
Abstract
Literature data indicate heat shock protein (Hsp) 32 and 90 as potential molecular targets in canine neoplastic mast cells (MCs). However, their immunoexpression patterns in canine mast cell tumors (MCTs) have not been investigated. Thus, the aim of this study was to evaluate the immunohistochemical expression of Hsp32 and Hsp90 in 22 canine cutaneous MCTs, in relation to KIT immunolabeling pattern, histological grade, and mitotic count. All cases showed cytoplasmic labeling of Hsp90, variably associated with nuclear and/or membranous labeling. Relationships of Hsp90 or Hsp32 immunolabeling with KIT pattern, mitotic count, and tumor grade were not observed. However, the reduced Hsp32 immunoexpression observed in most grade III/high-grade MCTs suggests a tendency toward a loss of immunosignal in poorly differentiated MCs. The great heterogeneity in extent and distribution of Hsp90 immunoexpression among the different MCT cases may also partially explain the difficulties in predicting the in vivo biologic activity of Hsp90 inhibitors on canine MCTs.
Cutaneous mast cell tumors (MCTs) are among the most common tumors in dogs, accounting for 16%–21% of all skin neoplasms. As KIT-driven tumors, canine cutaneous MCTs are considered a unique model to study several KIT-dependent human malignancies (supplemental reference 3). Overexpression of heat shock protein 90 (Hsp90), a molecular chaperone for which KIT is a client protein, is predictive of adverse behavior in human gastrointestinal stromal tumors. 3 The inhibition of its function may also represent a promising therapeutic solution for KIT-driven diseases. In this respect, Hsp90 inhibitors have been demonstrated to be effective in down-regulating mutated, constitutively activated KIT protein in human mast cells (MCs). 1 At the same time, these inhibitors exhibit activity against KIT-dependent and -independent canine malignant MCTs. 6
Hsp32, also known as heme oxygenase-1 (HO-1), has been identified as an important KIT-inducible survival factor and a potential therapeutic target in neoplastic human MCs. 5 As well, targeting of Hsp32 by pharmacologic inhibitors is associated with reduced growth and induction of apoptosis of neoplastic canine MCs. 2
Notwithstanding this, the Hsp immunoexpression pattern in canine MCTs has not been investigated so far. Thus, the aim of the present study was to evaluate the immunohistochemical expression of Hsp32 and Hsp90 in canine cutaneous MCTs, in relation to KIT labeling pattern, histological grade, and mitotic count.
The study was carried out using formalin-fixed, paraffin-embedded samples obtained from 22 cases of treatment-naïve canine cutaneous MCTs. Histologic diagnosis was performed on serial slides for each tumor sample stained with hematoxylin and eosin and toluidine blue (Sigma-Aldrich, St Louis, MO, USA). The neoplasms were classified according to Patnaik et al 9 and Kiupel et al, 4 as 13 grade II (10 low-grade and 3 high-grade) MCTs and 9 grade III (all high-grade) MCTs (Table 1). Mitotic count was calculated as the number of mitotic figures counted in a standard area of 2.37 mm2 (using 40× objective and 10× ocular with field number 22 mm; Table 1), as recently proposed (supplemental reference 2).
Summary of grading, mitotic count, KIT labeling pattern, and HSP immunosignal of canine cutaneous MCTs.a
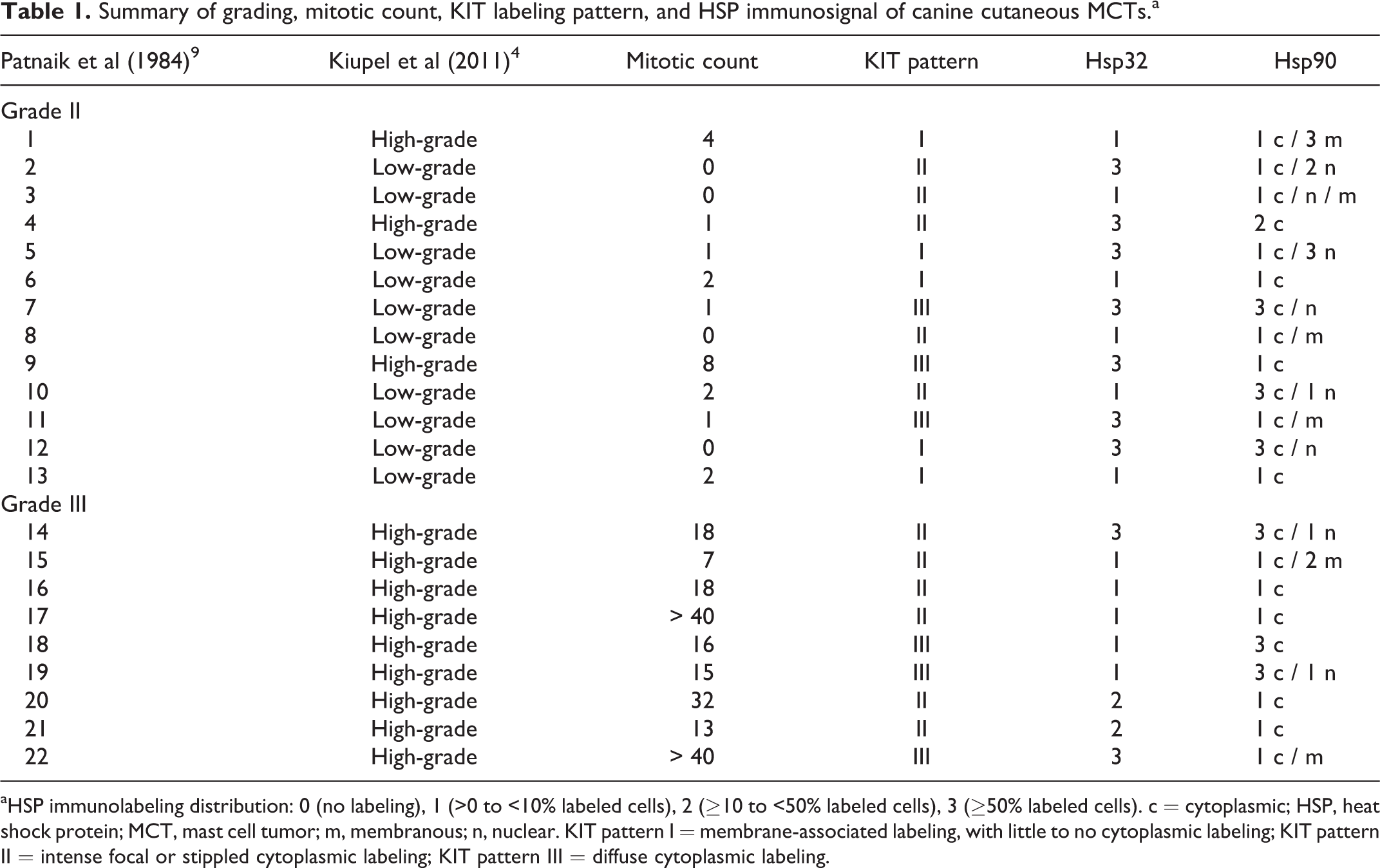
aHSP immunolabeling distribution: 0 (no labeling), 1 (>0 to <10% labeled cells), 2 (≥10 to <50% labeled cells), 3 (≥50% labeled cells). c = cytoplasmic; HSP, heat shock protein; MCT, mast cell tumor; m, membranous; n, nuclear. KIT pattern I = membrane-associated labeling, with little to no cytoplasmic labeling; KIT pattern II = intense focal or stippled cytoplasmic labeling; KIT pattern III = diffuse cytoplasmic labeling.
Immunohistochemistry was performed using primary antibodies specific for Hsp32 (1:400 dilution, polyclonal rabbit anti-rat; StressGen/Assay Designs, Inc), Hsp90α/β (1:1.800 dilution, monoclonal mouse anti-water mold, clone AC88, StressGen/Assay Designs, Inc), KIT (1:800, polyclonal rabbit anti-human; Dako, Glostrup, Denmark), according to a previously described technique (supplemental reference 4). Deparaffinized and rehydrated sections were incubated with 3% H2O2 in absolute methanol for 45 minutes to inhibit endogenous peroxidase activity and then rinsed in 0.05 M Tris-buffered saline (TBS; pH 7.6) for 5 minutes. Antigen retrieval was performed by heat treatment in citrate buffer, pH 6.0, in a microwave oven for 5 minutes (3 cycles). To reduce nonspecific binding, slides were then incubated with normal goat serum (Vector Laboratories Inc, Burlingame, CA, USA) for 15 minutes at room temperature before overnight incubation with the primary antibody in a humidified chamber at 4°C. After rinsing with TBS, sections were incubated at room temperature for 30 minutes with secondary biotinylated goat anti-mouse or anti-rabbit antibodies (1:200 dilution; Vector Laboratories). Labeling was subsequently detected using an avidin-biotin complex (ABC) method (Vectastain ABC Kit, Vector Laboratories) with 0.1% H2O2 in 3,3′ diaminobenzidine solution (Sigma-Aldrich) as chromogen. Sections were counterstained with Mayer’s hematoxylin (Merck, Darmstadt, Germany) for 1 minute before rinsing, dehydrating, and mounting. A negative control was performed in all instances by omitting the primary antibody and replacing it by an antibody of irrelevant specificity (polyclonal rabbit anti-human von Willebrand factor or monoclonal mouse anti-human desmin antibodies, both from DAKO, Glostrup, Denmark). Sections of bovine urinary bladder and canine nerve known to display HSP expression (supplemental references 4–5) were used as positive controls.
HSP immunohistochemical labeling of the cytoplasm, nucleus, and/or membrane was assessed semiquantitatively as 0 (no labeling), 1 (>0 to <10% labeled cells), 2 (≥10 to <50% labeled cells), 3 (≥50% labeled cells). The KIT immunolabeling pattern was classified according to Kiupel et al, 4 as follows: KIT-labeling pattern I (membrane-associated labeling, with little to no cytoplasmic labeling), KIT-labeling pattern II (intense focal or stippled cytoplasmic labeling), KIT-labeling pattern III (diffuse cytoplasmic labeling).
Fisher exact test was used to compare HSP expression scores with KIT pattern, histologic grade, and mitotic count. Cramer’s V was also calculated to measure the strength of association between the variables. Analyses were performed using SPSS statistical software, with P < .05 considered to be significant.
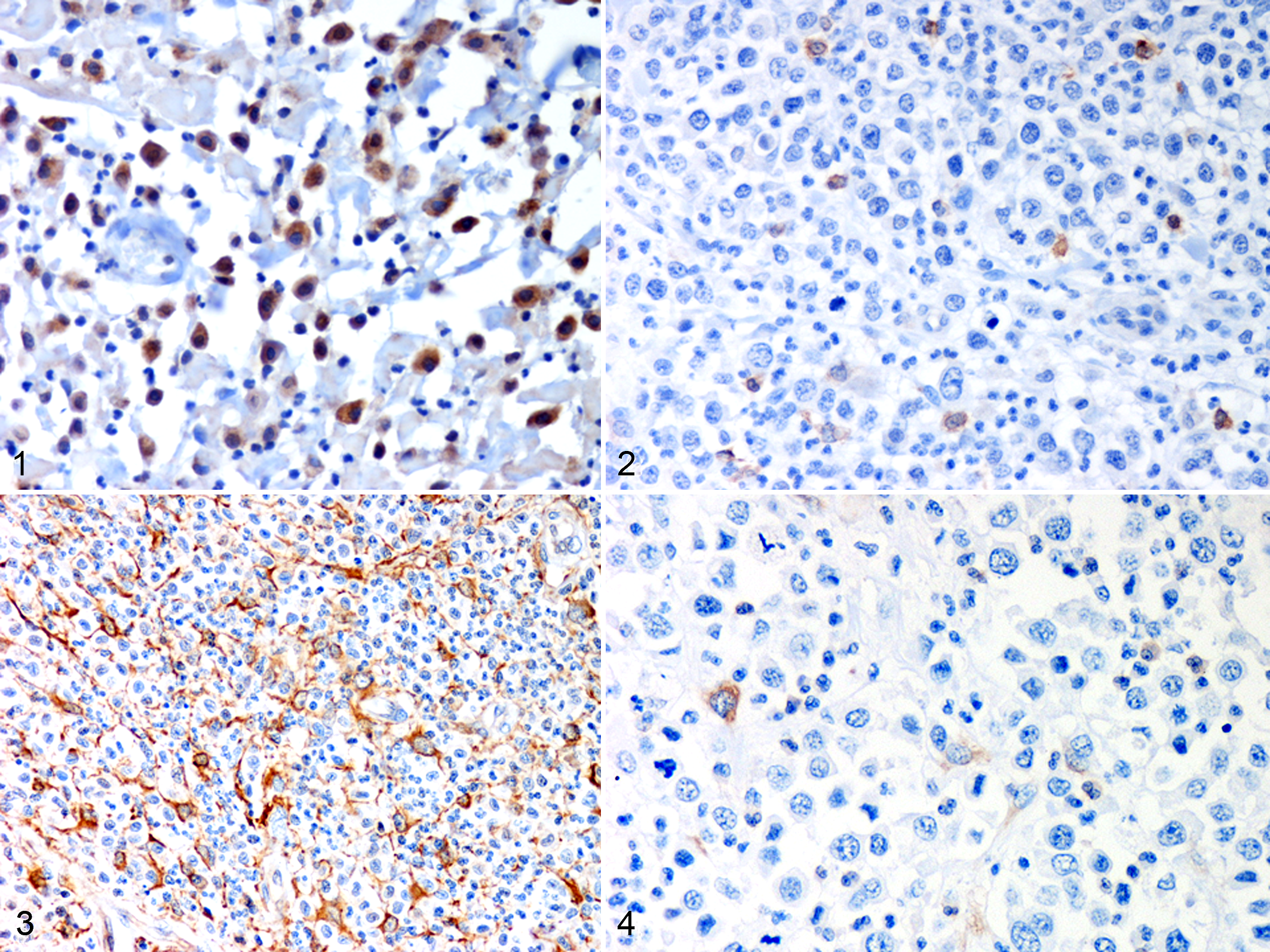
Hsp32 immunolabeling was variably intense, cytoplasmic, and generally strong with a granular appearance (Fig. 1). The expression pattern of this HSP was usually characterized by the presence of highly positive tumor cells admixed with a population of less intensely labeled tumor cells (Fig. 2). Hsp90 labeling was variably distributed, moderate to strong, and predominantly cytoplasmic but also nuclear and/or more rarely membranous (Figs. 3–4).
Although an association with the histologic grade and mitotic count was not detected for either of the HSPs, a reduced percentage of Hsp32 immunoexpression (<50% positive cells) was observed in most grade III (7/9)/high-grade (8/12) MCT samples (Table 1). With references to KIT expression status (Table 1), all 3 patterns of immunolabeling were observed. However, KIT labeling pattern I was the most rarely detected and was not observed in grade III MCTs. On the other hand, KIT-labeling pattern II was predominantly detected in grade III MCTs. Statistical analysis did not reveal significant associations with HSP immunolabeling.
The present study demonstrated the immunohistochemical expression of Hsp32 and Hsp90 in canine cutaneous MCTs. Hsp32 is considered a potential therapeutic target to be tested in clinical trials of canine patients with aggressive MCTs. 2 However, primary neoplastic MCs expressing Hsp32 were isolated from surgical samples of grade I and grade II, but not of grade III, canine MCTs, 2 so that Hsp32 expression in poorly differentiated MCs was not known. In this respect, the limited proportion of Hsp32 immunoexpression observed in most grade III/high-grade MCT samples included in the present study indicates a tendency toward a loss of Hsp32 expression during neoplastic MC dedifferentiation. Therefore, further investigations on the efficacy of the pharmacologic Hsp32 inhibitors on neoplastic MCs isolated from aggressive canine MCTs are needed. Even though an association between Hsp32 and KIT immunolabeling was not detected, this could be due to the multiple signaling pathways contributing to Hsp32 expression in neoplastic canine MCs. 2 In addition, it is known that Hsp32 induction is not limited to KIT mutations in MCTs, since stem cell factor (SCF)–activated wild-type KIT may also promote Hsp32 expression in neoplastic MCs. 5 The lack of association between Hsp90 immunoexpression and the MCT histologic grade further supports the hypothesis that up-regulation of Hsp90 expression is not a common event during neoplastic transformation and malignant progression of canine MCs. 6 The absence of association between Hsp90 and KIT immunoexpression pattern is also in line with the broad activity demonstrated by the Hsp90 inhibitors against MCs expressing wild-type or mutant KIT. 6 This finding probably reflects the multiple chaperone activity of Hsp90 directed toward a wide variety of client oncoproteins. However, the high variability of extent and distribution on tumor tissue, characterizing the Hsp90 immunosignal detectable among the different MCT cases, could be one of the reasons for the partial response of canine MCTs to treatments based on Hsp90 inhibition. 7 In fact, the in vivo biologic activity of Hsp90 inhibitors may not be as significant as predicted by experimental models, since responses may be dependent on addiction of the tumors to Hsp90 and its client oncoproteins (supplemental reference 1).
Finally, despite the lack of association between Hsp90 expression and grade, KIT pattern, or mitotic count, further studies are necessary to clarify the possible prognostic significance of the different intracellular localizations (cytoplasmic, nuclear, membranous) of the Hsp90 immunoexpression detected in canine MCTs. Indeed, such localizations in different subcellular compartments could be related to the different alpha and beta Hsp90 isoforms, both recognized by the antibody used in the present study, which may play different roles in carcinogenesis and tumor progression. 8 Particularly, Hsp90 alpha has been identified on the surface of cancer cells and is implicated in tumor migration, invasion, and metastasis. 10
In conclusion, this investigation demonstrated, for the first time, the immunohistochemical expression pattern of Hsp32 and Hsp90 in canine MCTs and revealed a tendency toward a loss of Hsp32 in poorly differentiated MCs. The great heterogeneity in extent and distribution of Hsp90 expression among the different MCTs may partially explain the difficulties in predicting the in vivo biologic activity of Hsp90 inhibitors on canine MCTs.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.