
Abstract
The ataxia telangiectasia mutated (ATM) gene encodes a protein associated with DNA damage repair and maintenance of genomic integrity. In women, ATM transcript and protein downregulation have been reported in sporadic breast carcinomas, and the absence of ATM protein expression has been associated with poor prognosis. The aim of this study was to evaluate ATM gene and protein expression in canine mammary tumors and their association with clinical outcome. ATM gene and protein expression was evaluated by reverse transcription-quantitative polymerase chain reaction and immunohistochemistry, respectively, in normal mammary gland samples (n = 10), benign mammary tumors (n = 11), nonmetastatic mammary carcinomas (n = 19), and metastatic mammary carcinomas (n = 11). Lower ATM transcript levels were detected in benign mammary tumors and carcinomas compared with normal mammary glands (P = .011). Similarly, lower ATM protein expression was observed in benign tumors (P = .0003), nonmetastatic mammary carcinomas (P < .0001), and the primary sites of metastatic carcinomas (P < .0001) compared with normal mammary glands. No significant differences in ATM gene or protein levels were detected among benign tumors and nonmetastatic and metastatic mammary carcinomas (P > .05). The levels of ATM gene or protein expression were not significantly associated with clinical and pathological features or with survival. Similar to human breast cancer, the data in this study suggest that ATM gene and protein downregulation is involved in canine mammary gland tumorigenesis.
Breast cancer is the most prevalent malignant disease among women. 10,15 Compared with other animals, female dogs have a high incidence of mammary gland tumors, 3 times higher than that in women. 9,19,21,22,27,38 A high homology between human and female dog mammary tumors has been observed with regard to the natural occurrence, sites of metastasis, hormone dependence, and pathways involved in tumorigenesis. 22,34,36
In Brazil, the estimated prevalence of malignant canine tumors exceeds 70%, 31,32 higher than that reported in the United States, where the rate is approximately 50%. 28,38 In dogs with malignant mammary tumors, the overall 2-year survival rate is 25% to 40%; however, this survival rate can be influenced by many factors, such as the histological type and grade, tumor stage, and treatment choice. 38
The ataxia telangiectasia mutated (ATM) gene, a primary transducer in DNA damage response pathways, is mainly activated by double-stranded breaks that, if not repaired, can lead to genomic rearrangements and chromosomal instability, contributing to tumorigenesis. 5 ATM encodes a protein kinase that when activated recruits and phosphorylates several proteins involved in DNA injury responses. 12,23
The pivotal role of ATM in the DNA damage response is related to cell cycle control, apoptosis, gene regulation, oxidative stress, and telomere maintenance. 25,30 ATM gene and protein downregulation has been reported in human sporadic breast carcinomas. 8,41,47
ATM may also be involved in chemoresistance and radioresistance mechanisms. 40 Radiation therapy plays an important role in breast cancer management in women, 20 and the major cause of radiation treatment failure is associated with therapy-induced radioresistant tumor cells, which exhibit increased DNA repair abilities. 20 Acquired resistance of tumor cells to ionizing radiation or chemotherapy is the major barrier to curing breast cancer and improving cancer survival in women. 40
It was recently suggested that ATM protein expression is an independent prognostic marker in human breast cancer with clinical significance for cancer prognosis. 8 However, to our knowledge, there are no similar studies in dogs. Therefore, the aims of this study were to evaluate ATM gene and protein expression levels in canine mammary tumors, to correlate them with clinical outcome, and to compare the results from female dogs and women. In addition to a better understanding of the role of ATM in canine mammary tumorigenesis, this approach may help direct new therapeutic strategies in both species.
Materials and Methods
Subjects
All procedures were performed under the approval of the Ethics Committee on the Use of Animals of FCAV, UNESP, Jaboticabal, Sao Paulo, Brazil (protocol No. 028129/10). Forty-one canine mammary tumor samples and 10 normal mammary glands were collected. Mammary gland tumors were obtained from the Veterinary Oncology Service, Veterinary Hospital, UNESP (Jaboticabal, SP, Brazil) during mastectomies; normal mammary gland samples were obtained from the Araçatuba Zoonosis Control Center (Araçatuba/SP, Brazil). Clinical examination included an evaluation of tumor size, the presence or absence of muscle adherence, tumor consistency, ulceration, local inflammation, and nodal status (confirmed by histopathology). Three-view thoracic radiographs and abdominal ultrasound examinations were used to assess metastatic disease. Clinical staging was obtained by the evaluation of tumor size, nodal status, and metastasis according to a modified World Health Organization clinical staging system as follows: stage I (T1N0M0), stage II (T2N0M0), stage III (T3N0M0), stage IV (TanyN1M0), and stage V (TanyNanyM1). 39
ATM gene expression was evaluated in fresh-frozen mammary gland tumors collected during surgery. A paired sample collected from the same tumor was formalin fixed and paraffin embedded to assess ATM protein expression. All cases were reviewed and reclassified independently by two veterinary pathologists. The histological classification was performed according to Misdorp et al 28 and complemented by the proposal of Cassali et al 9 (Table 1), and tumor grade I, II, and III was assigned according to Elston and Ellis. 13
Histological types of 41 canine mammary tumors.

Of 41 canine mammary tumor samples, 11 were identified as benign tumors and 30 as primary mammary carcinomas. Of those with mammary carcinomas, 19 were nonmetastatic and 11 had metastases. Metastasis affected the regional lymph nodes (5 cases), lungs (2 cases), lymph node and lung (1 case), liver and spleen (1 case), lymph node and skin (1 case), and lymph node and adrenal gland (1 case). ATM gene and protein expression data were compared with clinical and histopathologic information and survival time only in the carcinoma group (n = 30).
All dogs were subjected to mastectomy as the primary treatment and followed prospectively by clinical examination, Complete blood cell count, biochemistry profile, 3-view thoracic radiographs, and abdominal ultrasound were performed every 3 months, with a mean follow-up of 13.80 ± 6.4 months (1 to 26 months).
Analysis of ATM Gene Expression
Total RNA was extracted (TRIzol, Invitrogen Life Technologies, Carlsbad, CA, USA) from the primary tumor according to the manufacturer’s instructions from tissue samples having at least 80% neoplastic cells. The RNA samples were quantified (Nano-Drop ND-8000 spectrophotometer, Thermo Scientific, Wilmington, DE, USA), and the RNA quality was analyzed (Agilent 2100 Bioanalyzer 6000 Nanochip, Agilent Technologies, Waldbronn, Germany). cDNA synthesis was performed in a final volume of 20 μL containing 1 μg of total RNA treated with DNase I (Life Technologies, Rockville, MD, USA), 200 U of SuperScript III reverse transcriptase enzyme (Life Technologies), 4 μL of 5X SuperScript First-Strand buffer, 1 μL of dNTPs (10 mM each; Life Technologies), 1 μL of Oligo-(dT)18 (500 ng/μL; Life Technologies), 1 μL of random hexamers (100 ng/μL; Life Technologies), and 1 μL of DTT (0.1 M; Life Technologies). Reverse transcription was carried out for 60 minutes at 50°C, followed by inactivation for 15 minutes at 70°C. The cDNA was stored at –80°C.
Reverse transcription-quantitative polymerase chain reaction (RT-qPCR) to determine the expression of ATM and the reference genes (Supplemental Table S1) was performed in a total volume of 11.5 μL containing Power SYBR Green PCR Master Mix (Applied Biosystems, Foster City, CA, USA), 1 μL cDNA (1:10), and 0.3 μL of each primer. The reactions were performed in duplicate in 96-well plates using a StepOne Plus Thermal Cycler (Applied Biosystems). A dissociation curve was included in all experiments to determine PCR product specificity. The most stable reference genes (RPS5, RPS19, and ACTB) were selected as suggested by Brinkhof 7 and identified using geNorm software 44 from a group of 5 genes (ACTB, HPRT, RPS5, RPS19, and RPL8; Supplemental Table S1). Relative gene quantification was calculated using the 2–▵▵CT method. 26
Analysis of ATM Protein Expression
For immunohistochemistry, 5-μm-thick sections of tissues from the primary tumor were mounted on charged slides (Starfrost adhesive microscope slides; Lowestoft, UK), antigen retrieval was performed by heating for 40 minutes with Tris-EDTA buffer (pH 9.0) in a water bath at 96°C, and the endogenous peroxidase activity was blocked with 8% hydrogen peroxide solution in methanol. An anti-ATM polyclonal rabbit primary antibody (clone ab47575; Abcam, Cambridge, MA, USA) was applied at a 1:100 dilution, followed by incubation with a polymer system (Advance HRP, Dako, Dako Denmark A/S, Glostrup, UK). 3,3′-Diaminobenzidine chromogen (DAB Substrate System, Dako) was applied for 5 minutes. The slides were then counterstained with Harris hematoxylin (Sigma-Aldrich, St. Louis, MO, USA). Negative control slides were prepared by omitting the primary antibody and by using rabbit immunoglobulin instead of the primary antibody.
ATM protein expression was evaluated based on the distribution of positively immunostained tumor cells and by using the following scores: 1 (<25% positive cells), 2 (26% to 50% positive), 3 (51% to 75% positive), and 4 (>76% positive). 11 Staining intensity in mammary gland cells was also evaluated as follows: 1 = weak staining, 2 = strong staining. The product of the distribution and intensity was calculated for each sample (Supplemental Table S2), 2,24,42 and the results were compared with normal mammary gland (adapted from reference 29).
All normal mammary tissues had scores of 4 for distribution and 2 for intensity, resulting in a product score of 8 (intensity × extension). Reduced protein expression was considered in mammary tumors with lower scores.
Data Analysis
Statistical analyses were performed using GraphPad Prism v.5.0 (GraphPad Software, La Jolla, CA, USA). Kruskal-Wallis or Mann-Whitney U tests were applied to compare ATM transcription levels among the samples. The χ2 exact test was used to determine the significance of the association between ATM protein expression in the canine mammary tissue samples and clinical-histopathological characteristics. The survival rate was calculated using the Kaplan-Meier method, and statistical significance was determined using a log-rank test. Overall survival was defined as the period (in months) between the date of surgery and death caused by the disease. P < .05 was considered significant.
Results
Significantly decreased ATM transcript levels (by RT-qPCR) were detected in both benign and malignant tumors compared with normal samples (P = .011; Fig. 1). However, no differences in ATM levels were observed between benign mammary tumors and mammary carcinomas or between nonmetastatic and metastatic carcinomas (P > .05). Similarly, no association was found between ATM levels and clinical parameters, such as tumor size, clinical stage, nodal status, or histopathological type or grade (Supplemental Fig. S1–S4).
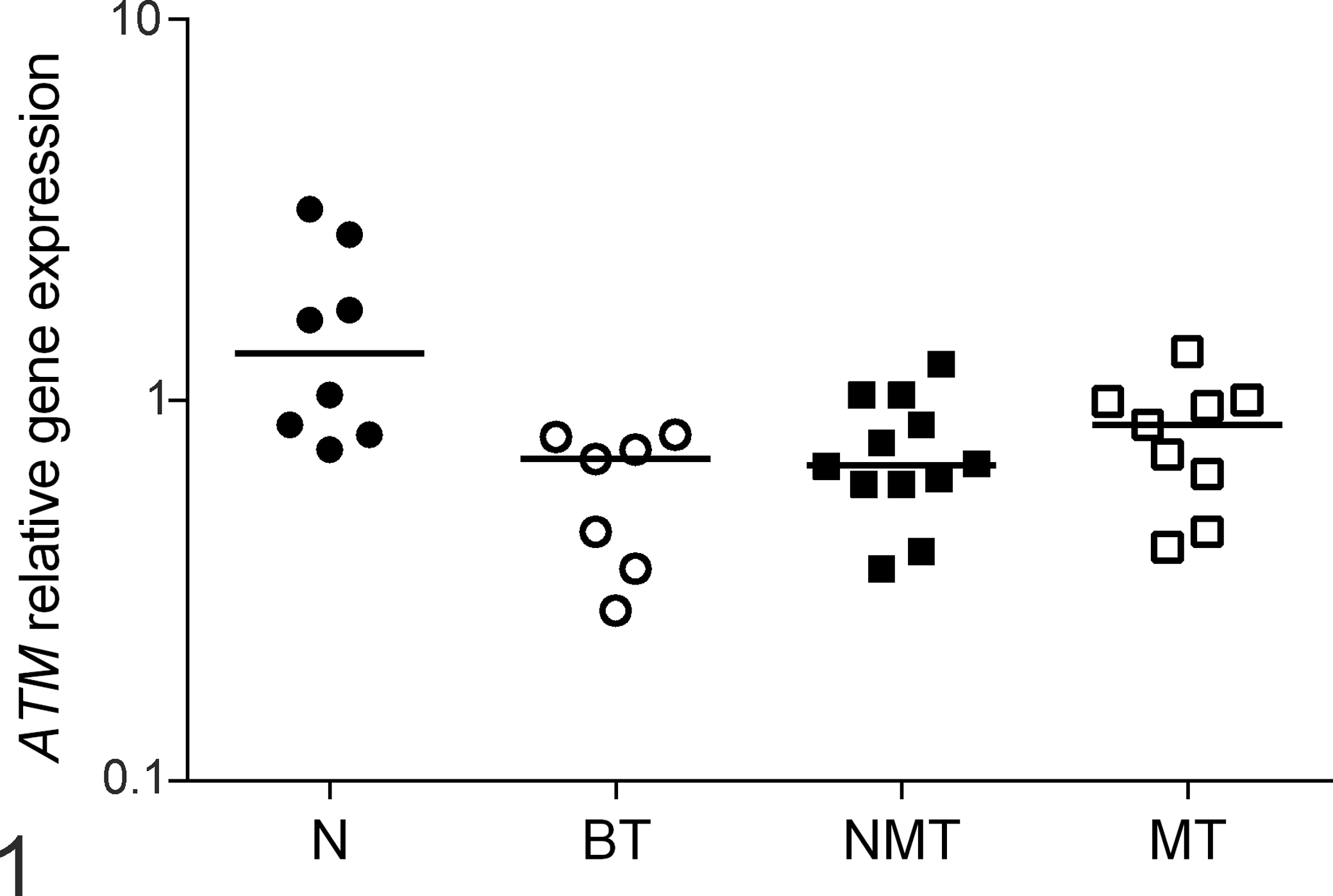
Canine normal mammary glands (N), benign mammary tumors (BT), nonmetastatic mammary carcinomas (NMT), and the primary sites of metastatic mammary carcinomas (MT) were evaluated for ataxia telangiectasia mutated (ATM) gene expression using reverse transcription-quantitative polymerase chain reaction. Bars: median value in each group. P = .011, Kruskal-Wallis test. Samples with unreliable results were excluded from analysis.
Immunohistochemistry for ATM revealed cytoplasmic and membranous staining (Fig. 2–5). Compared with normal mammary glands, lower ATM protein levels (score <8) were detected in benign mammary tumors (73% of samples, P = .0003) and in nonmetastatic (84%, P < .0001) and metastatic (100%, P < .0001) mammary carcinomas (Table 2). However, no significant differences were observed among benign tumors and nonmetastatic and metastatic mammary carcinomas (P > .05).
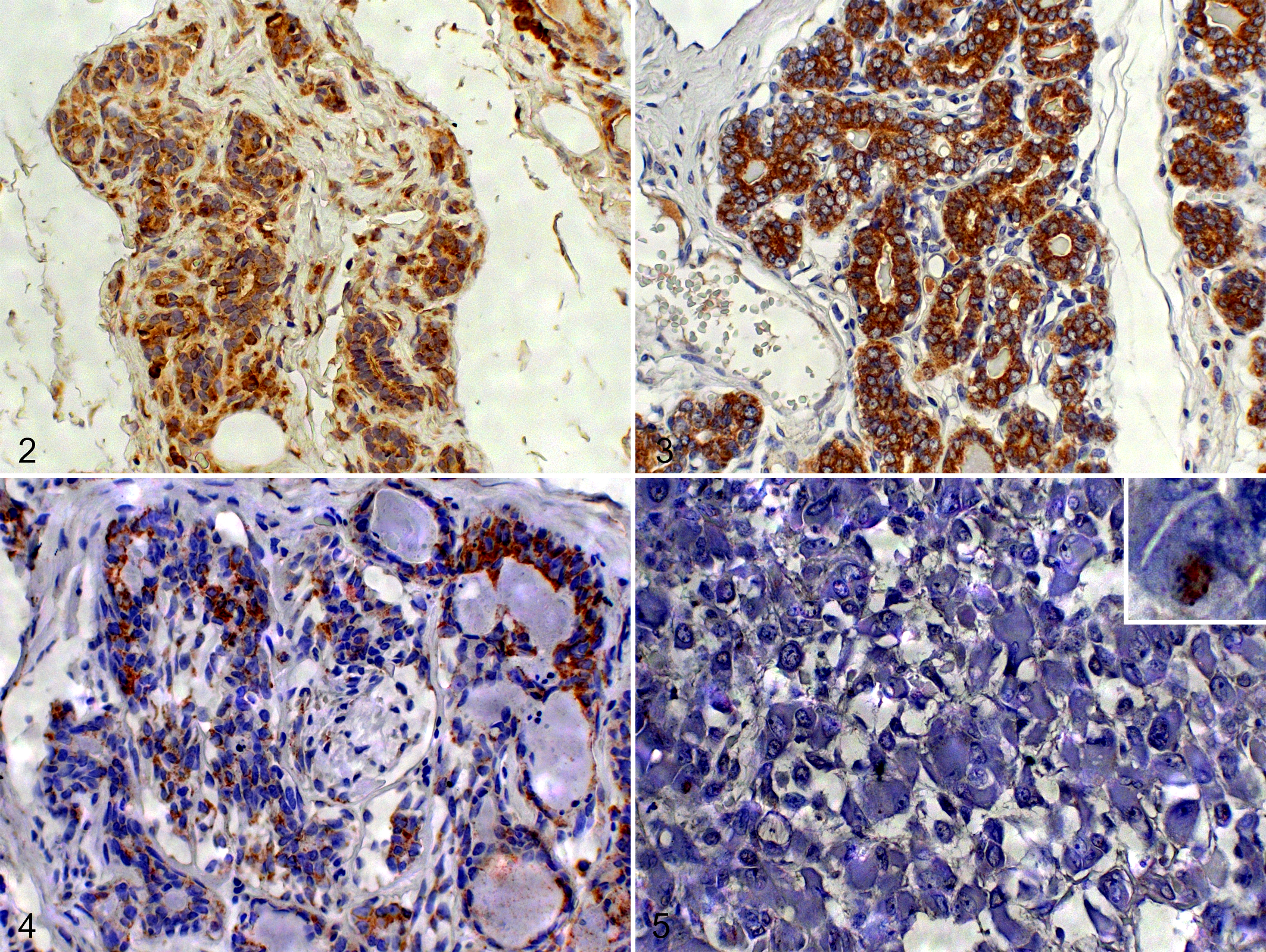
Mammary gland, dog. Immunohistochemistry for ataxia telangiectasia mutated (ATM) protein, cytoplasmic labeling.
ATM protein expression based on immunohistochemistry scores in 41 canine mammary tumors and 10 normal mammary glands.a
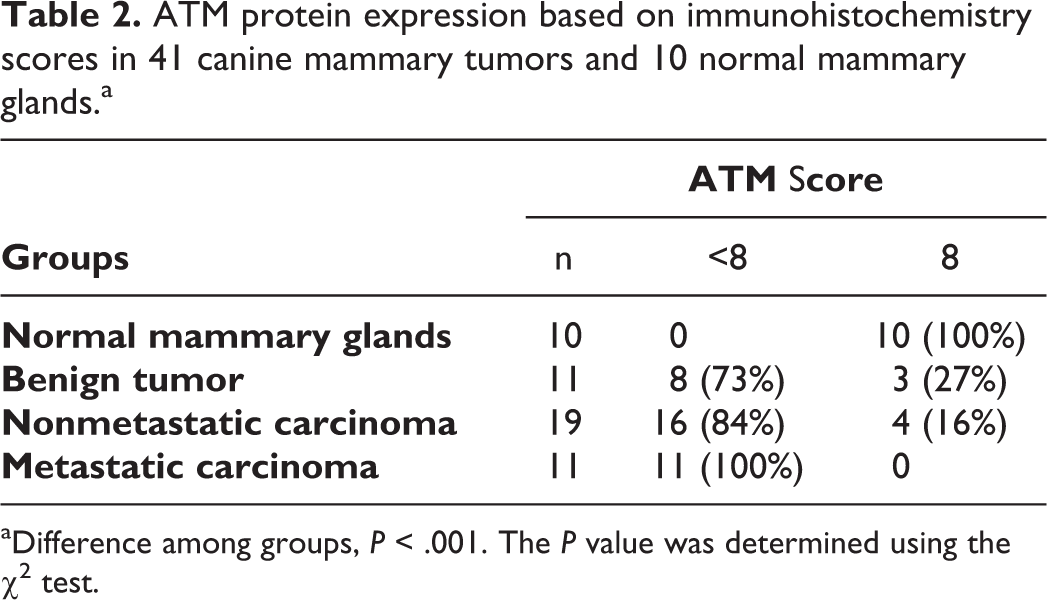
aDifference among groups, P < .001. The P value was determined using the χ2 test.
Clinical-histopathological parameters were also evaluated according to ATM protein expression, and no association was found (P > .05; Supplemental Table S3). Reduced survival was observed in dogs that had tumors with reduced ATM protein (score <8), but this difference was not statistically significant (Fig. 6).
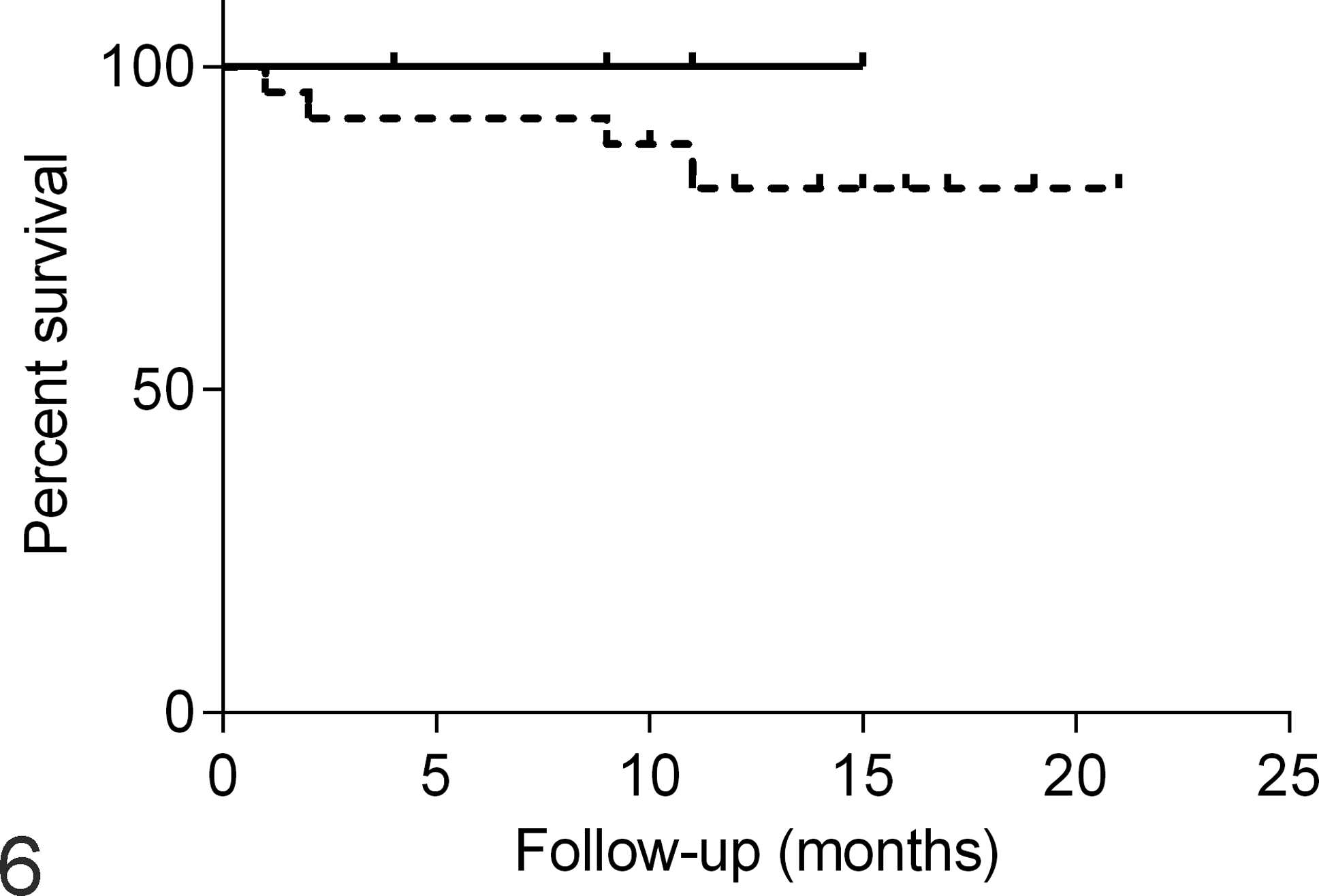
Survival time curve of female dogs with mammary carcinoma comparing those with normal (solid line) and reduced (dashed line) ataxia telangiectasia mutated (ATM) protein expression based on immunohistochemistry scores of 8 or <8, respectively. P = .61, log-rank test.
Paired protein and transcript expression was assessed in 21 mammary carcinomas (12 nonmetastatic and 9 metastatic), and concordance between gene and protein expression was observed in 4 cases: 2 cases presented normal levels by both procedures, and 2 cases showed gene and protein underexpression. Two nonmetastatic samples presented ATM gene underexpression and a normal ATM protein expression score. Normal ATM transcript levels were detected in 17 carcinomas, but ATM protein underexpression was detected in 15 (7 metastatic and 8 nonmetastatic).
Discussion
ATM is a highly conserved gene among many species (chimpanzee, rhesus monkey, frog, cow, mouse, rat, chicken, zebrafish, and dog), and its mRNA sequences present 89% similarity between humans and dogs. The canine ATM protein contains FAT-, PI3K-, PI4K-, FATC-, and ABL-1–binding domains, with greater than 90% similarity to the human protein. 1,16 Here, we demonstrated lower ATM gene and protein expression in canine mammary tumors compared with normal mammary tissues, as previously described in human sporadic breast cancer. 3,4,6,8,41,47 In women, the mechanisms involved in ATM deregulation remain unknown. 35 Several mutations in ATM have been described and were associated with a moderate risk of breast cancer development, 14,35 but these are rare and frequently associated with hereditary breast cancer. 18 Another possible mechanism involved in ATM downregulation is epigenetic silencing, although studies on ATM methylation status have shown contradictory results. 45 ATM posttranscriptional regulation by microRNAs is an additional hypothesis for ATM control. 37 Furthermore, the dysregulation of DNA repair networks associated with ATM/Chk2 can lead to the appearance of recurrent and metastatic cancer cells. 33
Canine benign mammary tumors and mammary carcinomas showed reduced ATM gene expression when compared with normal tissue, but no difference in ATM gene levels was observed among the neoplastic groups. One study 47 found similar ATM mRNA levels in human benign tumor tissues and adjacent normal mammary tissues, whereas another study 46 detected moderately decreased ATM transcripts in benign mammary tumors and suggested a correlation with disease development and/or the malignant progression of breast cancer. Our findings suggest a role for ATM expression in canine mammary tumor development but not with disease progression, as verified in breast cancer in women. However, additional studies are needed to understand the functional role of ATM in the development and progression of the canine mammary tumors.
A weak correlation between ATM gene and protein expression levels was verified in carcinoma samples; Bueno et al 8 described similar results in human breast carcinomas. Posttranscriptional regulation mechanisms associated with the ATM gene could be responsible for the discrepancy detected between transcript and protein expression, and the loss of ATM gene expression by microRNA regulation might be involved in ATM deregulation in human breast cancer. 8,37 Thus, it is possible that altered posttranscriptional mechanisms can also explain our data in canine carcinomas and should be further explored.
By performing global transcript analysis of canine mammary tumors and comparing the results with gene expression in human breast tumors, Uva et al 43 have shown that most prognostic signatures are strongly correlated in both species and have emphasized the high potential value of dogs as a preclinical model to test therapeutic agents targeting pathways that are commonly deregulated in mammary tumors in both dogs and women. Based on this finding and the fact that ATM inactivation may predict responses to PARP inhibitors, 17 female dogs with mammary gland tumors are acceptable for use in preclinical trials because reduced ATM gene and protein expression is observed in mammary tumors. Further, the findings suggest that peroxisome proliferator-activated receptor inhibitor treatment might be of benefit in dogs with mammary cancer.
Our data suggest an influence of ATM transcript and protein levels on canine mammary gland tumorigenesis, suggesting a novel target for therapy and providing additional support for the consideration of dogs as suitable models for comparative studies of invasive breast cancer.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Sao Paulo Research Foundation (FAPESP), grant 2010/13777-5 and 2011/10279-7.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.