMT, and possibly MET, in tumor progression and metastasis. Specifically, the down-regulation of epithelial cell-to-cell adhesion proteins, such as E-cadherin, and the up-regulation of mesenchymal proteins such as vimentin have been associated with aggressive breast cancer behavior and poor outcomes. † Growth factors, such as transforming growth factor–β and epidermal growth factor, induce EMT through induction of WNT and NOTCH signaling pathways and activation of downstream transcription factors such as SMAD, SNAIL, ZEB, and TWIST, although this remains controversial. 13,28,35,63
Research to further elucidate how EMT is regulated and the mechanisms by which it promotes aggressive cancer behaviors will improve our insight into the biology of metastasis and may inform the development of new drugs or treatment strategies. 13 Based on previous research on EMT in canine mammary carcinomas and its association with tumor progression and metastasis, we hypothesize that a subset of tumor cells within canine malignant mammary carcinomas co-express both E-cadherin and vimentin (E-cadherin+/vimentin+) during their transition from an E-cadherin–dominant expression (epithelial) to a vimentin-expressing phenotype (mesenchymal) and that there would be a decline in numbers of these E-cadherin+/vimentin+ cells in paired metastatic lesions of these tumors (due to the reversing process of MET). The primary objective of this research project was to test this hypothesis by studying the process of EMT, specifically E-cadherin and vimentin expression, and its core regulators (SNAIL/SLUG) in primary tumors and paired metastatic lesions of dogs with mammary carcinomas.
Materials and Methods
Case Selection and Histopathological Analysis
Twenty canine primary mammary tumors from 20 different dogs with grade I, II, or III carcinoma and biopsy-confirmed metastasis (lymph node or distant metastasis) were randomly selected from archived samples from cases in which tissues from the primary tumors as well as metastatic lymph node and/or distant metastasis were available. All samples were obtained from patients enrolled in prospective clinical studies performed at the Penn Vet Shelter Canine Mammary Tumor Program, School of Veterinary Medicine, and the Penn Vet Cancer Center of the University of Pennsylvania (N = 7) and Faculty of Veterinary Medicine and Biosciences, Norwegian University of Life Sciences (N = 13). Both studies were approved by their respective Institutional Animal Care and Use Committees. All primary tumors and metastases were routinely processed through 10% formalin fixation and paraffin embedded (FFPE), as previously described. 17,48 For this study, freshly cut histopathologic sections were stained with hematoxylin and eosin (HE). All previously diagnosed canine mammary carcinoma biopsies were independently assessed by 2 board-certified veterinary pathologists (A.C.D., R.L.A.) and classified and graded according the 2011 and 2012 system by Goldschmidt and Peña 18,47 (Table 1). Four primary tumors were grade I, 6 primary tumors were grade II, and 10 primary tumors were grade III. Sections from regional lymph nodes (n = 10; collected from the superficial inguinal [n = 5] or axillary [n = 5] lymph nodes) and/or distant metastasis (n = 16; collected from the lung [n = 12], liver [n = 1], or sternal [n = 2] or bronchial [n = 1] lymph nodes) corresponding to individual primary tumors were evaluated to ensure the histopathological features were consistent with metastatic mammary carcinomas.
Histological Types of 20 Canine Primary Mammary Tumors.

E-Cadherin and Vimentin Immunofluorescence Analysis
The FFPE tissues were evaluated for E-cadherin and vimentin expression by immunofluorescence. Sequential (4-μm-thick) sections from the blocks of all primary tumors, lymph node metastases, and distant metastases described above were prepared on charged slides for immunofluorescence staining. Sections were deparaffinized by immersion in Clear-Rite 3 (Thermo Fisher Scientific, Waltham, MA) for 10 minutes at room temperature (RT), followed by immersion in solutions of decreasing ethanol concentration (100%, 95%, and 70%), for 3 minutes each. The slides were then washed in deionized water, and antigen retrieval was performed by boiling in citrate buffer (DAKO; Dako Denmark A/S, Glostrup, UK), according to the established laboratory procedures. 4 Following a protein block performed in a 5% bovine serum albumin (BSA), 5% goat serum, and 0.05% Tween-20 (Bio-Rad, Hercules, CA) in phosphate-buffered saline (PBS) solution for 30 minutes at RT, samples were incubated with the primary antibodies. Samples were then rinsed 3 times for 5 minutes in PBS prior to incubation with the secondary antibodies for 1 hour at RT. Primary and secondary antibodies used are described in Table 2. For negative controls, no primary antibody was added. Finally, samples were washed 3 times in PBS for 10 minutes and then mounted with DAPI (Thermo Fisher Scientific).
Antibodies and Protocols Used for Immunohistochemistry.
Abbreviation: RT, room temperature.
Immunofluorescence analysis
Five images were taken for each slide per sample, at high-power (40× objective) and 20× objective fields using an Olympus Microscope (BX51). The fields chosen were based on areas with malignant epithelial cells confirmed by HE. To assess the potential transition through the EMT process, the number of tumor cells positive and negative for E-cadherin or vimentin (E-cadherin+
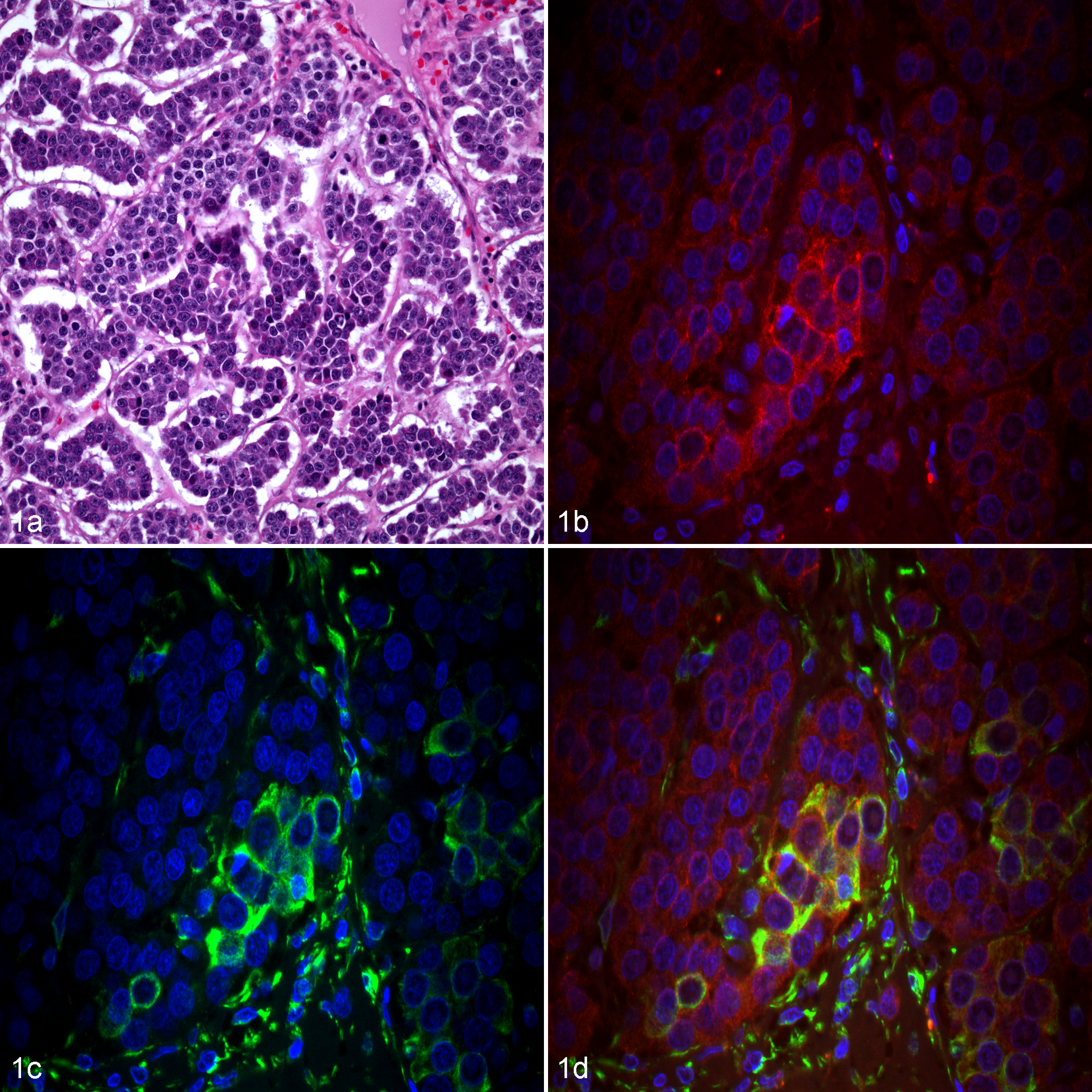
Mammary carcinoma, female dog. (a) Grade III canine mammary carcinoma. Hematoxylin and eosin. (b-d) Immunofluorescence. Nuclear staining with DAPI (blue). (b) Neoplastic epithelial cells exhibit positive membranous and cytoplasmic immunoreactivity for E-cadherin (red). (c) Epithelial cells with positive cytoplasmic immunolabeling for vimentin (green). (d) Co-localization of E-cadherin and vimentin in the cytoplasm of neoplastic epithelial cells (yellow).
Results were expressed as the median value (with ranges) of the percentage of E-cadherin and vimentin-positive/-negative cells. In addition, the area of E-cadherin+/vimentin+ immunoreactivity, viewed with a 20× objective lens, was determined per section using ImageJ software (National Institutes of Health, Bethesda, MD; http://rsb.info.nih.gov/ij/; Tables 3 and 4).
Percentage of Cells With Immunolabeling for E-Cadherin, Vimentin, and SNAIL/SLUG+ in Primary Tumors (PT), Lymph Node Metastasis (LM), and Distant Metastasis (DM) of Canine Mammary Carcinoma.a
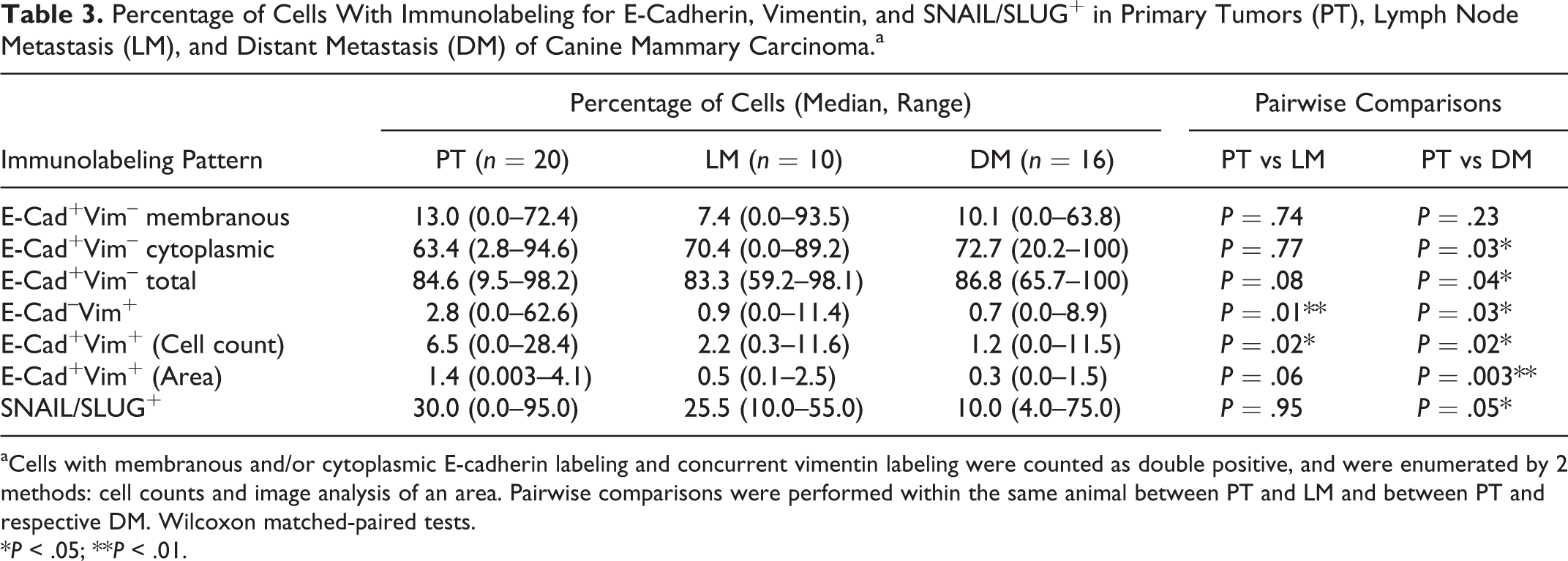
aCells with membranous and/or cytoplasmic E-cadherin labeling and concurrent vimentin labeling were counted as double positive, and were enumerated by 2 methods: cell counts and image analysis of an area. Pairwise comparisons were performed within the same animal between PT and LM and between PT and respective DM. Wilcoxon matched-paired tests.
*P < .05; **P < .01.
Percentage of Cells With Immunolabeling for E-Cadherin (E-cad), Vimentin (Vim), and SNAIL/SLUG+ in Canine Mammary Carcinomas of Grades I, II, and III.a
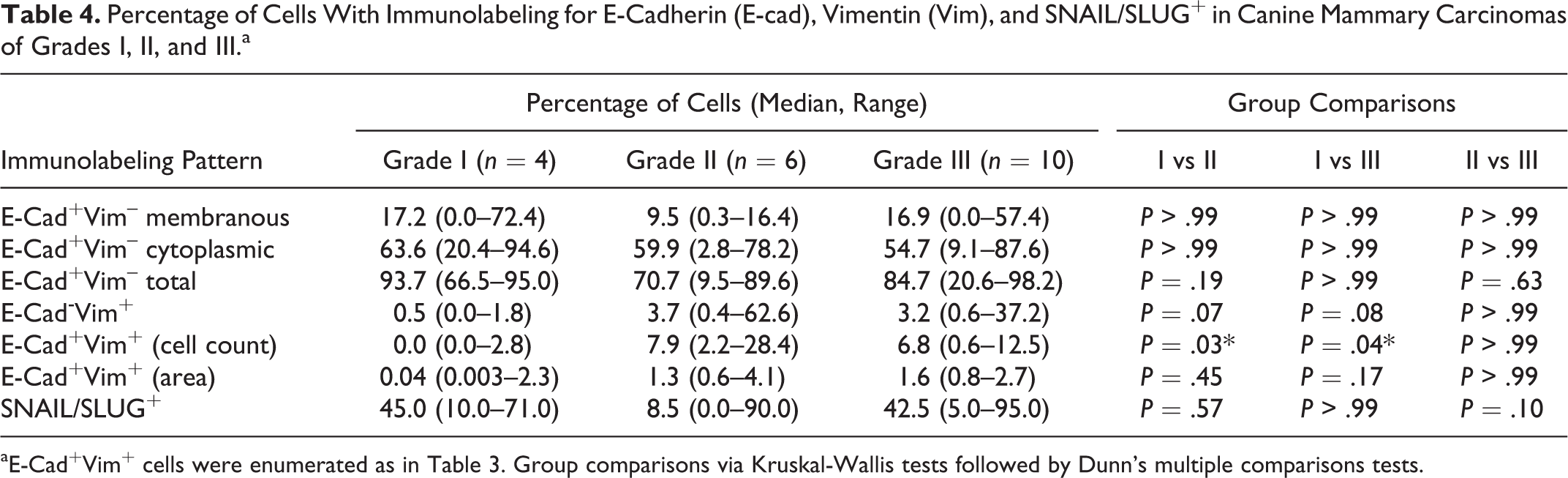
aE-Cad+Vim+ cells were enumerated as in Table 3. Group comparisons via Kruskal-Wallis tests followed by Dunn’s multiple comparisons tests.
SNAIL/SLUG Immunohistochemical Analysis
SNAIL/SLUG protein expression was evaluated in FFPE tissues by immunohistochemistry. Sections of each tumor were prepared on charged slides, deparaffinized, and rehydrated. Antigen retrieval was performed by citrate buffer boiling, followed by blocking in 3% H2O2, PBS containing 1% BSA (A5611 Sigma-Aldrich, St Louis, MO) and 10% goat serum (protein blocking solution), avidin blocking solution, and biotin blocking solution (Avidin Blocking Kit; Vector Laboratories Inc, Burlingame, CA). 4 Primary antibody and IgG control (negative control; Table 2) were incubated for 1 hour at RT in protein blocking solution. Secondary antibody (Table 2) was applied and incubated in 1% of BSA/PBS for 1 hour at RT. Sections were incubated in an avidin–biotin–peroxidase system (ABC Elite; Vector Laboratories Inc) and in 3,3′-diaminobenzidine, DAB Substrate (Dako, Carpinteria, CA) for 1 minute 30 seconds at RT. Slides were counterstained in hematoxylin, before dehydration and mounting. A canine fibrosarcoma sample served as a positive control. 25 The same specific areas that were evaluated for E-cadherin/vimentin co-expression were evaluated for SNAIL/SLUG staining and limited to regions with malignant epithelial cells. This was confirmed by comparing the slides with parallel sections stained with HE.
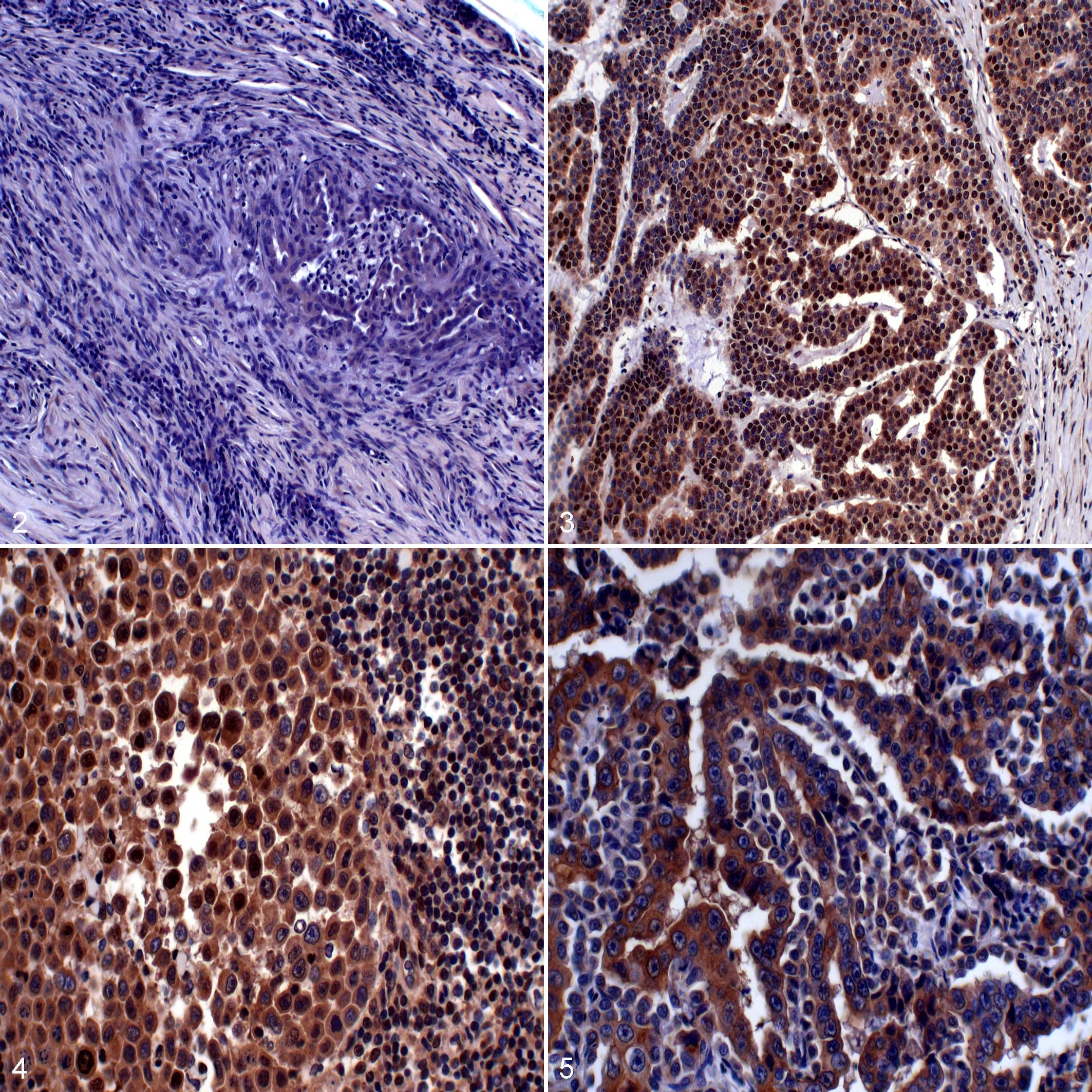
SNAIL/SLUG immunopositivity was counted as percentage positive cells and scored according to the criteria described by Gamba et al., 17 where 0 represents <5%, +1 equals 5% to 50%, and +2 equals >50% of immunopositive neoplastic cells of the total neoplastic cell population. The tumors were further grouped as SNAIL/SLUGlow (scores 0 and +1) and SNAIL/SLUGhigh (score +2; Figs. 2–5). Raw data are provided as Supplemental Table S1.
Data Analysis
Statistical analyses were performed using GraphPad Prism v.7.0 (GraphPad Software Inc, La Jolla, CA). P < .05 was considered significant. A Kruskal-Wallis test was applied to compare the expression of E-cadherin and/or vimentin between grades (I, II, or III), followed by a Dunn’s multiple comparison test. Wilcoxon matched-paired tests were used for pairwise comparisons between the expression of E-cadherin and/or vimentin in the individual primary tumor and its respective metastasis (lymph node or distant metastasis). Mann-Whitney U-tests were used to compare E-cadherin+/ vimentin+ cells in SNAIL/SLUGhigh versus SNAIL/SLUGlow primary tumors. The chi-square exact test was used to determine the association between SNAIL/SLUG score and tumor location. Linear regression (Spearman correlation) was used to assess the association between the percentage of SNAIL/SLUG–positive cells and E-cadherin+/vimentin+.
Results
E-Cadherin and Vimentin Expression by Immunofluorescence
The analysis of E-cadherin or vimentin is shown in Table 3. E-cadherin staining was recorded as membranous cytoplasmic and total (membranous + cytoplasmic). Overall, there were low percentages of cells with membranous E-cadherin+ labeling in primary tumors, lymph node metastases, and distant metastases regardless of primary tumor grade (median values of 13%, 7.4%, and 10.1%; Tables 3 and 4). Considering the pairwise comparison of E-cadherin and/or vimentin between the respective primary tumor, lymph node metastasis, and distant metastasis, there was a significant increase in percentage of cytoplasmic or total E-cadherin+/vimentin
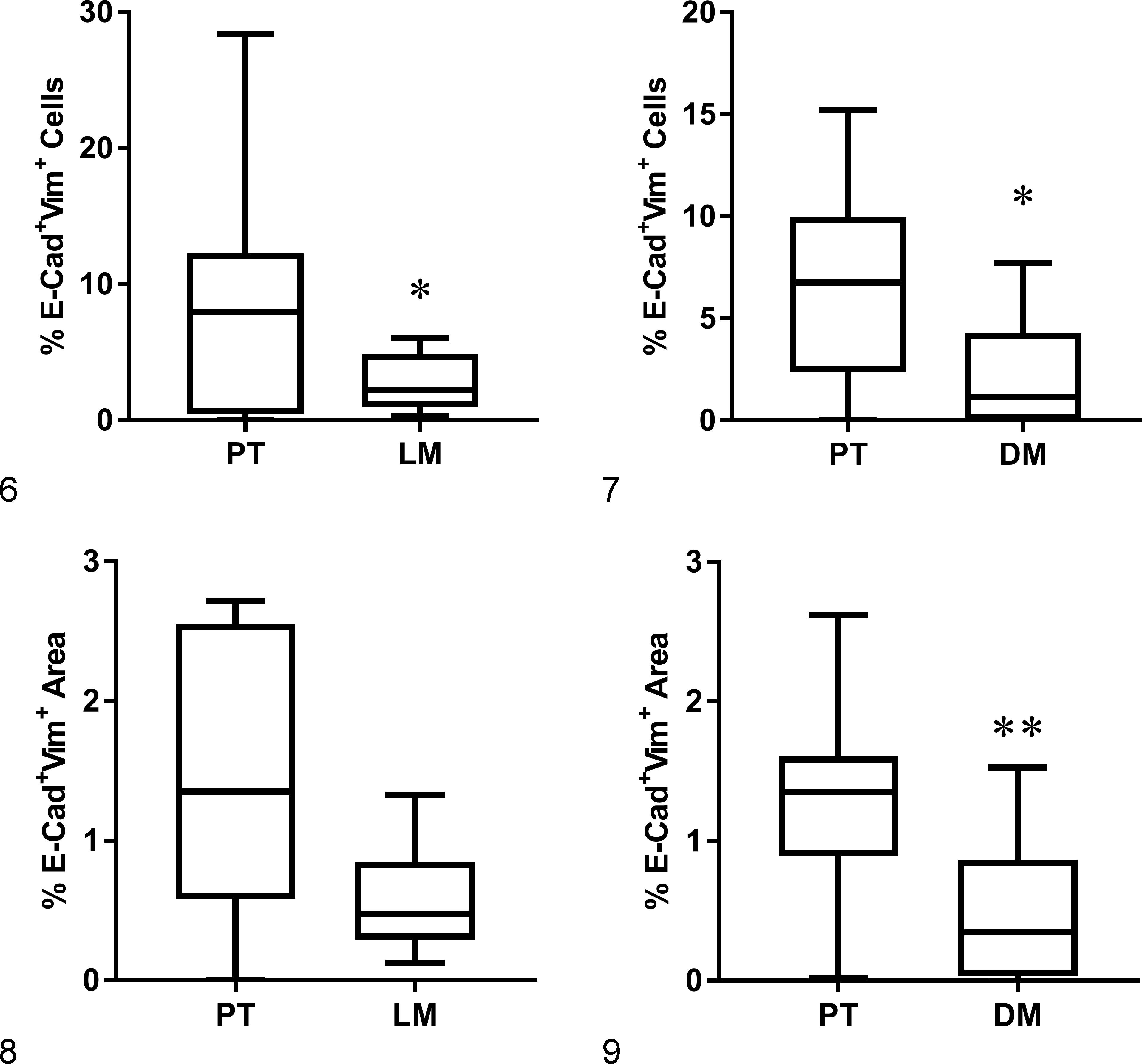
To identify cells expressing both epithelial and mesenchymal markers transitioning through the process of EMT/MET, E-cadherin and vimentin co-localization was assessed in the neoplastic epithelial cells. Pairwise comparison between primary tumor and matched lymph node metastasis or distant metastasis from individual dogs showed significantly more E-cadherin+/vimentin+ cells/area when comparing the primary tumor with its respective distant metastasis when analyzing by percentage of cells or area of co-localization, while a pairwise comparison between primary tumor and lymph node was significantly different only when the analysis was performed by cell counting (Table 3; Figs. 6–9). Thus, based on cell counting and computer-assisted analysis (ImageJ), E-cadherin and vimentin co-localization was higher in primary tumors compared with respective distant metastases.
We extended our analysis of E-cadherin and vimentin expression with respect to both number (cell counting) and area (ImageJ) between grades I, II, and III primary tumor samples (Table 4).
An overall low number of cells with membranous E-cadherin+/vimentin
Analysis of SNAIL/SLUG Expression by Immunohistochemistry
To further characterize the process of EMT in canine mammary carcinoma primary tumors and metastatic lesions, we assessed the relationship of E-cadherin+/vimentin+ cells with the core regulators SNAIL/SLUG. Nineteen of 20 primary tumors expressed SNAIL/SLUG with a median percentage immunopositivity of 30.0%, whereas all of the lymph node metastases (median 25.5%) and all of the distant metastases expressed SNAIL (median 10.0%). Pairwise comparison between matched samples showed a significantly lower percentage with SNAIL/SLUG expression in the distant metastasis compared with the primary tumor (P = .05; Table 3). When assigning positivity as SNAIL/SLUGlow (0 and +1) versus SNAIL/SLUGhigh (+2) categories according to Gamba et al., 17 there was no difference between tumor locations, with 7 of 20 (35%) in primary tumors, 2 of 10 (20%) in lymph node metastases, and 1 of 16 (6.2%) in distant metastases in the SNAIL/SLUGhigh category (chi-square test; P = 0.11).
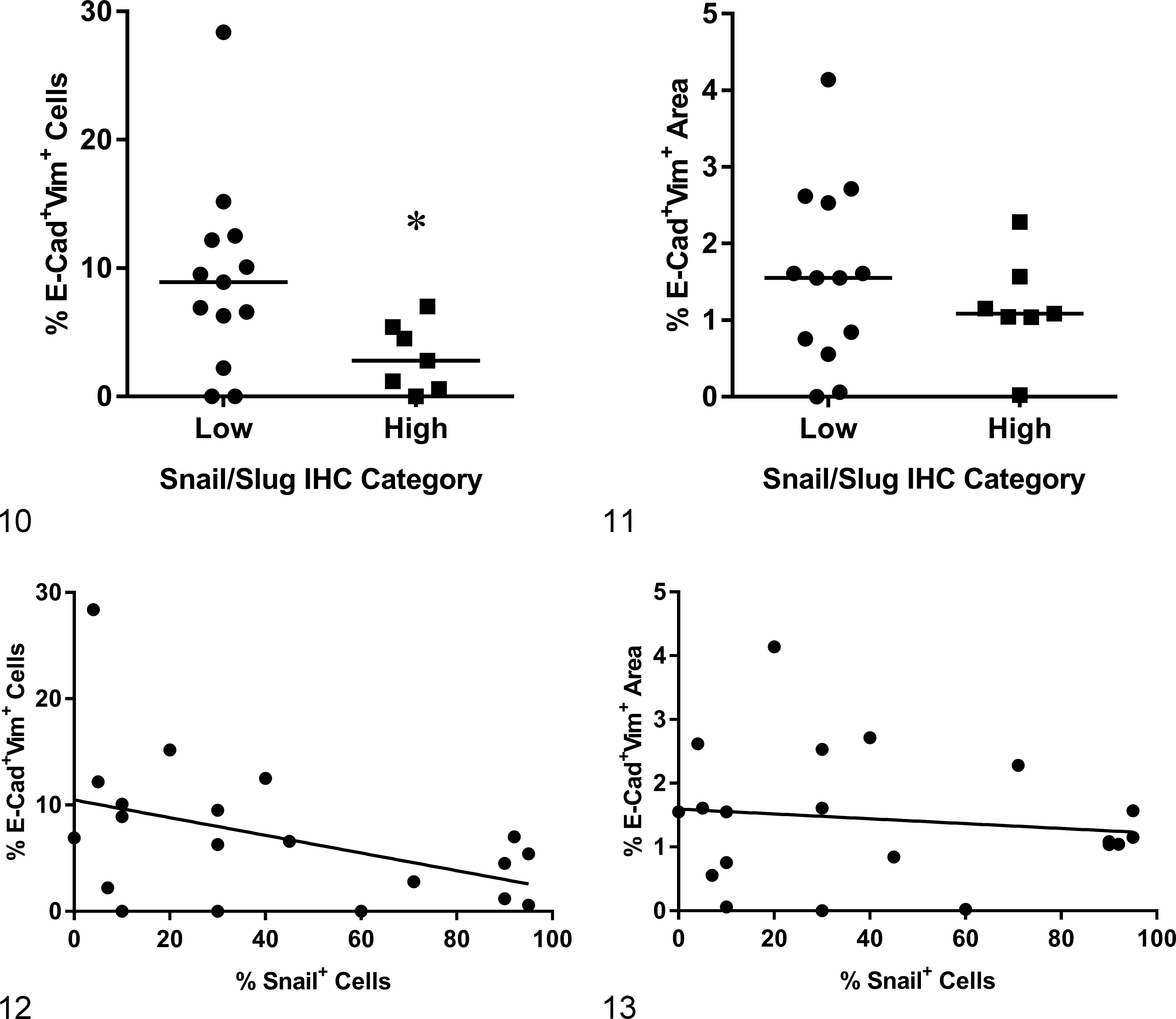
Interestingly, an inverse relationship between E-cadherin+/vimentin+ and SNAIL/SLUGhigh was found in primary tumors. Specifically, SNAIL/SLUGhigh primary tumors had significantly fewer E-cadherin+/vimentin+ cells than SNAIL/SLUGlow primary tumors (P = .0453; Fig. 10). However, when the area of E-cadherin+/vimentin+ co-immunoreactivity was evaluated (expressed as a percentage of the total area), no statistical significance was noted (Fig. 11). When analyzing the association between percentage of SNAIL/SLUG+ cells and E-cadherin+/vimentin+ cells, no significant correlation was noted (cell counting, P = .06, nor percentage area, P = .59; Figs. 12 and 13). Similarly, there was no association between grade and SNAIL/SLUG immunostaining.
Discussion
In human breast cancer, loss of E-cadherin is associated with high-risk clinicopathological characteristics, including increased tumor size, higher grade, 22,53 lymph node and distant metastasis, 53 and decreased survival. 39,45,53 Similarly, down-regulation of E-cadherin in canine mammary tumors has also been associated with a poor prognosis in several studies. 10,16,17,25,41,68 We found a surprisingly low number of cells that were positive for membranous E-cadherin in primary tumors, irrespective of grade, compared with previous literature. Notably, all the cases selected for this study had documented metastatic disease. This may explain the aberrant staining pattern (cytoplasmic) of E-cadherin in our study, if the low membranous but high cytoplasmic E-cadherin expression indicates a loss of membranous E-cadherin or a transitioning of E-cadherin from the cell membrane into the cytoplasm, which occurs as an early event in EMT. In support of this hypothesis, aberrant E-cadherin immunostaining has been reported as significantly more common in patients with metastatic gastric carcinomas than in patients without metastasis. 37 Cytoplasmic E-cadherin staining has also been demonstrated in invasive lobular carcinomas in women 21,34 and in less-differentiated mammary canine carcinomas. 55
Vimentin expression is typically expected in mesenchymal cells. 15 Its expression in neoplastic epithelial cells is consistent with the EMT process. 40,52,68 In breast cancer, vimentin expression has been associated with the triple-negative (estrogen receptor, progesterone receptor, and HER2-negative) molecular subtype, aggressive behavior, and a poor prognosis. 23,52,56,62 Previous studies have evaluated epithelial and mesenchymal markers in separate analyses and found an increase in vimentin and a decrease/loss of E-cadherin associated with higher grade and worse outcomes. ‡ Our study found a relatively low overall incidence of E-cadherin–/vimentin+ cells regardless of location (primary tumor, lymph node, or distant metastases) or grade (Tables 3 and 4). This may be because the lower-grade tumors included in this study all had metastasis and therefore were not representative of the typical non-metastatic grade 1 tumors.
Importantly, a significant increase in co-expression of E-cadherin and vimentin in the primary tumor compared with their paired distant metastases was noted, when evaluated through both cell counting and analysis of area of co-localization. This observation is consistent with the diversity in histologic types, grade, and biological behavior between primary mammary carcinomas and illustrates the unique advantage of using dogs with available matched metastatic lesions for breast cancer research. These findings support the theory that EMT is a dynamic process characterized by a changing phenotype from the primary tumor to the metastatic site. In fact, the lower incidence of E-cadherin+/vimentin+ cells in distant lesions relative to their primary tumors may support a reversal of the process, MET. When comparing grade and immunostaining pattern, we found, not surprisingly, a positive correlation between grade and E-cadherin+/vimentin+ phenotype; specifically, higher-grade tumors (grade II and III) had significantly more E-cadherin+/vimentin+ cells than grade I tumors. Histologic grade is an important prognostic factor in canine mammary tumors. 8,9,20,29,31,48,61 Four of the cases included here, however, developed regional and/or distant metastasis despite having grade I tumors. Further studies are needed to evaluate E-cadherin+/vimentin+ phenotype as a prognostic marker in low-grade tumors to identify dogs at risk for metastasis that may benefit from adjuvant therapies. The results from this current study, which confirm the presence of E-cadherin+/vimentin+ double-positive cells in all gross metastatic mammary carcinomas examined, provide the scientific rationale for performing such a study.
Multiple drivers are involved with the EMT process. In this study, we chose to evaluate SNAIL/SLUG. The transcriptional factors SNAIL and SLUG belong to the SNAIL superfamily of zinc finger transcriptional repressors that are known to negatively regulate E-cadherin transcription. 2,46,49 No difference in SNAIL/SLUG expression between primary tumor and its respective regional and distant metastases was observed when analyzed according to group (primary tumor, lymph node, distant metastases). However, when performing pairwise analysis based on percentage-positive SNAIL immunostaining, a significant difference between primary tumor and distant metastases was noted, again underscoring the value of using paired samples (Table 3). Interestingly, an inverse correlation between SNAIL/SLUGhigh expression and E-cadherin+/vimentin+ was noted; specifically, primary tumors with higher levels (>50% of cells) of SNAIL/SLUG staining had significantly fewer E-cadherin+/vimentin+ cells, and SNAIL/SLUGlow tumors had a higher number of E-cadherin+/vimentin+ cells. These results were somewhat unexpected. As a driver of EMT, one might theorize that SNAIL/SLUG up-regulation would result in an increased number of E-cadherin+/vimentin+ cells. Our results suggest that these events do not happen synchronously but rather in succession and that SNAIL expression precedes the transitioning from an epithelial to a mesenchymal phenotype; this in fact could be expected given its role as a transcriptional repressor of E-cadherin. Further mechanistic studies are needed to understand the role and interaction of the many transcriptional factors in EMT.
Previous literature also shows mixed results regarding the association between E-cadherin and SNAIL/SLUG. Gamba et al. 17 documented a direct correlation between E-cadherin down-regulation with SNAIL up-regulation in spontaneous canine invasive micropapillary mammary carcinoma, while Im et al. 25 did not confirm these findings in canine mammary tumors. Similar to Im et al., 25 we found no correlation between E-cadherin and SNAIL/SLUG. Lastly, in contrast to other studies both in human and veterinary breast/mammary tumors, where increased expression is associated with negative prognostic factors and poor outcomes, we did not find any correlation between SNAIL/SLUG expression and histologic grade. 25,26,44
The positive correlation between E-cadherin+/vimentin+ cells and higher-grade tumors, as well as differences between primary tumors and their respective metastases, provides further support that EMT plays a critical role in metastasis of canine mammary carcinoma. The difference between primary tumors and metastatic lesions suggests that this is a dynamic reversible process. However, given that excisional biopsies taken during surgery or those obtained at necropsy show only a snapshot illustration of a dynamic process, it is difficult to completely elucidate the direct interactions between the key mediators and the complex mechanisms involved with this approach. Nevertheless, the results support the presence of EMT in mammary carcinoma primary tumors and their metastatic lesions in spontaneously occurring mammary tumors and provide additional support for the development of prognostic biomarkers of EMT to identify patients at high risk for metastatic disease and poor outcomes. Further research to determine the predictive value of E-cadherin+/vimentin+ staining needs to be performed to confirm whether this method can be incorporated into routine diagnostics in canine mammary carcinomas and will suggest whether EMT may provide a potential target to improve outcomes in these patients.
Supplemental Material
Supplemental Material, DS1_VET_10.1177_0300985818776054 - Characteristics of the Epithelial-Mesenchymal Transition in Primary and Paired Metastatic Canine Mammary Carcinomas
Supplemental Material, DS1_VET_10.1177_0300985818776054 for Characteristics of the Epithelial-Mesenchymal Transition in Primary and Paired Metastatic Canine Mammary Carcinomas by Talita M. M. Raposo-Ferreira, Becky K. Brisson, Amy C. Durham, Renee Laufer-Amorim, Veronica Kristiansen, Ellen Puré, Susan W. Volk, and Karin Sorenmo in Veterinary Pathology
Footnotes
Acknowledgments
This research was made possible through the generous sharing of tissue block from dogs with metastatic mammary tumors, including Penn Vet Diagnostic Laboratory, School of Veterinary Medicine, University of Pennsylvania; The Department of Pathology, School of Veterinary Medicine, Norwegian University of Life Science; and The Norwegian Veterinary Institute. We would like to thank Dr Ken Drobatz for advice regarding statistical analyses.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Penn Vet Shelter Canine Mammary Tumor Program School of Veterinary Medicine, University of Pennsylvania, through funding provided by the Petco Foundation and the Blue Buffalo Foundation. Additional funding was provided by the Jack and Mina Ebrahimi Foundation, as well as a grant from the Sao Paulo Research Foundation (FAPESP; grant 2015/09046-9).
Supplementary material for this article is available online.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.