
Abstract
Estrogen receptor α (ERα) has an important role in mammary carcinogenesis, prognosis, and treatment. In human and canine mammary cancer, the most aggressive tumors show loss of ERα expression, which in human breast cancer has been attributed to methylation of the cytosine followed by guanine (CpG) island within the estrogen receptor α gene (ESR1) promoter. This study aimed to investigate the role of ESR1 CpG island (CGI) methylation in ERα expression in canine mammary tumors. Twenty-one canine mammary samples were sorted into three groups: malignant tumor (n = 9), benign tumor (n = 8), and normal gland (n = 4). Immunohistochemical analysis and reverse-transcription quantitative real-time PCR were performed to assess ERα expression and ESR1 mRNA levels. The methylation status was determined using sodium-bisulfite-treated DNA sequencing. All normal mammary glands and benign tumors showed high ERα expression (score range, 5–8). Six of the nine malignant tumors did not show ERα expression (score 0), two had score 2, and one had score 4. Lower ERα (P < .005) and ESR1 mRNA levels (P < .005) were found in malignant mammary tumors than in the other two groups. Canine ESR1 has an intragenic and non-promoter-associated CGI, different from humans. No significant variation in methylation percentage was observed among the groups, suggesting that ESR1 is not regulated by DNA methylation, unlike that in humans. This difference should be considered in further research using ERα as a biomarker for mammary tumors in canine studies on ERα-targeting therapy.
Breast cancer is the most prevalent neoplasm in women and female dogs. 26,38 Several factors are involved in mammary carcinogenesis, and exposure to the hormone estrogen is one of them. 40 The main role of estrogen in tumor initiation and progression is through its binding with estrogen receptor α (ERα). It promotes the transcription of proliferative genes and suppresses apoptotic genes, leading to cell proliferation and consequent DNA mutations arising from errors during cell replication. 46
The presence of ERα in human and canine mammary cancer has been widely reported, and owing to its relevance has become a prognostic biomarker and therapeutic target in human breast cancer. 41,43 However, the prognostic value of ERα is not yet well established in canine mammary tumor. Many studies have shown lower ERα expression in more aggressive mammary tumors. 8,23,25,28 However, the high variation in positive tumor percentage among the studies does not allow for the determination of a cut-off value for ERα positivity. 30 The loss of ERα expression in more aggressive tumors is also observed in human breast cancer, wherein it is mainly attributed to DNA methylation. 22,29,32
DNA methylation is an epigenetic event based on the addition of a methyl group in a DNA nitrogenous base, most commonly in a cytosine followed by a guanine (CpG). Regions of DNA with a high concentration of CpG dinucleotides are called CpG islands (CGIs). The methylation of gene-promoter-associated CGIs is related to gene silencing, but the role of CGIs in other gene locations, such as intragenic islands, remains unclear. 4 Owing to its capacity to regulate gene expression, DNA methylation is an important target in cancer studies. 9 The relevance of DNA methylation in cancer, including canine mammary tumors, has also been demonstrated in the veterinary field. 7,11,14
The goal of this study was to evaluate the expression of ERα protein and transcript and to compare them to the methylation status of the estrogen receptor α gene (ESR1) to investigate the regulation of ESR1 by DNA methylation in canine mammary tumors.
Materials and Methods
Samples and Histopathology
Seventeen canine mammary tumor samples and four normal mammary glands from the same mammary chain as the tumor samples were collected from the Veterinary Oncology Service, Veterinary Hospital, UFPR (Curitiba, Brazil). All patients had undergone clinical examination as well as thoracic radiography and abdominal ultrasonography for metastasis assessment. The type and histological grade of the tumor as well as the nodal status was identified by histopathological analysis. A sample weighing approximately 200 mg was frozen at –80°C before DNA and RNA extractions. All procedures were performed under the approval of the Ethics Committee on the Use of Animals at the Sector of Agricultural Science, Parana Federal University (Protocol No. 048/2015).
Tumor and normal gland tissues were fixed in 10% buffered formalin for 24–48 hours, embedded in paraffin wax, and stained with hematoxylin and eosin. Histologic classification followed the Goldschmidt classification, 12 and grading was performed according to the Elston and Ellis method. 10 Only tumors without myoepithelial proliferation or areas of metaplasia were included in this study.
Immunohistochemical Analysis
Immunohistochemical assays were performed using the tissue microarray (TMA) technique according to Schuler. 36 TMA sections of 3-μm thickness were mounted on charged slides. Immunohistochemical analysis was performed using the EnVision FLEX/HRP (Dako Code K8000, Carpinteria, CA, USA) detection method in an Autostainer Link 48 (Dako). Antigen retrieval was performed in a pretreatment tank PT Link (Dako) at 97°C by using the EnVision FLEX Target Retrieval Solution (Dako) for 20 minutes. The ready-to-use primary antibody monoclonal rabbit antihuman ERα clone EP1 (Dako) was incubated for 20 minutes at room temperature, following a specific protocol. The reaction was visualized using the EnVision FLEX DAB + chromogen system (Dako). Canine uterus tissue samples were used as the external positive control. Normal mammary gland tissues adjacent to the tumor and stromal cells were used as the internal positive and negative controls, respectively.
Protein expression was evaluated using the Allred scoring system, 30 wherein the final score is the sum of the percentage of the labeled cells and the intensity of the reaction scores. The final score can vary from 0 or 2 to 8 (Supplemental Table 2).
RNA Isolation and Reverse-Transcription Quantitative Real-Time PCR (RT-qPCR)
Total RNA was isolated using TRIzol (Invitrogen, Carlsbad, CA, USA), according to the manufacturer’s protocol. The quality of extracted RNA was assessed using agarose gel electrophoresis, and the quantity was measured using the Nanodrop Spectrophotometer (Thermo Scientific, Waltham, Ma, DE, USA). We included only those samples that presented an RNA 260/280 ratio around 2.0 and an RNA 260/230 ratio of 2.0–2.2 in the Nanodrop Spectrophotometer analysis and showed no signs of degradation on agarose gel electrophoresis. Reverse transcription reactions were performed using 1 μg of RNA in the High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA), according to the manufacturer’s protocol.
RT-qPCR for ESR1 and for the reference genes RPS18 and HPRT was performed using the SYBR Green PCR Master Mix (Applied Biosystems) containing 1 μL cDNA (1:100) and 150 nM primer. The reactions were performed in duplicate in 96-well plates by using a Step-One Plus Thermal Cycler (Applied Biosystems). A standard curve was generated for each primer. The sequences of all the primers are provided in Supplemental Table 1. Relative expressions of the mRNA were normalized using the geNORM3.4 software. 31,45
ESR1 CGI Methylation Analysis
To determine the CGIs in ESR1, the promoter region and the gene sequence obtained from GenBank (NCBI Reference Sequence: NC_006583.3) were submitted to the CpGPLOT program (https://www.ebi.ac.uk), an online software that identifies CGIs. Primers for CpG island amplification were designed using the Methprimer software (http://www.urogene.org/methprimer). The primers amplified a 421-bp fragment containing 50 CpG dinucleotides.
Genomic DNA was isolated from frozen samples by using the phenol/chloroform protocol. Sodium bisulfite treatment of DNA was performed using the EpiTect Bisulfite Kit (Qiagen, Valencia, CA, USA), according to the manufacturer’s instructions. PCR was performed with 1 μL of DNA, as described below: 1 cycle of 95°C for 5 minutes; 35 cycles of 94°C for 45 seconds, 48°C for 30 seconds, and 72°C for 1 minute; and 1 cycle of 72°C for 5 minutes. The products were electrophoresed on a 1% agarose gel and stained with ethidium bromide. PCR products were purified from the gel by using the PureLink Quick Gel Extraction Kit (Invitrogen), cloned in a pGEM T-easy vector (Promega, Madison, WI, USA), and introduced in DH10B competent Escherichia coli by electroporation. Eight recombinant clones of each sample were selected by α-complementation, and plasmid DNA was collected using the QIAprep Spin Miniprep Kit (Qiagen). Plasmid DNA sequencing was performed using Big Dye Terminator technology (Applied Biosystems) on an ABI 3500 sequencer (Applied Biosystems). The global methylation percentage was calculated by dividing the number of methylated CpG dinucleotides by the total number of CGIs analyzed.
Data Analysis
Statistical analyses were performed using the software GraphPad Prism v.5.0 (GraphPad Software, La Jolla, CA, USA). Gene expression was evaluated using Student’s t-test. The χ2 exact test was performed to determine the significance of protein expression among the groups. A P value < .05 was considered significant.
Results
Sample and Histopathology
According to the histopathologic examination, the samples were sorted into three groups: normal mammary gland or control group (n = 4), benign mammary tumor (n = 8), and malignant mammary tumor (n = 9). All benign tumors were histologically classified as simple adenoma, and the malignant tumors were classified as solid carcinoma (n = 4) or papillary carcinoma (n = 5). All patients with malignant tumors presented metastasis; four patients had metastasis to the lung, liver, or ovary and eight to the lymph node (Table 1).
Pathological Features and Estrogen Receptor α Protein and mRNA Expression in Canine Mammary Carcinoma Samples.

Immunohistochemical Analysis and RT-qPCR Assay
The immunohistochemical assay for ERα revealed nuclear labeling in the glandular acini and ductal cells (Figs. 1–3). Six of the nine carcinomas did not show any reactivity, and were thus given score 0. Two carcinomas were given score 2 (dogs 2 and 3), and one was given score 4 (dog 1). All normal and benign tumor samples showed scores higher than 5. Samples with a score 0 were considered ERα-negative, while those with scores of 2 or higher were considered ERα-positive (Table 2). The malignant tumors presented significantly more ERα-negative samples than did the control group and benign tumors (P < .005, χ2 test).
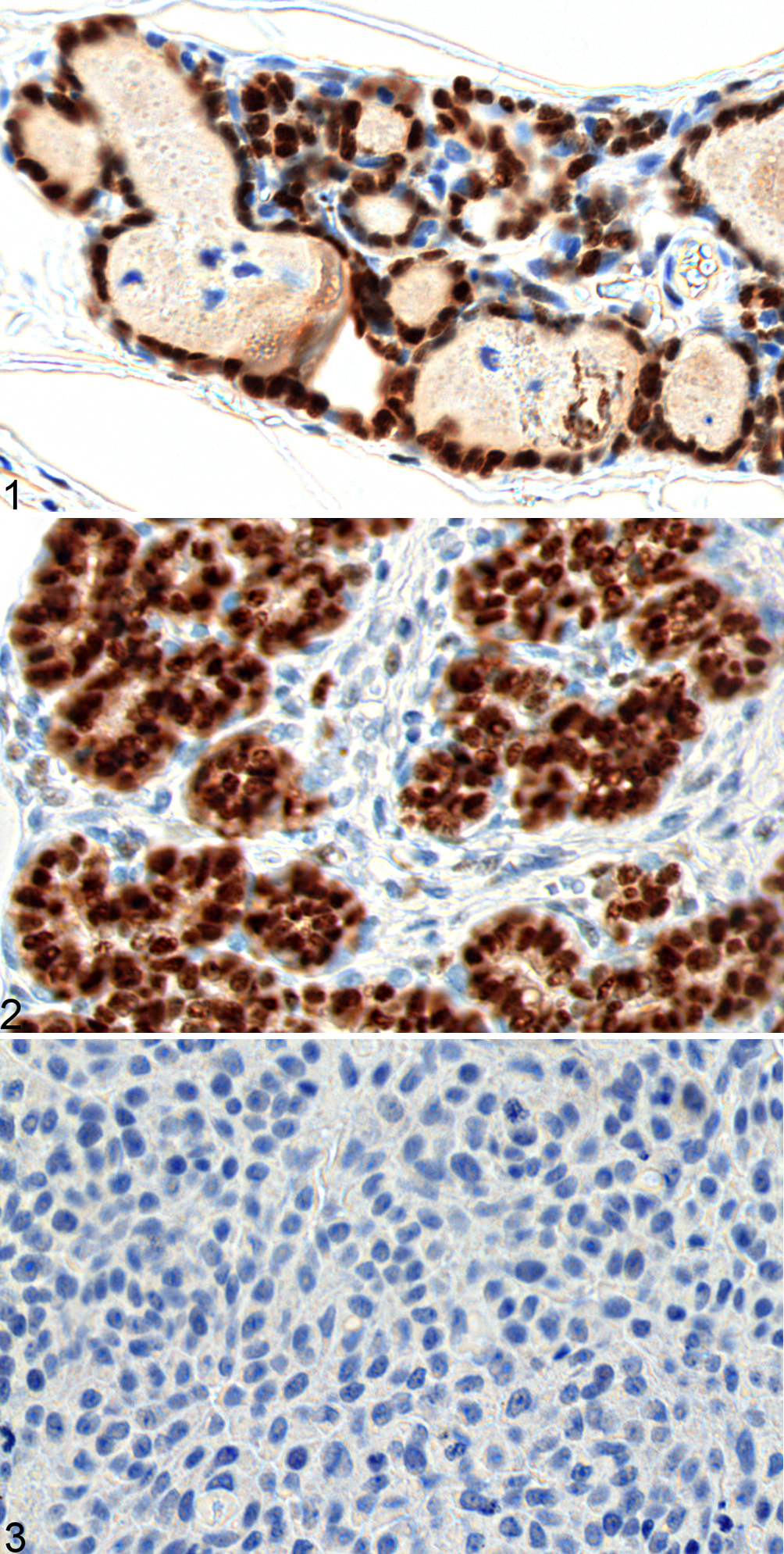
Estrogen Receptor α Expression Based on Immunohistochemistry in Normal Canine Mammary Glands, and in Benign and Malignant Mammary Tumors.
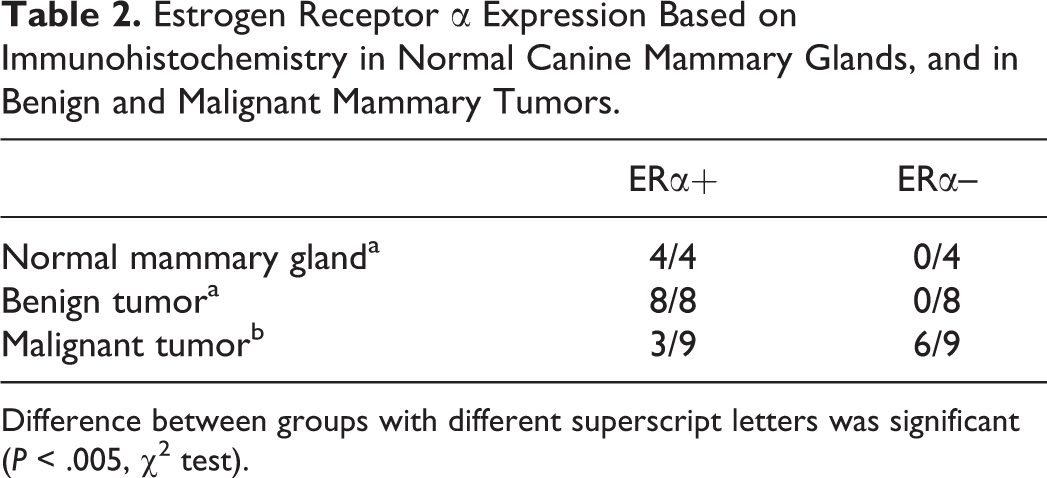
Difference between groups with different superscript letters was significant (P < .005, χ2 test).
The mRNA expression of ESR1 showed a positive correlation with protein expression. The mean relative expression of the malignant tumors (0.51; Fig. 4) was significantly lower (P < .005; Student’s t-test) than that of the control samples or benign tumors (0.89 and 1.0, respectively). A carcinoma sample that received a score 4 in the immunohistochemical assay (dog 1) showed the highest transcription expression among the malignant tumors. Three carcinomas (50%) with a protein score 0 (dogs 4, 5, and 6) had a similar relative mRNA expression as carcinomas with a score 2 (dogs 2 and 3). The other three malignant tumors (50%) with a score 0 (dogs 7, 8, and 9) presented the lowest transcript levels among all the samples.
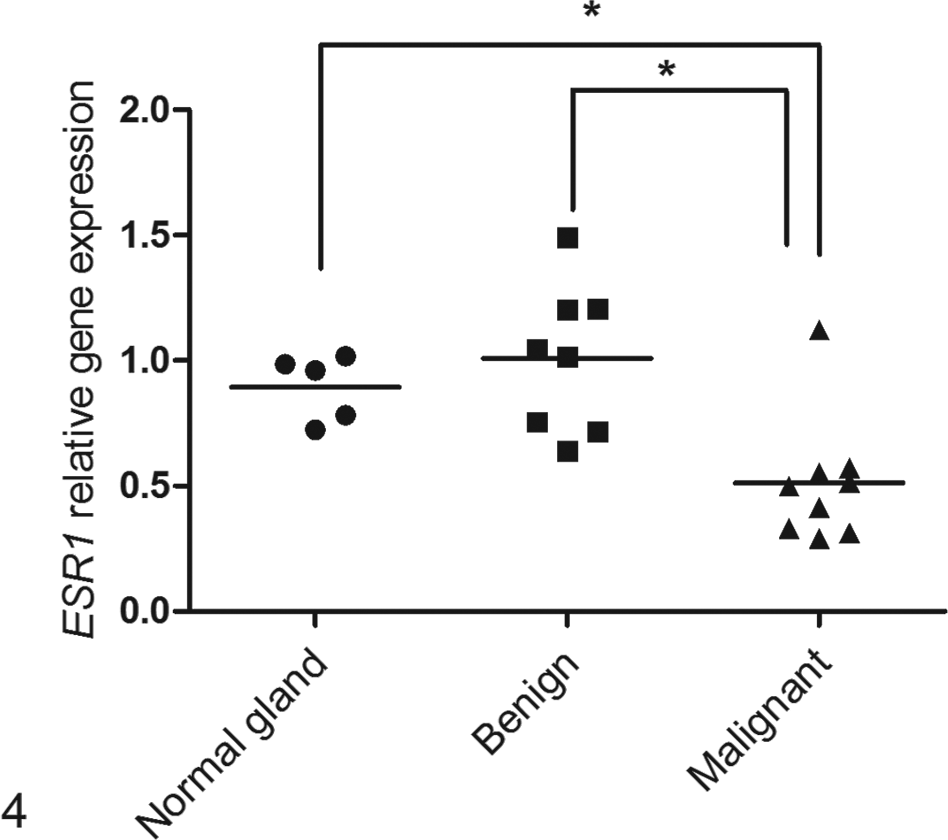
Relative expression of the ESR1 gene for the normal mammary gland (n = 4), benign tumor (n = 8), and malignant tumor (n = 9) groups. Bars: median value for each group. *P < .005, Student’s t-test.
Methylation Analysis
An in silico search comparing the human and canine ESR1 genes revealed a different pattern of CGI distribution. In humans, this gene presents a long CGI that extends from the promoter region to the first exon; however, in dogs, three intragenic CGIs were found (Fig. 5).
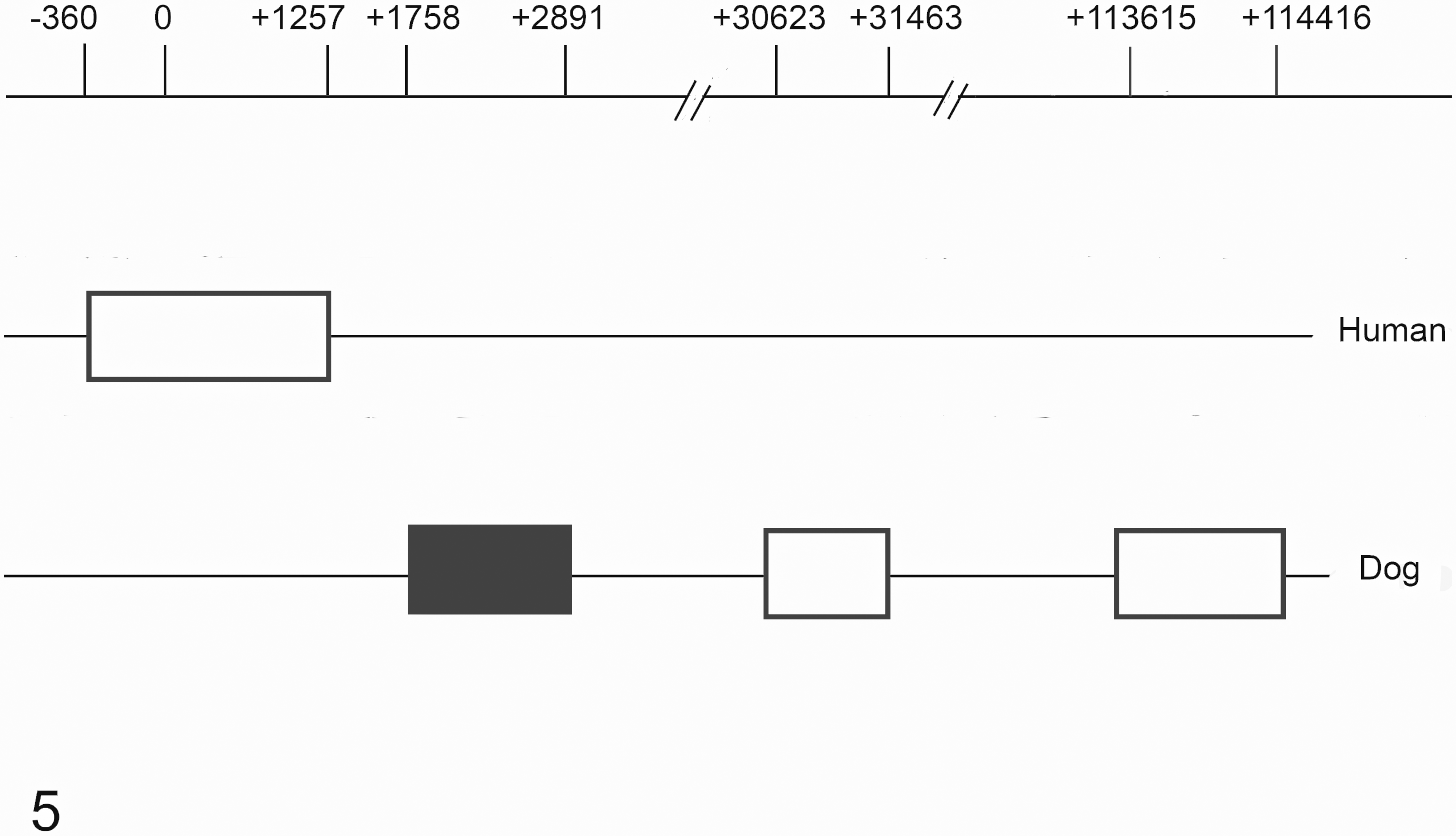
Distribution of CpG islands within the ESR1 gene in human and dog genomes. On the top line 0 represents the transcription start site (TSS). Black and white boxes: CpG islands. Black box: CpG island studied in this work.
In dogs, the closest island to the transcript start site (TSS) extends from the +1758 to +2891 position and comprises part of intron 1, exon 2, and part of intron 2. This CGI covers an alternative TSS (NCBI Reference Sequence: XM_014114137.1) located at position +1882 and the ATG site. The other two islands are found within introns 2 and 4. These were not included in this study. No CGI was found in the gene-promoter region or up to 2000 base pairs upstream of the TSS. The sequenced fragment of the CGI contained 50 CpG dinucleotides and covered the putative alternative TSS and the ATG site.
To analyze the correlation between protein expression and DNA methylation, the groups were sorted again according to their ERα status. The normal mammary glands formed the control group, while the other samples were divided into the ERα-positive or ERα-negative groups, independent of the tumor classification (benign or malignant). Methylation analysis showed a general hypomethylation in all groups. Slightly higher methylation rate was observed in the ERα-negative group (2.6%) than in the ERα-positive group (1.9%), but the methylation rates were virtually identical in the ERα-negative group and normal mammary gland group (2.5%). The methylation rate of each CGI sequenced was established (Fig. 6).
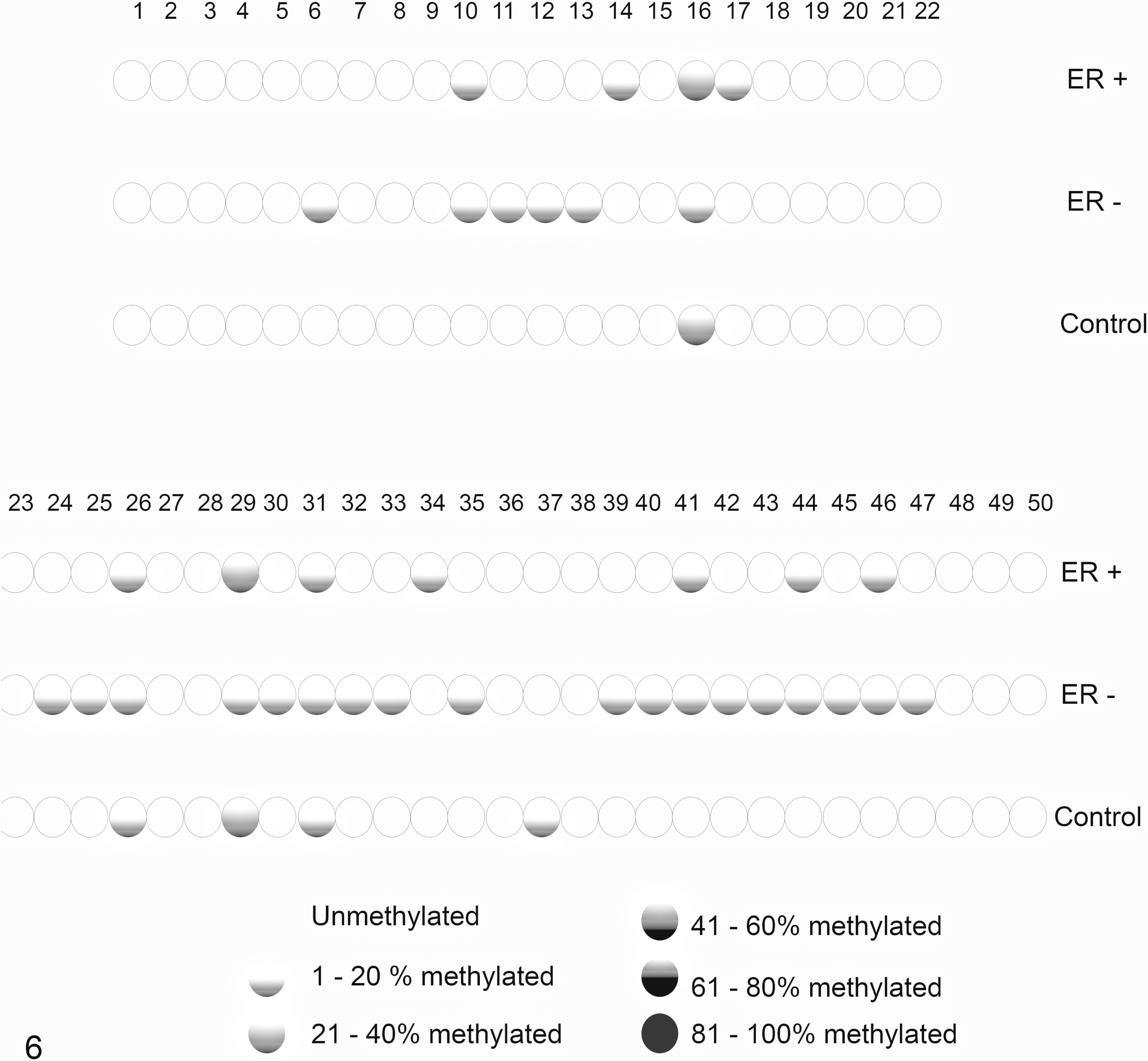
CpG island methylation. Each circle represents a CpG dinucleotide. The gradation from white to black circles corresponds to higher methylation percentages of the CpG, such as shown at the bottom right of the figure. ER+, estrogen receptor-positive group; ER–, estrogen receptor-negative group; Control, control group.
When individually analyzed, CpGs 16 and 29 were more methylated in the ERα-positive and control groups than in the ERα-negative group. Although we searched for transcription factors that could bind to these CpG sites, we could not identify any relevant candidates.
Discussion
The importance of the estrogen receptor in mammary gland tumorigenesis and cancer progression is well known. 46 Many studies have found lower ERα expression in more aggressive canine mammary tumors. 11,23,25,28 The results of immunohistochemical analysis in the present study are consistent with those in the literature; that is, carcinomas presented lower protein scores than did benign tumors and normal mammary glands. Regarding the mRNA expression of ESR1, the results reported in the literature are controversial. Some studies 18,20 did not find any difference in ESR1 mRNA expression among normal mammary glands, benign tumors, and primary or metastatic carcinomas, whereas another study reported lower significant mRNA expression in mammary carcinomas than in normal mammary glands, but not in benign tumors. 42 Our study, in turn, showed a significantly lower expression of the transcript in carcinomas than in adenomas and normal glands, a finding also described in the literature. 27
As expected, ESR1 transcript level corresponded to ERα protein expression. However, three of the six carcinomas that showed a score 0 for the ERα protein expressed ESR1 mRNA. This controversy was described by another author 22 who also found mRNA expression in ERα-negative samples. Similar data concerning ERα have been reported in human breast 15 and ovarian cancers. 19 This divergence among transcript and protein levels has been attributed to the variations in the immunohistochemical assay 17,27 techniques, such as antibody clones and fixation time of the tissue. 30 However, other factors such as posttranscriptional regulation should also be considered.
When the dog ESR1 was evaluated for CGIs, three intragenic CGIs were found, but none was found in the promoter region; this finding was different from that in human ESR1 (Fig. 5). The dog genome was reported to have a higher CGI density than that of many other mammals such as humans, mice, and some livestock such as cows, horses, sheep, and goats. 3,13 However, the number of promoter-associated CGIs in the dog genome is lesser than that in humans, 13 a feature that could be exemplified by CGI distribution in ESR1. The larger number of CGIs found in the dog genome can partially be attributed to the elevated recombination A/T to G/C alleles, which seem more dense in the vicinity of CGIs, thereby provoking a self-reinforcing process. 2 Thus, the relatively lesser number of promoter-associated CGIs in the dog genome could be a mechanism to avoid recombination in important functional elements. 2
While the hypermethylation of promoter-associated CGIs is well known to repress gene expression, the role of intragenic CGIs remains unclear. 4 Some studies have demonstrated that hypermethylation of CGIs located in the first 6,35 and third exons 39 is tightly related to gene silencing, whereas other authors have associated transcription repression to intragenic CGI hypomethylation. 1,21,37 Transcription silencing by intragenic CGI hypomethylation can be connected to the loss of repression of repetitive elements, such as retrotransposons. In the absence of methylation, these elements are amplified, and disrupt the associated gene mRNA processing. 1 Similarly, when an alternative promoter-associated CGI is originally methylated but becomes hypomethylated, it enables alternative TSS transcription and can alter the expression levels of isoforms. 5
The activity of the putative alternative TSS was not analyzed in this study; therefore, it remains unknown if the methylation could regulate its expression. However, we did not find a significant difference in methylation status between the control, ERα-positive, or ERα-negative groups, which suggests that even if this alternative promoter is related to canine mammary carcinogenesis, it is probably not regulated by DNA methylation.
The sequencing of CGIs in this study showed a slightly lower methylation rate (1.9%) in ERα-positive samples, and surprisingly, the control and ERα-negative groups shared a similar rate (2.5% and 2.6%). This unexpected result could probably be attributed to the stromal cells that are present in larger numbers in normal mammary glands. These cells do not express ERα and probably have their own epigenome, different from the parenchymal cells.
The absence of different methylation levels between the group with high ERα expression and the ERα-negative group suggests that in canine mammary tumor, ESR1 is not regulated by DNA methylation. In humans, however, one of the regulatory mechanisms of ESR1 is the methylation of a promoter-associated CGI. 29 Many studies on human breast cancer have reported an important association between ESR1 promoter methylation and low ERα expression, mainly in invasive, ductal, poorly differentiated, and triple-negative tumors. 22,32 This correlation was also found between ESR1 methylation in free-circulating cells and tumor protein expression. 24
Because of many similarities between canine and human cancers, such as spontaneous occurrence, biological behavior, and molecular pathways, 33,44 dogs have been considered promising models for human cancer research. 33,34 These similarities have also allowed the development of veterinary cancer studies based on well-established human cancer biomarkers and therapeutics. 16 However, some differences still remain between these two species, such as those presented in this study. While in human breast cancer, ESR1 is regulated by DNA methylation, it does not seem true in canine mammary tumors. Thus, the difference in ESR1 regulation between canine mammary tumors and human breast cancer can be a reason why establishing ERα as a canine mammary tumor biomarker has been difficult.
In conclusion, this study revealed a lower expression of ESR1 in more aggressive canine mammary tumors; moreover, this gene does not seem to be regulated by DNA methylation in canine mammary tumors unlike in human breast cancer. Despite the many similarities between canine and human breast cancer, this study presents an important difference between these two species, and highlights the need for comparative research. Additional studies on ESR1 regulation are needed to confirm the use of ERα as a prognostic biomarker in canine mammary tumor and to validate the use of the dog as a model for studying ERα-targeting therapies in humans.
Supplemental Material
Supplemental Material, DS1_VET_10.1177_0300985818763711 - DNA Methylation Status of the Estrogen Receptor α Gene in Canine Mammary Tumors
Supplemental Material, DS1_VET_10.1177_0300985818763711 for DNA Methylation Status of the Estrogen Receptor α Gene in Canine Mammary Tumors by Yara de Oliveira Brandão, Mariana Busato Toledo, Andressa Chequin, Thierry Grima Cristo, Renato Silva Sousa, Edneia Amancio Souza Ramos, Giseli Klassen in Veterinary Pathology
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Supplementary material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.