
Abstract
Clostridium haemolyticum causes bacillary hemoglobinuria (BH), an infectious and usually fatal disease that occurs mostly in cattle, which is clinically characterized by jaundice, hemoglobinuria, and anemia. The trematode Fasciola hepatica has been commonly reported as the main predisposing factor that triggers this condition. The authors evaluated 20 naturally occurring cases of bovine BH to characterize the pathology and pathogenesis of the disease. Grossly, the most consistent finding was a large, frequently single focus of necrosis surrounded by a red to purple halo, observed most frequently on the parietal surface of the right and left hepatic lobes. Other findings were jaundice, dark-brown discoloration of kidneys, and red urine in the urinary bladder. Microscopically, characteristic lesions were locally extensive, necrotizing hepatitis with thrombosis and numerous intralesional Gram-positive rod-shaped bacteria, and acute renal tubular necrosis. By immunohistochemistry, many hepatocytes outside the necrotic focus in the liver were positive for activated caspase 3, suggesting that those cells were undergoing apoptosis. Ultrastructural evaluation revealed hepatocyte necrosis, hemolysis, and clumps of vegetative and sporulating bacilli within the liver. Polymerase chain reaction for the C. haemolyticum beta toxin gene was positive in randomly selected liver samples. No gross or microscopic lesions indicative of fascioliasis were detected in the liver of any animal, suggesting that other yet undetermined predisposing factors were associated with these cases of BH.
Bacillary hemoglobinuria (BH) is a disease of cattle and occasionally sheep that was first described in 1916. 17 The disease occurs endemically in many South American countries and the western United States, and it has been sporadically reported in several other countries, including Canada, Australia, New Zealand, Ireland, Wales, Iran, Turkey, India, and Japan. 1,22,25,38 BH is caused by Clostridium haemolyticum (formerly known as Clostridium novyi type D), an anaerobic, motile, sporulating, rod-shaped bacterium, which is mainly found in soil of areas with poorly drained pastures and alkaline pH, where viable spores of the microorganism can survive for years. 19,34 Spores of C. haemolyticum have also been found, albeit rarely, in the liver, bloodstream, intestinal content, bone marrow, and kidneys of clinically healthy cattle. 13,26 BH is usually observed during summer and early autumn, being more common in areas with high prevalence of Fasciola hepatica infestation. 6,35,38 Clinical signs typically include hemoglobinuria, jaundice, reduction of rumination and appetite, fever, lethargy, milk drop, and production of blood-stained feces. 29,35 Animals with BH develop severe reduction of packed cell volume, red blood cell count, and hemoglobin concentration, and the leukocyte response may vary from leukopenia to leukocytosis. 33 The disease is almost always fatal. 35
The pathogenesis of BH has been mainly associated with the migration of the immature stages of F. hepatica and other trematodes through the liver parenchyma. The resulting local anaerobiosis triggers germination of spores of C. haemolyticum resident in the liver; toxin production by the resulting vegetative cells causes necrosis and other lesions characteristic of BH. 6,19 Beta toxin, a phospholipase C similar to the beta-lecithinase of C. novyi type B, is considered the main factor responsible for the virulence of the microorganism. Beta toxin hydrolyzes phosphatidylcholine into phosphocholine and a diglyceride, 10 inducing hemolysis, hepatocyte and endothelial cell damage, 2 platelet aggregation, and increased vascular permeability. 10
Even though the damage caused by liver fluke migration is commonly accepted as the most important trigger of the disease, BH has also been diagnosed, albeit rarely and in individual cases, in animals free from this parasite. 6,12 The predisposing factor in those cases has not always been determined.
The pathology of BH has been described in the past in individual cases of spontaneous disease and in larger numbers of experimentally inoculated animals. 21,33 In this retrospective study, we describe in detail the main gross, microscopic, and ultrastructural pathology, the mechanism of cell death, and other findings of spontaneously occurring BH in 20 animals.
Materials and Methods
Case Selection
Twenty-five archival cases submitted between 2001 and 2014 to the Eastern Regional Laboratory, Veterinary Laboratories Division Miguel Rubino (Treinta y Tres, Uruguay) had received a presumptive diagnosis of BH based on gross and microscopic pathology. From these, the 20 cases with the least autolysis were selected for further study. All animals involved in the selected cases were necropsied in the field by the attending veterinarian (F.D.) within 1 to 3 hours after death, and tissues were immediately collected and fixed by immersion in formalin. For this study, BH was confirmed through the detection of C. haemolyticum in the liver by polymerase chain reaction (PCR) and/or C. novyi by immunohistochemistry.
Gross and Microscopic Pathology
The gross findings were retrieved from the necropsy reports and photographs, which had been prepared by one of the authors of this study (F.D.). Special attention was paid to the gross liver lesions, including number, location, distribution, size, shape, consistency and color.
Samples of liver, kidneys, spleen, lungs, skeletal muscle, heart, and central nervous system were collected from all animals and fixed by immersion in 10% buffered formalin, pH 7.2, for 24 to 72 hours, and 4-μm-thick sections were prepared routinely and stained with hematoxylin and eosin (HE). Preliminary examination identified significant microscopic lesions in the liver, kidney, and spleen, and these organs were studied in detail in the 20, 11, and 3 animals, respectively, for which specimens were available. These specimens were routinely processed to produce 4-μm-thick sections stained with HE, Hucker and Conn’s Gram stain, and phosphotungstic acid hematoxylin (PTAH). In addition, selected sections of kidney were stained with Okajima stain for hemoglobin.
Sections including affected and unaffected portions of the liver of each animal were examined. The following liver lesions were semiquantitatively scored using an ordinal scale from – (absent) to ++++ (most severe): necrosis, inflammation, hemorrhage, congestion, hepatocyte vacuolation, thrombosis, and number of Gram-positive rods.
Immunohistochemistry
Sections of liver were processed by immunohistochemistry (IHC) using an indirect immunoperoxidase technique for C. novyi according to standard operational procedures (SOPs) of the California Animal Health and Food Safety Laboratory System (CAHFS). Briefly, the RTU Vectastain Elite ABC system (Vector Laboratories, Burlingame, California) was used according to the manufacturer’s instructions. A 3% solution of hydrogen peroxide was used to block endogenous peroxidase, followed by antigen retrieval with pepsin. Nonspecific binding was blocked by treating the samples with Background Punisher (Bicare Medical, Concord, California) before incubation with goat anti-C. novyi polyclonal antibody (VMRD, Pullman, Washington) for 60 minutes at 37°C. Vector NovaRED (Vector Laboratories) was used as chromogen for visualization. Tissues incubated with normal goat serum instead of primary antibodies were used as a negative control. Liver from a cow with BH and in which C. haemolyticum had been identified by culture and PCR was used as a positive control.
Additionally, sections of liver from 8 animals were processed by IHC for activated caspase 3, and kidney sections from 5 animals were processed for Leptospira spp IHC, using the Dako EnVision Kit (Dako, Carpenteria, California) according to the manufacturer’s instructions and following SOPs of CAHFS. The primary antibodies used were a rabbit polyclonal anti-activated caspase 3 antibody (ASP175, Cell Signaling Technology, Danvers, Massachusetts) and a rabbit polyclonal anti-Leptospira spp antibody (GenWay Bio, San Diego, California), both at a 1:100 dilution. The immunoreactivity was visualized with Vector NovaRED chromogen. Sections from a lymph node of a cow with lymphoma were used as positive controls for activated caspase 3 IHC. Kidney sections of a cow from which Leptospira spp had been isolated and tested positive for Leptospira spp by fluorescent antibody test (FAT) and PCR were used as positive controls for Leptospira spp IHC. Lymph node and kidney from a normal cow were used as negative controls for activated caspase 3 and Leptospira IHC, respectively. Additional negative controls for both techniques included tissues incubated with normal rabbit serum instead of the corresponding primary antibodies.
Transmission Electron Microscopy
Formalin-fixed liver samples from 3 study animals (Nos. 4, 17, and 19) were placed in half-strength Karnovsky’s fixative, and thin sections were processed for transmission electron microscopy (TEM). After aldehyde fixation, tissues were washed in 0.2 M sodium cacodylate, fixed again in 2% osmium tetroxide reduced with 2.5% potassium ferrocyanide, dehydrated in ethanol, and infiltrated and embedded in a formulation of Spurr’s epoxy resin. One-micrometer-thick sections were placed on glass slides, stained with toluidine blue, and observed by light microscopy. Thin sections were then obtained and mounted on 150 mesh copper grids, stained briefly with 6% methanolic uranyl acetate, post stained in Reynold’s lead citrate, and observed in a LEO 906E transmission electron microscope at 60 kV accelerating voltage. The length and width of 20 vegetative bacteria, spores, and their main structural components were measured; means and standard deviations were calculated for these values.
PCR on Formalin-Fixed, Paraffin-Embedded Tissues
Four 5-μm-thick sections of formalin-fixed, paraffin-embedded (FFPE) liver from 6 randomly selected study animals and from 1 normal cow were placed into microcentrifuge tubes for dewaxing by adding 1 ml of xylene, followed by centrifugation for 2 minutes at 12,000 × g. Then, the xylene was removed and the pellet was washed with 1 ml of 100% ethanol. After centrifugation at 12,000 × g for 2 minutes, the ethanol was discarded and the samples were air-dried at room temperature for 15 to 45 minutes. The dewaxed tissues were then subjected to DNA extraction by using a commercial kit (QIAamp DNA FFPE Tissue Kit; Qiagen, Hilden, Germany) following the manufacturer’s instructions. The extracted DNA was used as a template for conventional PCR detection of the C. haemolyticum beta toxin gene (GenBank AN: AF525415) using the following primers: 5′-ATTTTTGGGATCCTGACACA-3′ (ChbF) and 5′-CCCCAAAGTAATGTAAGCCT-3′ (ChbR), which amplify a 186 bp DNA fragment of the C. haemolyticum beta toxin gene. PCR was performed in a total volume of 25 μL containing 5 μL of extracted DNA, 0.25 μL of each primer (10 μM), 7 μL of nuclease free water, and 12.5 μL of PCR Master Mix 2X Promega (Madison, Wisconsin), which contains Taq DNA polymerase (pH 8.5, 50 units/ml), dNTPs (400 μM), and MgCl2 (3 mM). The following thermocycler profiles were used: 95°C for 4 minutes, 35 cycles at 94°C for 30 seconds, 55°C for 30 seconds, and 68°C for 1 minute followed by a final extension step at 68°C for 10 minutes and a final hold at 4°C. DNA extracted from a C. haemolyticum strain previously isolated from a cow with BH was used as positive control. Reactions in which study tissues were replaced by liver of a normal cow, or in which water was used instead of DNA or primers, were used as negative controls. PCR amplicons were visualized in ethidium bromide-stained 1% agarose gels (Agarose SFR; Amresco, Solon, Ohio).
Results
Clinical and Epidemiological Findings
Table 1 presents a list of the individual animals including signalment and epidemiological data. Briefly, male and female cattle of ages between 10 months to ≥4 years were available. Breeds included Hereford, Aberdeen Angus, and crossbreeds. Cases of BH occurred during all seasons of the year. The affected animals originated from mixed sheep and cattle farms exhibiting outbreaks of the disease, with morbidity ranging from 0.2% to 8.3%. Case fatality varied between 67% and 100%. Each animal presented here corresponded to a different outbreak. Farms included in this study were situated on the eastern plains of Uruguay, mostly rice-farming areas characterized by low, poorly drained soils. No diagnosis of F. hepatica infestation had been made in any of the farms or in the general area where the farms were located. No treatments were attempted on any of the 20 animals.
Signalment and Epidemiological Information of 20 Cases of Bacillary Hemoglobinuria in Cattle.
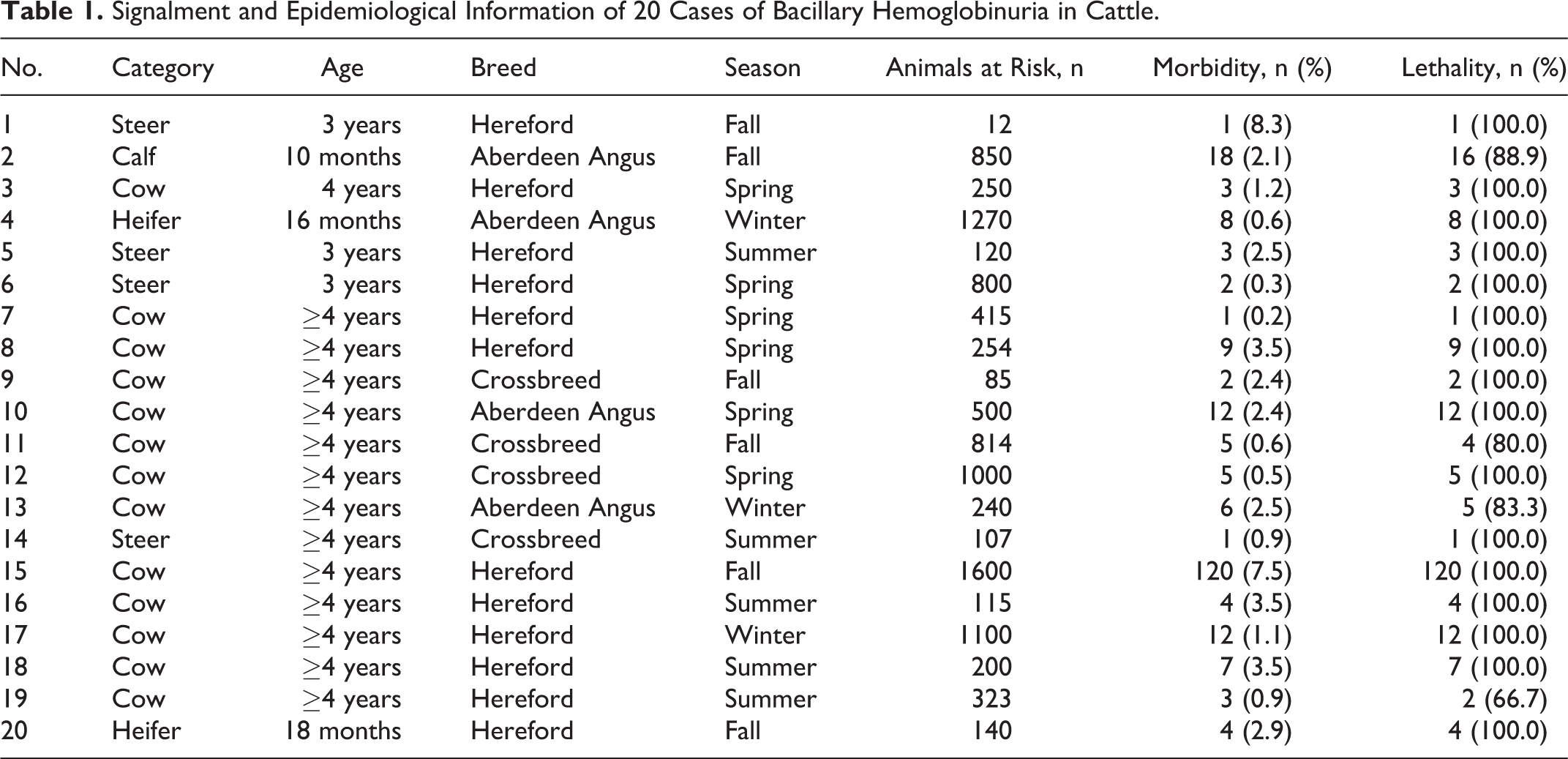
Gross Pathology
Gross lesions were similar in all animals but varied in severity and distribution. Externally, the tail and perineal area were stained by dark red feces. The subcutaneous tissue, muscular fascia, and serosa and adipose tissue surrounding internal organs had diffuse and intense yellow-orange discoloration. The topographic distribution and extension of the hepatic gross lesions were available in 14 animals, and these were summarized to create a schematic representation (Fig. 1). These lesions varied in size, affecting between 5% and 30% of the hepatic parenchyma. In most animals (10/14; 71.4%), the liver had a single and roughly wedge-shaped necrotic focus with its base on the parietal surface (Fig. 2); the other 4 animals had multiple (2–5) necrotic foci. One animal presented a single focus extending to include both hepatic lobes (Figs. 1 and 3). The foci of hepatic necrosis were in only the right hepatic lobe in 7 of 14 animals (50%), in only the left hepatic lobe in 4 of 14 animals (29%), and in both lobes in 3 of 14 animals (21%). These lesions tended to be localized in the central portion of the right lobe or in the ventral portion of the left lobe (Fig. 1). Regardless of location and size, the foci of necrosis were pale, soft to firm, and surrounded by a very thin red to purple halo. The liver capsule over the foci of necrosis was partly covered by a thin layer of fibrin. Thrombi were present in small and medium-sized veins located around and inside necrotic lesions, equally involving portal and hepatic veins. The rest of the liver was frequently friable and orange-brown (Fig. 2) probably due to icterus. No liver flukes or migration tracks were seen in the study animals.
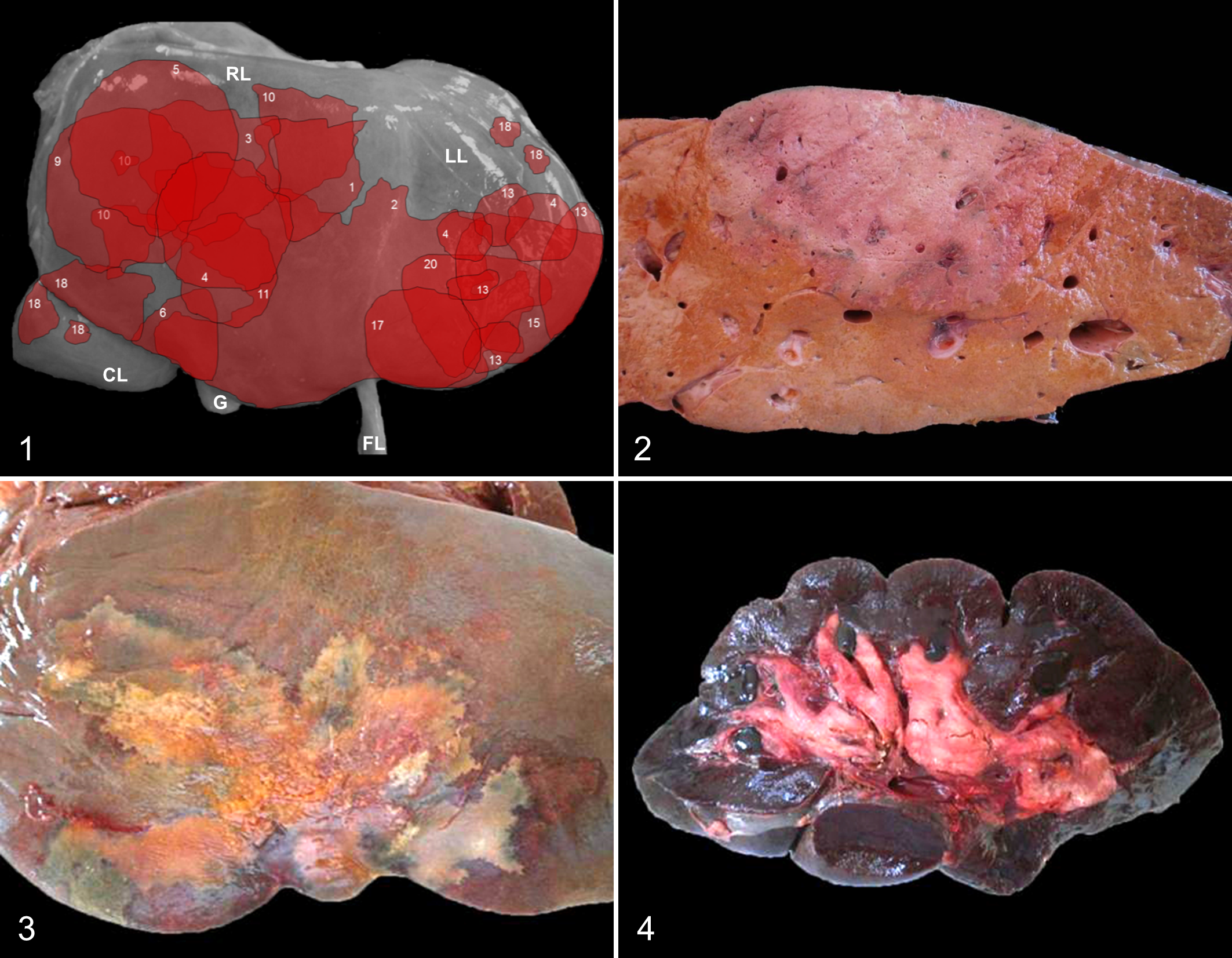
The renal cortex and medulla were mottled or diffusely dark-brown to black (Fig. 4), and the renal pelvis was reddish in all cases. The urinary bladder was filled with dark red urine in all animals except one animal, in which the urine was grossly normal. Petechiae were present on the mucosa of the small intestine. In all animals, the lumen of the abomasum and/or the large intestine contained large quantities of gelatinous and bright red or tarry blood clots. The spleen was turgid and enlarged and had a gelatinous, dark purple pulp. Subepicardial, subendocardial, and subpleural petechiae and ecchymoses were observed in all animals, in addition to severe pulmonary congestion and edema.
Microscopic Pathology
The scoring of the liver lesions is presented in Supplemental Table 1. The most prevalent microscopic lesion, observed in all 20 animals examined, consisted of large, randomly distributed areas of locally extensive or occasionally multifocal hepatic coagulative necrosis. The necrotic tissue was surrounded by a narrow rim of inflammatory cells composed mostly of viable and degenerated neutrophils and fewer lymphocytes, plasma cells, and macrophages (Fig. 5). Necrosis usually extended to the overlying capsule and occasionally beyond the boundaries of the rim of leukocyte infiltration. The sinusoids in the peripheral rim of inflammation were engorged mainly with degenerated neutrophils (Fig. 5). Larger blood vessels, including arteries and veins, were necrotic and some contained fibrin thrombi that stained positively with PTAH (Fig. 6). Consistently, moderate to severe hepatic centrilobular necrosis was present in areas beyond the main necrotic focus. There was only mild hepatocyte vacuolation, interpreted as hepatic lipidosis. Variable numbers of large Gram-positive rods, many of which contained subterminal spores that deformed the cell, were present singly or in clusters throughout the necrotic foci, including the lumen of thrombosed blood vessels. These rods were more prominent on the inner side of the inflammatory rim at the periphery of the necrotic foci (Fig. 7). In one animal, there was a mild to moderate degree of bile duct proliferation, fibrosis, and infiltration of neutrophils, lymphocytes, and plasma cells, affecting the portal areas outside the focus of necrosis.
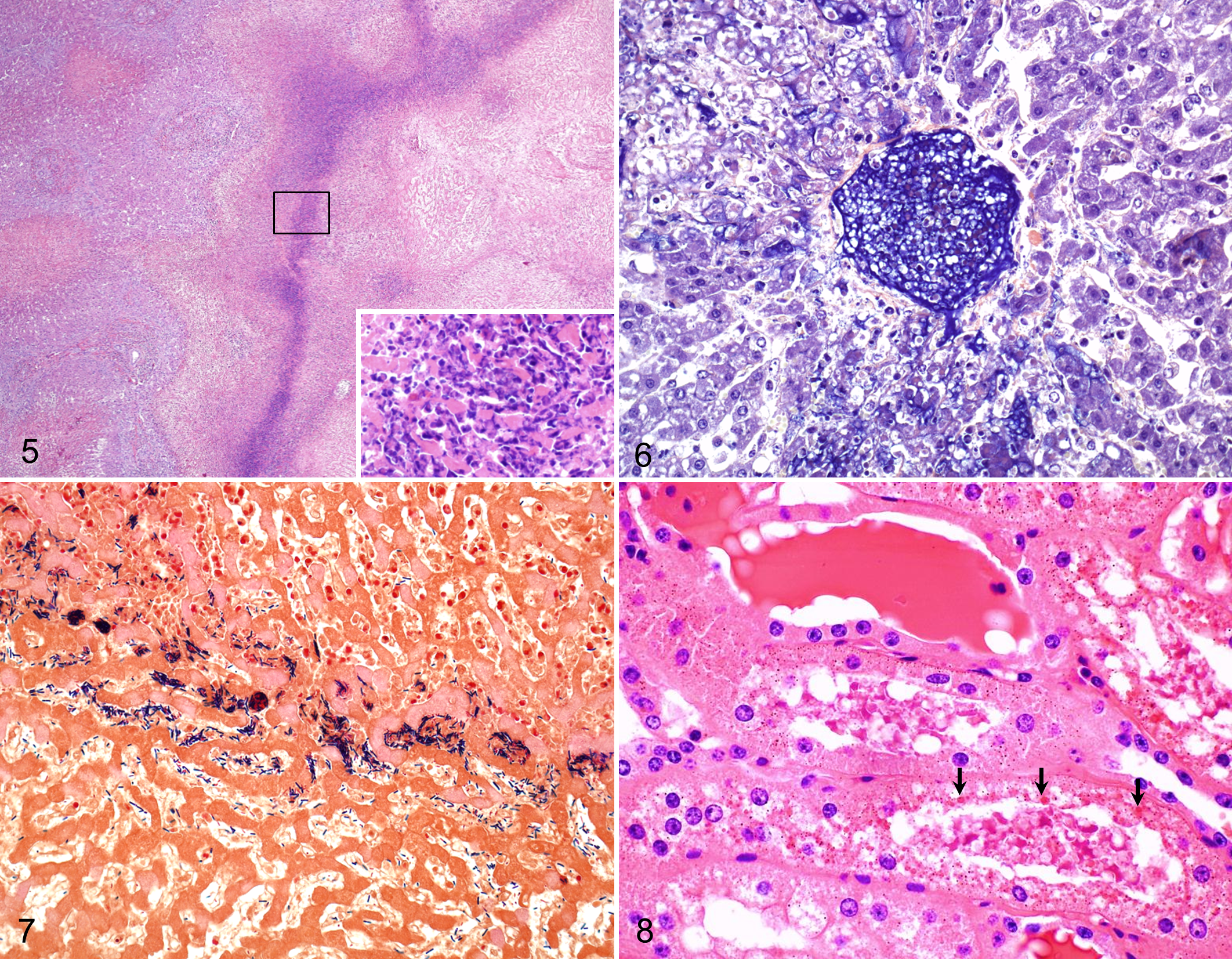
Microscopic examination of the kidneys revealed similar changes in the 11 animals examined. These changes consisted of acute tubular necrosis affecting both proximal and distal convoluted tubules, characterized by degeneration and coagulation necrosis of tubular epithelium, which had varying degrees of attenuation, vacuolation, loss of cellular detail, pyknotic and/or karyorrhectic nuclei, and detachment from the basement membrane. Eosinophilic intracytoplasmic granules were common in epithelial cells of convoluted tubules, and the lumen of these tubules and glomeruli contained protein casts (Fig. 8). The cytoplasmic granules in the tubular epithelium and the luminal protein casts stained orange-brown with Okajima stain for hemoglobin (Fig. 9). One animal also had mild to moderate, multifocal, interstitial infiltration of lymphocytes and fewer plasma cells in both the cortex and the medulla.
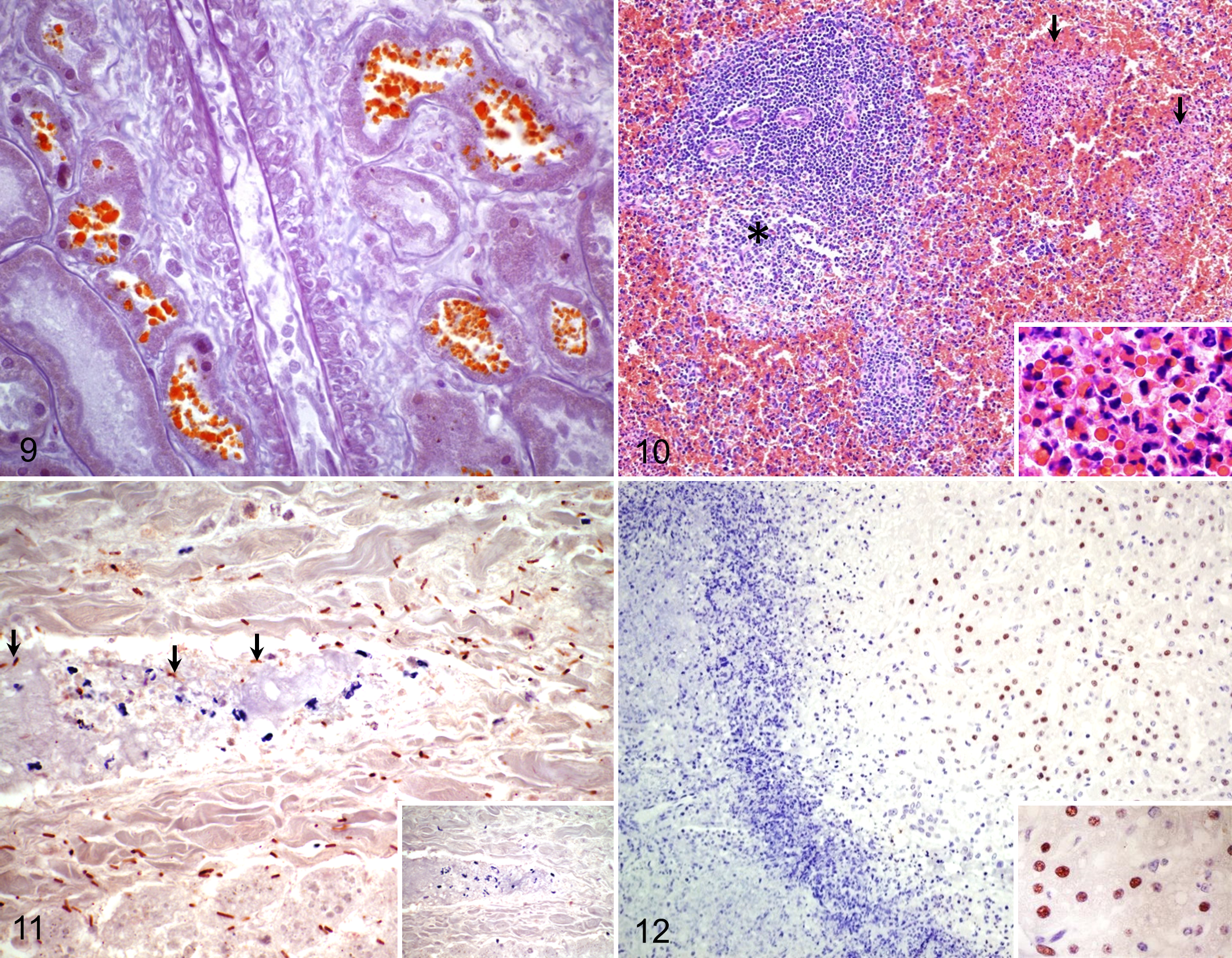
In the spleen of the 3 animals from which this organ was available for examination, sinusoidal congestion and multifocal areas of degeneration, necrosis, and infiltration with degenerate and viable neutrophils were observed in the central areas of the white pulp and throughout the red pulp (Fig. 10).
Microscopic examination of the other organs revealed diffuse, proteinaceous alveolar edema in the lungs of all animals and multifocal subendocardial and subepicardial hemorrhages in all animals. No significant histological abnormalities were observed in other organs.
Immunohistochemistry
Most of the intralesional rods stained positive for C. novyi by IHC in all the liver sections tested (Fig. 11). Occasionally, positive immunostaining was also detected in the cytoplasm of macrophages in the form of irregular granules. Immunostaining for Leptospira spp was negative in sections of kidneys of all 5 animals tested, including the animal with interstitial lymphocytic infiltration. Hepatocytes located outside the necrotic area, which appeared unremarkable on HE-stained sections, exhibited strong positive nuclear immunostaining for activated caspase 3 in all 8 sections tested (Fig. 12), whereas those in the foci of necrosis showed weak or no reaction.
Transmission Electron Microscopy
Ultrastructural examination of areas of hepatic necrosis revealed dissociation of hepatocytes and decrease in cell and nuclear volume. Nuclear and plasma membranes were undulant, and the cytoplasm frequently contained lipids, vacuoles, and debris (Fig. 13). Loss of nucleolus, chromatin, and rough endoplasmic reticulum was also common. Similar changes were observed in endothelial cells. Many erythrocytes showed signs of lysis, appearing as cellular ghosts of variable size, and precipitates compatible with free hemoglobin were observed in the lumen of blood vessels including the hepatic sinusoids (Fig. 14). Clumps of vegetative and sporulating bacilli (Fig. 15) were present throughout the necrotic tissue and within blood vessels. On average, vegetative bacilli were 0.93 ± 0.09 μm wide and 7.11 ± 2.02 μm long (mean ± SD), with a plasma membrane (∼11.3 nm thick), a cell wall (∼26.5 nm thick), and a cell-surface layer (∼10.2 nm thick) (Fig. 16). Spores were 0.85 ± 0.06 μm wide and 1.93 ± 0.05 μm long (mean ± SD).
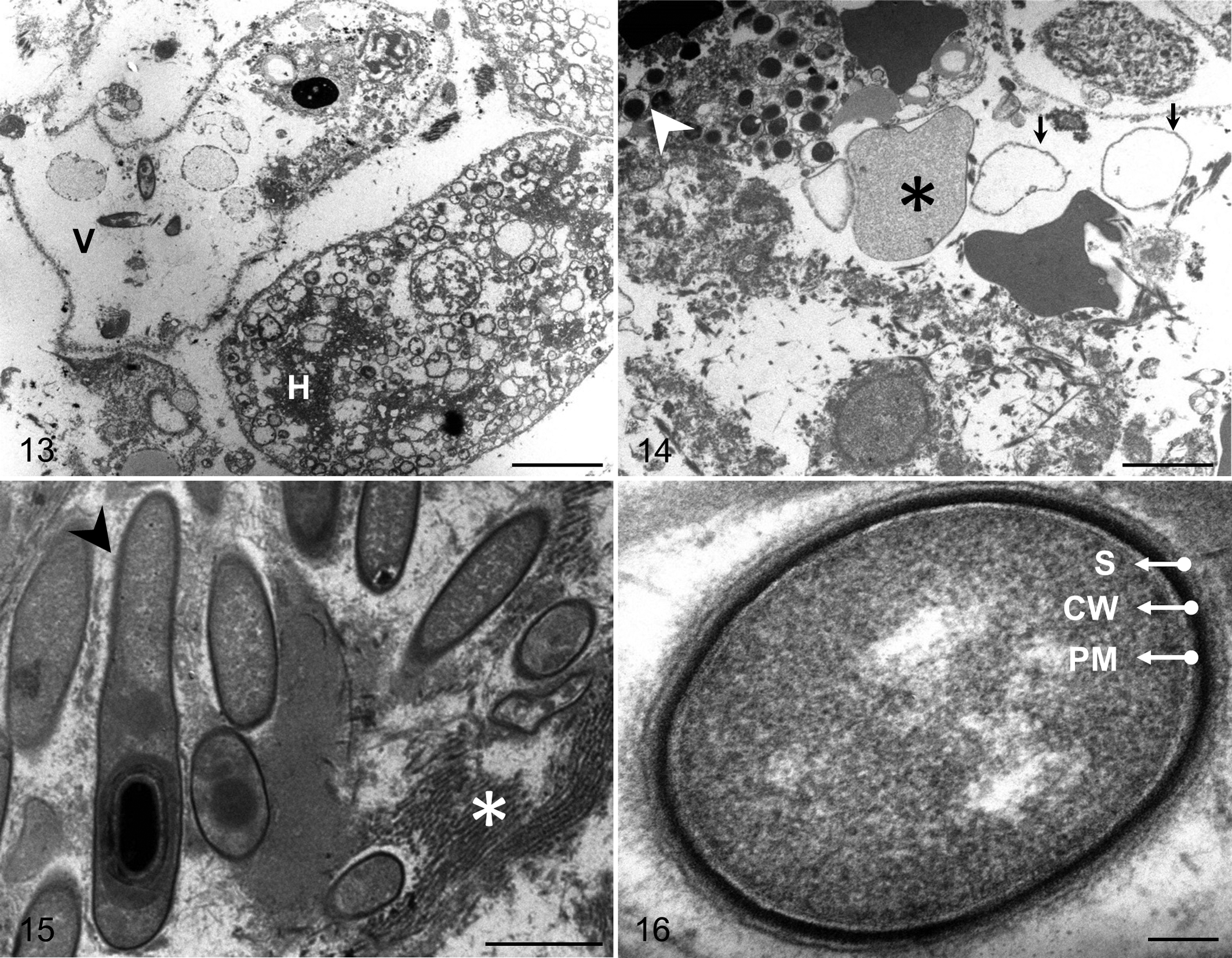
Bacillary hemoglobinuria, liver, bovine No. 17. Transmission electron microscopy.
Polymerase Chain Reaction
PCR of DNA extracted from all 6 randomly selected FFPE liver samples and from the C. haemolyticum strain used as positive control resulted in ∼186 bp amplicons, using primers designed to amplify the C. haemolyticum beta toxin gene (Supplemental Fig. 1). The PCR results were negative when DNA from a normal cow was used as template or when DNA or primers were replaced by water.
Discussion
The aim of this study was to describe the gross, microscopic, and ultrastructural lesions and to study the mechanism of cell death in the liver of a series of bovine cases with confirmed spontaneous BH. A presumptive diagnosis of BH is based on clinical, gross, and microscopic findings, with a more certain diagnosis being supported by FAT and/or immunohistochemistry. The last 2 tests add certainty to the diagnosis, although they do not provide final evidence of the role of C. haemolyticum, as they are usually performed, as in this study, with antibodies to the related bacterium C. novyi. 5,23,32 Confirmation of a diagnosis of BH in animals with clinical signs and lesions compatible with this disease is achieved by demonstration of C. haemolyticum in the liver of affected animals by culture and/or PCR. 10,19
In this study, the presumptive diagnosis was based on clinical and pathological findings, including IHC, and was confirmed in a representative number of animals by PCR testing. No anaerobic culture facilities were available in the area at the time of necropsy, and culture was therefore not attempted. In addition, C. haemolyticum is considered the strictest anaerobe of the pathogenic clostridia, 19 and the success rate when isolation is attempted is low; thus, PCR confirmation represented a valuable diagnostic tool in this series of cases.
Although little is known about the pathogenesis of BH, it is presumed that once the spores of C. haemolyticum are ingested, they arrive to the liver via the portal circulation and are phagocytized by resident macrophages (Kupffer cells), where the spores may remain latent in the cytoplasm for long periods. 19,26 An anaerobic environment in the liver is required to allow the germination of the phagocytized spores. When this happens, the vegetative forms of C. haemolyticum produce the highly necrotizing and hemolytic beta toxin that hydrolyzes phosphatidylcholine into phosphocholine and a diglyceride, probably causing the disruption of the plasma membrane of Kupffer cells. Once released, beta toxin leads to hepatocyte, erythrocyte, and endothelial cell disruption (the latter inducing thrombus formation). The progression of this initial lesion is likely responsible for the large necrotic focus observed in all cases of BH. Necrosis is, in turn, likely to be exacerbated by the anaerobiosis of the liver parenchyma caused by the hemolysis and thrombosis, thus allowing bacterial survival and the production of larger quantities of beta toxin in a vicious cycle. 10,11,19
In this series of cases, the most consistent gross and microscopic lesion was found in the liver, and it was characterized by locally extensive or multifocal hepatic necrosis, in which thrombosis was a consistent finding. The liver lesion characteristic of BH has been traditionally described as an infarct associated with thrombosis. 29 Even though thrombi were seen in all animals of this study, it is unlikely that they were solely responsible for the extensive damage observed in the liver parenchyma. The double circulation of the liver provided by branches of the hepatic artery and portal vein makes the occurrence of hepatic infarction relatively unusual. 9 A more plausible explanation is that beta toxin initially produces at least some parenchymal and endothelial cell necrosis, with the latter predisposing to thrombosis; this may, in turn, be responsible for ischemia and associated necrosis, perpetuating the anaerobiosis. The latter would provide an ideal environment for continuous proliferation of C. haemolyticum with additional toxin production and a repetitive cycle of tissue damage.
In this study, we used IHC for activated caspase 3 to characterize the mechanism of cell death in the liver of animals with BH. While most cells within the necrotic foci were negative, many hepatocytes beyond the inflammatory cell rim were positively labeled for activated caspase 3, indicating apoptosis as the main mechanism of cell death occurring in the periphery of the lesions and suggesting that the mechanism of cell death in the center of the main lesion was in fact necrosis. It could be speculated that within these areas of bacterial infection, high concentrations of toxin cause necrosis, while lower concentrations of toxin peripheral to these areas may induce apoptosis. The same has been demonstrated for other clostridial toxins, such as Clostridium perfringens enterotoxin, which causes apoptosis of cells when present in low concentration and necrosis when present in high concentration in vitro. 3,4
Apoptosis may also be associated with factors other than clostridial toxins, or it may be the result of a combination of several events. For instance, recruitment and activation of inflammatory cells lead to cytokine production, including tumor necrosis factor α (TNF-α) and TNF-related apoptosis-inducing ligand (TRAIL), resulting in ligation with Fas receptor on hepatocytes and further activation of caspase 8; 15,36,37 a similar event has been proposed with the increase of toxic bile acids during cholestasis, 27 a phenomenon likely to occur in cases of BH. This inflammatory mechanism of apoptosis could be exacerbated by the production of danger associated molecular patterns (DAMPs) from dying hepatocytes. 15 Additionally, the ischemic and hypoxic state of the liver during BH, mentioned above, may contribute to the establishment of apoptosis. 18,24 Further studies are needed to determine whether apoptosis in cases of BH occurs via extrinsic or intrinsic pathways and also to confirm the mechanism of cell death in the main hepatic lesion.
Migrating liver trematodes, mainly F. hepatica, have been indicated as the main predisposing factor for BH, and the treatment and control of fascioliasis usually result in the decline of the disease in areas where this parasitosis is endemic. 29 A significant finding in this study was the absence of hepatic trematodes and the absence of gross and microscopic lesions consistent with F. hepatica infestation in all animals with BH. The liver of only one animal in our study had moderate changes in portal areas, including fibrosis, mononuclear inflammation, and bile ductal hyperplasia. While these changes may suggest a previous infestation with liver flukes, 16 they are nonspecific and could have been the response to a variety of causes, including, but not limited to, the action of pyrrolizidine alkaloids 6 and prolonged intrahepatic cholestasis followed by leakage of bile. 14 Furthermore, none of the animals in this study originated in areas where the environmental conditions allow completion of the life cycle of F. hepatica.
In addition to liver flukes, other less frequent triggering events for BH have been described, including hepatic necrosis due to Fusobacterium necrophorum infection secondary to rumenitis, 12 liver damage associated with Cysticercus tenuicollis, 28,29 sampling for liver biopsies, 21 incidental vascular disorders such as telangiectasia, 29 and other causes of reduced liver perfusion. Among the latter, reduced liver perfusion during late pregnancy or due to rumen filling by indigestible roughage has been anecdotally reported, although no scientific evidence is available to support this claim. It has been also suggested, albeit never proved, that obesity or hepatic lipidosis may increase susceptibility to BH. Since no evidence of these potential predisposing factors was found in the analyzed animals, the inciting hypoxic events leading to BH in the present study remain undetermined. Further studies and observations are needed to determine the predisposing factors for BH in cattle free of fascioliasis.
The majority of the BH-affected farms in this study were situated on the eastern plains of Uruguay, mostly rice-farming areas characterized by low, poorly drained soils. As C. haemolyticum can survive for years in wetlands or poorly drained soils, flooding and any land disruptions (such as vegetation clearing, soil breaking, or drainage modifications, common in rice production areas) could unearth spores, thus favoring ingestion by cattle, infection of the liver with spores, and resulting in outbreaks of the disease. Multiple liver infarcts in some of the cases presented here, and a higher prevalence of lesions in the right lobe of the liver (which receives the portal blood supply mostly from the anterior digestive tract 7 ), favor the hypothesis that these cases developed as a consequence of spore overload, rather than activation of spores that may have been already present in the liver.
As the disease progresses, C. haemolyticum beta toxin concentration increases in the blood, leading to systemic consequences, including hemolytic anemia, hemoglobinuria, endothelial damage, and hemorrhages. 10,31 Hepatic centrilobular necrosis and renal tubular necrosis observed in this study are most likely consequences of anemia-associated hypoxia, which is clinically manifested with severe dyspnea prior to death. The effect of hemoglobin as nephrotoxic is controversial since intravenous infusions of hemoglobin in healthy animals produce no distinct lesions, but it can induce additive negative effects on tubular epithelium undergoing ischemic necrosis. 20
Multifocal necrosis with neutrophilic infiltration of both red and white pulp of the spleen was seen in all 3 samples of spleen available for this study. Although this was previously described in experimentally infected animals, 21 no descriptions of these lesions in spontaneous cases of BH have been published. The pathogenesis of this lesion is not known but may be the result of lymphocyte cell death 8,30 induced by one or more of the C. haemolyticum toxins.
In summary, the cases of BH presented in this study occurred in F. hepatica–free animals without a seasonal predominance. The gross and microscopic hallmark lesions were locally extensive coagulative necrosis with an adjacent rim of necrosuppurative hepatitis and thrombosis, as well as jaundice, hemoglobinuria, and acute renal tubular necrosis. Apoptosis seems to play a significant role as a mechanism of cell death in areas of liver adjacent to the necrotic foci. Postmortem examination and histopathology coupled with detection of C. novyi by IHC and/or FAT, and of C. haemolyticum by PCR, are recommended for diagnosis. Even though migrating liver trematodes have been considered the main triggering factor for BH, there are other yet undetermined predisposing factors for this disease.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.