
Abstract
Swine dysentery is a severe enteric disease in pigs, which is characterized by bloody to mucoid diarrhea and associated with reduced growth performance and variable mortality. This disease is most often observed in grower–finisher pigs, wherein susceptible pigs develop a significant mucohemorrhagic typhlocolitis following infection with strongly hemolytic spirochetes of the genus Brachyspira. While swine dysentery is endemic in many parts of the world, the disease had essentially disappeared in much of the United States by the mid-1990s as a result of industry consolidation and effective treatment, control, and elimination methods. However, since 2007, there has been a reported increase in laboratory diagnosis of swine dysentery in parts of North America along with the detection of novel pathogenic Brachyspira spp worldwide. Accordingly, there has been a renewed interest in swine dysentery and Brachyspira spp infections in pigs, particularly in areas where the disease was previously eliminated. This review provides an overview of knowledge on the etiology, pathogenesis, and diagnosis of swine dysentery, with insights into risk factors and control.
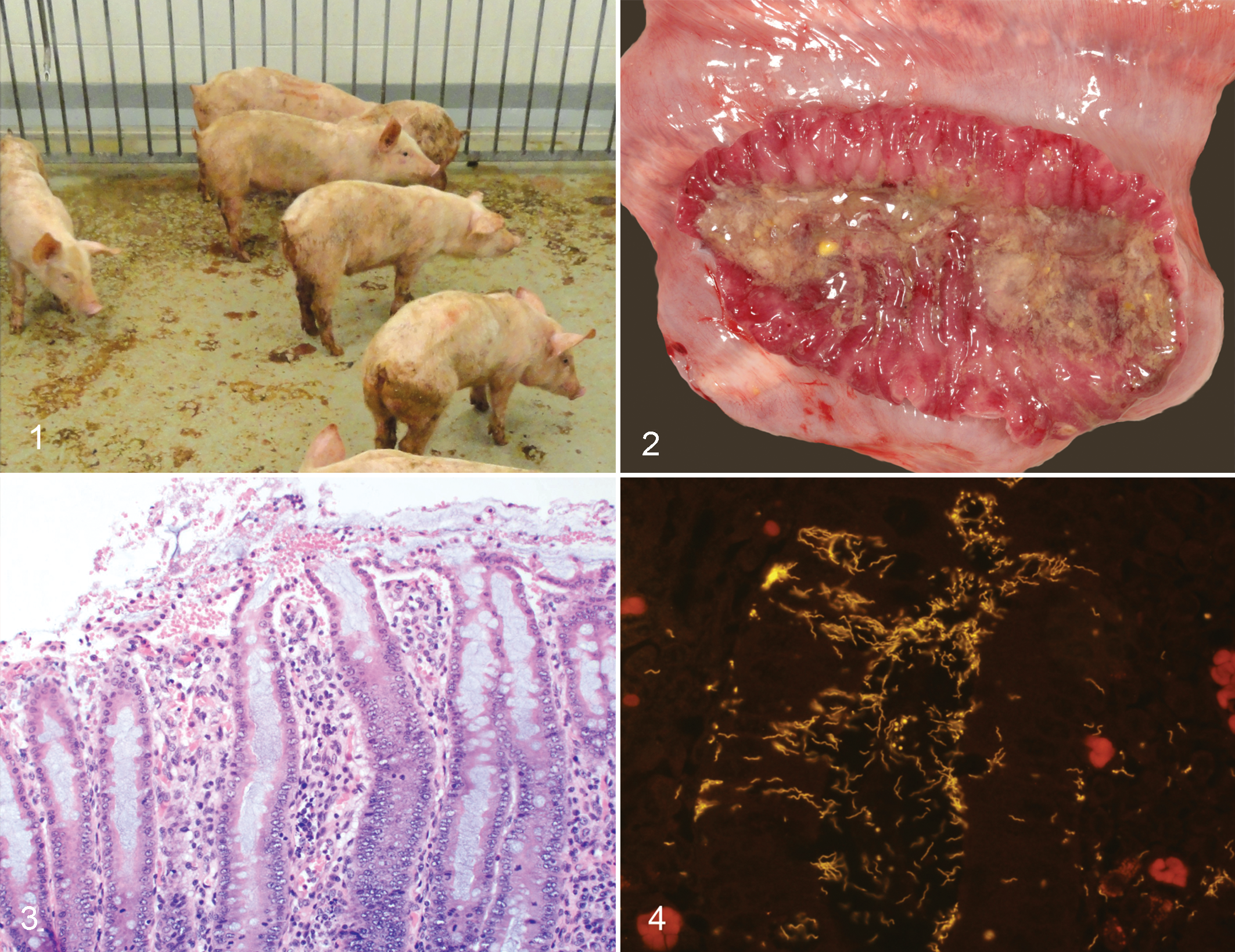
Brachyspira spp infections in pigs have recently gained renewed interest in the US swine industry due to a period of reemergence throughout North America. 12 Of particular concern has been an increase in the detection of strongly β-hemolytic Brachyspira spp from pigs with clinical swine dysentery (SD), given that the disease had been eliminated from most US production systems since the early 1990s. 42,91 Swine dysentery is most often observed in growing and finishing pigs (approximately 8–26 weeks of age) and is characterized by profuse mucohemorrhagic diarrhea and loss of body condition (Fig. 1). 42 Field outbreaks were first described in the literature in 1921, with disease occurring in feeder hogs 5 days to 2 weeks after arrival on the farm. 133 In naive weaned pigs, morbidity and mortality can approach 90% and 30%, respectively. 42 Accordingly, the financial impact on affected farms can be severe due to a combination of poor feed conversion, medication costs, and mortality. Swine dysentery occurs worldwide, yet there is very limited information available regarding the prevalence of SD in different countries. 42 Sporadic reports of detection rates in field diagnostic and surveillance samples provide at least an indicator of disease incidence, and recent reports have been published regarding detection in multiple countries in North America, Europe, and Asia. 8,17,21,37,45,49,117 Since 2007, there has been an increase in SD diagnoses at North American veterinary diagnostic laboratories, 12 yet the precise factors underlying this reemergence remain unclear.
Etiology
Clinical SD was successfully reproduced experimentally in the 1920s through the feeding of intestinal contents from diseased pigs, 133 yet the precise etiologic agent was not elucidated for many years. In the initial pathologic description, 133 lesions were associated with curved bacterial rods, delicate spirochetes, and ciliated protozoa, and Campylobacter (Vibrio) coli was proposed as the causative agent 28,83 ; however, other groups failed to experimentally reproduce SD following inoculation with pure cultures of C. coli in both gnotobiotic and conventional pigs. 2,3 In the 1970s, 2 separate groups reported the reproduction of SD following oral inoculation of pigs with cultures of spirochetes. 48,123 This novel spirochete was originally named Treponema hyodysenteriae 48 and was then briefly reclassified as Serpula 121 and Serpulina 119 before its most recent reclassification in the genus Brachyspira. 94 Brachyspira hyodysenteriae is a gram-negative, oxygen-tolerant, anaerobic spirochete measuring approximately 8–10 μm in length and 0.3–0.4 μm in diameter with 14–18 periplasmic flagella per cell. 94 Early experimental reproduction of SD was shown to be limited to pigs concurrently infected with C. coli and B. hyodysenteriae and a role for both agents as copathogens was proposed 35 ; however, this association was later refuted when multiple attempts to reproduce the disease in gnotobiotic pigs following infection with a combination of these 2 agents were unsuccessful. 11,86 Campylobacter spp infections are very common in pigs with Brachyspira-associated colitis, 17 which may suggest a potential relationship between these bacteria in the pig gut. Interestingly, multiple groups reported a lack of clinical disease in gnotobiotic pigs following inoculation with B. hyodysenteriae alone, 47,86 while clinical disease did occur in gnotobiotes inoculated with colonic scrapings from pigs with clinical disease 87 or combinations of B. hyodysenteriae and one or more gram-negative anaerobes presumed to comprise part of the normal porcine intestinal microbiota. 47,88,132 Taken together, these findings suggest that SD can only occur when susceptible pigs, where susceptibility is at least in part driven by their intestinal microbiota, are exposed to a virulent Brachyspira spp.
Coincident with the recent reemergence of SD in US swine has been the identification of multiple novel Brachyspira spp recovered from pigs with mucohemorrhagic diarrhea including Brachyspira suanatina in Europe 93,103 and the proposed “Brachyspira hampsonii” in North America. 22 Initial genetic analysis of “B. hampsonii” isolates revealed 2 distinct clades (clade I and clade II) 22 ; however, this genetic distinction does not appear to be clinically significant, as disease consistent with classic SD has been experimentally reproduced following inoculation with strains from each clade. 15,16,26,109 Although “B. hampsonii” was first reported in the USA and Canada, it has been subsequently detected in pigs and waterfowl in multiple European countries 84,85,106 and in geese in the Canadian Arctic. 110 Recent genetic analysis of global “B. hampsonii” isolates using multilocus sequence typing (MLST) revealed 4 distinct genetic groups (genetic groups I–IV), which supplants the previous clade designations (the previous clade I and clade II isolates belong to genetic groups I and II, respectively) and supports the inclusion of all 4 genetic groups in a single species. 90 B. suanatina has a much more limited geographic range and has thus far not been reported outside Sweden and Denmark. 93,103 The one unifying feature of all 3 Brachyspira spp recently recovered from pigs with SD is strong β-hemolysis when cultivated on blood agar, and this feature has been suggested as an indicator of virulence in clinical isolates from pigs. 16 Indeed, the importance of hemolysins as virulence factors has been emphasized through experimental infections where B. hyodysenteriae mutant strains lacking the hemolysin gene tlyA were associated with reduced lesions in mice and a lack of SD development in pigs. 56,125 Recent publication of the whole genome of B. hyodysenteriae revealed genes for 7 potential hemolysins 7 including 4 that were previously reported (tlyA, tlyB, tlyC, and hlyA). 53,124 The precise role of each individual hemolysin in the pathogenesis of SD is unclear; however, it seems reasonable to conclude that the expression of strong β-hemolysis is an important factor in disease development. A 36-kb plasmid has also been described in virulent strains of B. hyodysenteriae that likely contains genes involved in colonization or disease expression. 79
There are numerous weakly β-hemolytic Brachyspira spp that commonly colonize the colon of pigs including B. innocens, B. intermedia, B. murdochii, and B. pilosicoli. 42 Aside from B. pilosicoli, which is the etiologic agent of porcine intestinal spirochetosis, 128 these are generally considered of low pathogenic potential and are commensal in nature. None of the currently recognized weakly β-hemolytic Brachyspira spp have been associated with bloody diarrhea typical of SD.
Pathogenesis
As previously noted, gnotobiotic pigs are variably colonized by B. hyodysenteriae but do not develop signs of SD, 47,86 whereas gnotobiotes orally inoculated with intestinal scrapings and content from pigs infected with SD readily develop disease. 87 These findings suggest that infection with at least one other microorganism 88 is required for disease expression, and anaerobes such as Bacteroides vulgatus and Fusobacterium necrophorum have been shown to successfully fulfill this requirement. 47,132 Natural transmission of Brachyspira spp occurs primarily via ingestion of infected feces. The spirochetes must survive the acidic environment of the stomach and pass through the small intestine until ultimately reaching, colonizing, and proliferating in the cecum and colon. Although the precise dose required in natural infections is unknown, and experimental models and inoculation protocols vary, an inoculum containing at least 105 colony-forming units (CFUs) is often required for experimental infection. 16,70
Once established in a susceptible pig, B. hyodysenteriae colonizes the mucus layer and is often found deep within crypts likely driven at least in part by its strong chemotaxis to porcine mucins. 68,69,89 B. hyodysenteriae can metabolize substantial amounts of oxygen, 118 and NADH oxidase activity appears important in colonization capacity as NADH-oxidase-negative mutants of B. hyodysenteriae have reduced capacity to colonize pigs and are also associated with milder clinical disease. 122 Colonization within the mucus layer and crypts helps further protect the spirochetes from potential oxygen toxicity at the surface, and flagellar function is important in mucus penetration. Brachyspira spp often exhibit a corkscrew-like motility generated by the periplasmic flagella that helps the organisms penetrate the mucus layer, and the importance of flagellar activity has been demonstrated for B. hyodysenteriae where FlaA and FlaB mutants were associated with reduced colonization in vivo. 67
Once colonized, the spirochetes may appear in the feces of pigs from 1–4 days prior to the observance of clinical signs. 70 In experimental B. hyodysenteriae infections, SD is often first observed in a few pigs approximately 7–10 days postinoculation, with morbidity approaching 90% or greater within 3 weeks. 134,139 In experimental inoculation studies with “B. hampsonii”, clinical disease has been reported in individual pigs as early as 4–5 days postinoculation and morbidity is often less than 90%. 109,135 The spirochete load may also be important in lesion development as experimental studies suggest at least 105 CFU per gram of B. hyodysenteriae in the mucosa is associated with the presence of lesions, while lower loads are asymptomatic and lack lesions at necropsy. 139
In early stages of B. hyodysenteriae infection, there is a reduction in mucigen within the goblet cells at the base of crypts and concurrent dilation of the luminal aspects of crypts with accumulations of expelled mucin. 139 Serial evaluation via endoscopy through a cecal cannula has demonstrated that expulsion of mucin in the deeper crypts typically occurs within the first 3 days of clinical disease and that goblet cell hyperplasia begins after the fifth day. 58 Histochemical analyses have shown that reduced expression of sulfated mucins and expression of sialomucins increases within crypts at the apex of the spiral colon in pigs with acute SD. 134,136 Immunohistochemistry has further revealed that there is a specific reduction in mucin 4 and an increase in mucin 5AC (MUC5AC) in pigs 48 hours after clinical expression of SD associated with either B. hyodysenteriae or “B. hampsonii”. 136 Mucin 2 (MUC2), which is constitutively expressed in the pig colon, is predominantly detected within goblet cells of healthy pigs and is to a large amount expelled into colonic crypts within 72 hours of development of clinical SD, whereas MUC5AC expression is not normally detected in the pig colon and appears within goblet cells over the same time frame in pigs with acute SD. 134 Mucin production is 5-fold higher in experimentally infected pigs with SD relative to controls due to the de novo production of MUC5AC and an increase in MUC2 production. 101 Mucin 5AC production is not upregulated in those pigs that are inoculated but do not develop SD, and MUC5AC expression is significantly correlated with the presence of neutrophilic infiltration. 101,134 Not only does mucin production increase in pigs with experimental SD, but the mucus layer also becomes disorganized and lacks the striations apparent in age-matched control pigs. 101 In vitro mucin binding by B. hyodysenteriae is 7-fold higher in mucus samples from pigs with dysentery relative to controls, suggesting that the mucus niche formed during SD may substantially increase the quantity of mucus binding sites available to the spirochete. 101
Early microscopic lesions of SD include superficial mucosal necrosis, concurrent neutrophilic infiltration of the lamina propria, crypt elongation, hemorrhage, and abundant mucus secretion (Fig. 3). 139 By transmission electron microscopy, B. hyodysenteriae cells can be visualized along the colonic luminal surface and within epithelial cells, goblet cells, mucigen droplets, and the lamina propria. 39,40 Consistent ultrastructural changes in colonic epithelial cells of infected pigs include sparse and shortened microvilli, swollen mitochondria, and reduced numbers of organelles. 40 As lesions progress, the superficial mucosa becomes eroded and there is superficial hemorrhage and fibrinous exudation. 136 In clinical cases of SD, ciliated protozoa consistent with Balantidium coli are often observed along the mucosal surface and extending deeply into areas of ulceration. The attachment of spirochetes to the epithelium does not appear essential for lesion development as in vitro studies have not associated cellular attachment with cellular damage or invasion. 9,72,140
Gross lesions are limited to the cecum and colon and typically include a flaccid to fluid-filled colon with serosal hyperemia and variable expansion of the mesocolon by edema. Lesions can be multifocal and are frequently observed in the centripetal coils and at the apex of the spiral colon but may extend through the centrifugal portion to the terminal aspects with time. Affected segments of colon often have copious mucus, hemorrhage, and fibrinous exudate along the mucosal surface, and mucohemorrhagic to watery feces are observed (Fig. 2). Diarrhea occurs due to colonic malabsorption and this appears to be the result of impaired sodium and chloride transport. 4 Increased secretion of colonic mucins in SD, which are rich in sodium and potassium, may also increase the levels of these ions in dysenteric contents. 139 Additionally, the fecal microbiota of pigs that develop SD after “B. hampsonii” infection is significantly altered with reduced bacterial density and a relative increase in Firmicutes relative to Bacteroidetes when compared with noninoculated control pigs and inoculated pigs that do not develop disease. 25
Systemically, there is a significant increase in circulating neutrophils and monocytes during SD, 65 with an increase in the acute-phase proteins serum amyloid A (SAA) and haptoglobin at the onset of clinical disease 57 ; however, a febrile response is not typically observed. 58,65 The proinflammatory cytokine interleukin (IL) 1β increases significantly on the first day of clinical disease, 73 and SAA levels remain elevated more than 6 times the baseline during the first 3 days of mucohemorrhagic diarrhea. 65 During the dysentery period, there is a decrease in serum concentrations of some nonessential gluconeogenic amino acids including alanine, glutamine, and tyrosine. 66 As the disease progresses, SD is characterized by dehydration, metabolic acidosis, and hyperkalemia, with mortality in severely affected pigs. 42 Following experimental infection, recovery has been observed around 5 days after the development of clinical SD, which coincides with a decline in SAA, haptoglobin, and monocyte levels in the blood. 58 During recovery, IL-10 levels rise and appear to peak around day 7 of the recovery period. 73
A specific humoral immune response to outer membrane antigens of B. hyodysenteriae is induced following infection, 129 including antibodies against the membrane-associated lipoproteins A and B (SmpA and BmpB), 80,126 and antibodies to these proteins have been detected in serum as early as the first day of the recovery period. 65 Clinical SD is associated with the development of pathogen-specific immunoglobulin (Ig) G, IgA, and IgM in serum as well as local production of IgA in mucosal tissues. 105 Pigs that survive and recover from SD have been shown to be protected from reinfection for up to 17 weeks. 62
Diagnostic Methods
Historically, diagnosis of SD was based upon appropriate clinical signs, characteristic gross and microscopic lesions, and isolation of strongly β-hemolytic spirochetes from colonic tissue or feces. The advent of molecular diagnostic methods has increased opportunities for antemortem diagnosis, and numerous assays have been described for testing of feces and oral fluid samples including polymerase chain reaction (PCR) and in situ hybridization (ISH) targeting known pathogenic Brachyspira spp. 31,76,138 When postmortem samples are available, demonstration of spirochetes within histologic lesions is often readily accomplished with the aid of silver staining (Warthin-Starry) or more specifically with ISH, and characteristic delicate spirochetes can be observed within goblets cells, epithelial cells, crypt lumens, and luminal mucus (Fig. 4).
Microbial Culture
Selective culture for Brachyspira spp from clinical samples provides a high degree of diagnostic sensitivity for SD; however, the ability to recover spirochetes from clinical samples depends largely upon sample handling and the sample itself. Medication will greatly reduce the potential for isolating spirochetes so samples should be obtained from untreated, clinically affected animals, and the samples should be kept moist and refrigerated during transit to the laboratory. Brachyspira spp are typically isolated on selective media, 19,60,74 incubated anaerobically over 6–10 days, and are broadly characterized based upon phenotypic assessment of the isolate, which includes the degree of β-hemolysis on blood agar. 32,33 Brachyspira spp recovered from pigs with SD typically induce a strong degree of β-hemolysis and display a “ring phenomenon” around defects or slits in the agar. The presence of these phenotypic characteristics has been associated with virulence. 16 Speciation of Brachyspira from primary cultures is commonly performed using various biochemical tests, species-specific PCRs, partial sequencing of the nox gene, restriction fragment length polymorphism analysis, and matrix-assisted laser desorption time-of-flight mass spectrometry. 5,20,32,95,100,108,130,131 Epidemiologic studies exploring the diversity and relatedness of Brachyspira spp using MLST and multilocus variable number tandem repeat analysis (MLVA) have also been described. 50,77,90,91,97
Polymerase Chain Reaction
For many infectious agents, direct PCR from clinical samples is common and highly effective; however, it should be noted that for Brachyspira spp, direct PCR on clinical specimens may lack sensitivity relative to microbial culture. 16,102 Fortunately, primary cultures of Brachyspira spp can be readily differentiated by PCR, and assays have been described targeting the nox gene, 5,131 16S rRNA gene, 98 23S rDNA, 82 and the tlyA hemolysin gene. 34 Although this increased precision has greatly increased the specificity of diagnosis, dependence upon these assays may fail to detect novel species or potentially lead to misclassification. Accordingly, PCR testing of direct clinical samples with primer sets limited to the detection of B. hyodysenteriae and/or B. pilosicoli, 76 which is commonplace in many diagnostic laboratories, will most likely fail to detect novel potentially pathogenic Brachyspira spp.
In Situ Hybridization
In situ hybridization is an emerging and highly specific assay for direct detection of infectious agents within tissues. This assay combines the molecular specificity of PCR and direct visual confirmation of microscopy into one diagnostic test. Assays targeting the rRNA of B. hyodysenteriae, B. pilosicoli, B. murdochii, and B. hampsonii have been described. 10,18,61
Serology
Cross-reactivity between pathogenic and nonpathogenic Brachyspira spp in immunological tests using antisera developed from whole-cell preparations has historically been reported and suggests close similarity between at least some antigens in or on the surface envelope. 55,81 Enzyme-linked immunosorbent assay (ELISA)-based techniques for the detection of antibodies to lipopolysaccharide antigen are effective in differentiating B. hyodysenteriae infection from B. innocens; however, these assays are serotype specific and therefore not universally applicable. 64 Recently, novel serologic assays have been developed with improved specificity to detect circulating antibodies against B. hyodysenteriae-specific outer membrane lipoproteins and may be useful for detecting infected pigs 75,114 ; however, these assays have not yet found broad application. Recent work has also suggested that ELISAs run on meat juices at slaughter may be useful in detecting herds infected with B. hyodysenteriae. 114
Risk Factors for Infection and Disease Development
Poor environmental hygiene, overstocking, transportation, and cold temperatures have been described as predisposing factors to development of SD in endemically infected farms. 46 Wild rodents are recognized as potential carriers of Brachyspira spp and are an important potential source of interfarm and intrafarm spread. 6,63 Mice colonized with B. hyodysenteriae do not show signs of disease but may shed the organism for at least 200 days. 46 Feral pigs have been shown to harbor pathogenic Brachyspira spp and may also serve as a source of interfarm transmission in areas where feral pigs are common and in proximity to domesticated pigs. 96 Lagoon effluent is another potential source of environmental exposure to pathogenic Brachyspira spp, 41 particularly in systems where lagoon water recycling is used to flush gutters between pens. B. hyodysenteriae has been shown to survive in lagoon water samples in vitro for up to 17 days and in pit sludge for up to 21 days with longer survival at cooler temperatures. 112 Similarly, B. hyodysenteriae can survive in dysenteric pig feces for 48 days when stored at less than 10°C. 23 Open lagoon systems are frequently visited by wild birds. The potential importance of waterfowl as a source of spirochete transmission between farms is supported by the recent isolation of B. suanatina and B. hyodysenteriae from the feces of mallard ducks in Sweden, 59,103 and “B. hampsonii” isolation from the feces of migratory birds including lesser snow geese in Canada and mallard ducks and graylag geese in Spain. 85,110 Experimental inoculation of pigs with mallard-origin B. suanatina induced colonization, spirochete shedding, and clinical signs of SD in at least 1 pig. 103 Although experimental inoculation with a goose-origin Canadian isolate of “B. hampsonii” induced colonization and shedding but not clinical disease in pigs, 110 inoculation with a goose isolate of “B. hampsonii” from Spain resulted in spirochete shedding in all inoculated pigs, horizontal infection of a sentinel pig, and development of SD in 1 inoculated pig. 1 Taken together, these findings support the potential virulence of avian origin B. suanatina and “B. hampsonii” isolates to pigs and suggest the differences observed may be dependent on the laboratory and animal model. Further investigation into the potential virulence of avian origin, strongly hemolytic Brachyspira spp, is warranted.
Clinical expression of SD can be significantly influenced by dietary factors and the associated impact on the colonic microenvironment. Although there is some discord in the literature, it is generally accepted that feeding pigs highly digestible, cooked rice–based diets is associated with decreased expression of SD following B. hyodysenteriae infection, 99,113 while feeding of sources of certain resistant carbohydrates and lignins may increase disease expression. 127 Diets high in inulin have also been associated with decreased disease expression following experimental infection with B. hyodysenteriae, 44,127 and it has been postulated that this may be due to associated alterations in the colonic microbiota. 44,92 Such a role for the microbiota in the pathogenesis of SD is supported by previous studies where the presence of various synergistic bacteria was required for SD development in gnotobiotic pigs. 132 Diet-associated alterations in the colonic microbial community may increase or decrease the number of synergistic bacteria 30 or alternatively yield an increase in species that inhibit colonization by pathogenic Brachyspira spp. A recent study revealed that at least 4 different bacteria isolated from the porcine gastrointestinal tract can inhibit B. hyodysenteriae growth in vitro 71 and suggested a colonic environment favoring the presence of such bacteria may aid in the prevention of SD. Interestingly, during the recent period of SD reemergence in the USA, there has been an increase in availability of coproducts of biofuel production, such as distillers dried grains with solubles (DDGS), which are a source of insoluble dietary fiber and are frequently incorporated into commercial swine diets. Feeding 30% DDGS to swine results in a significant shift in the microbiome with a reduction in Lactobacillus spp and a predominance of Prevotella spp, 13 and feeding a diet containing 30% DDGS was associated with a shorter time to onset of SD and an increased overall disease incidence following experimental infection with B. hyodysenteriae relative to pigs fed no DDGS. 135 Additionally, experimental models using diet modification to include large quantities of soybean meal have been reported to enhance disease expression after B. hyodysenteriae infection. 57 In total, it is clear that dietary composition is a major contributing factor for SD development and may influence time to onset and severity of disease. Additional studies are warranted to further characterize the impact of specific dietary components on the colonic microbiota and the potential for practical dietary manipulation in the management of SD.
Treatment and Control
Pleuromutilins (tiamulin and valnemulin) are among the most widely used antibiotics to treat SD, given the frequent occurrence of tylosin and lincomycin resistance. 42 Resolution of diarrhea has been reported as early as 24 hours after 0.006% tiamulin water treatment, with viable spirochetes no longer detectable in the feces within 72 hours of treatment initiation in pigs experimentally infected with susceptible isolates of either B. hyodysenteriae or “B. hampsonii”. 137 Unfortunately, pleuromutilin resistance has been increasingly reported in porcine B. hyodysenteriae isolates from multiple countries including Czechoslovakia, 116 Germany, 107 Italy, 111 and Spain. 51 Moreover, this resistance appears to increase over time and on endemically infected farms, suggesting local selection of resistant clones. A recent analysis of Italian isolates suggests transnational spread of resistant B. hyodysenteriae throughout Europe. 111 US B. hyodysenteriae isolates are generally susceptible to tiamulin, although a slight rise in the MIC90 was observed between values reported in the 1990s and a more recent report in 2011, 24 suggesting that while tiamulin resistance is not currently widespread, it may be developing in the USA. Accordingly, the judicious use of these antimicrobials in the treatment of SD with close monitoring of MICs is warranted. In countries where available, carbadox has been shown to be efficacious as a prophylactic measure for SD 104 ; however, recent work has demonstrated that carbadox has the capacity to induce expression of gene transfer agents in B. hyodysenteriae and suggests this compound may facilitate lateral transfer of resistance genes between strains. 120
Control measures for SD should focus on elimination of environmental risk factors through cleaning and improved biosecurity. Moving recently medicated animals to a clean, segregated environment is often successful in elimination efforts. Extensive environmental cleanup with removal of all contaminated fecal material is essential for infected facilities, and a protocol of pressure washing, disinfection, and application of concentrated lime solution to environmental surfaces has been reported to be effective. 14 Improved biosecurity efforts should include aggressive rodent control, isolation from waterfowl, and reductions in foot and vehicle traffic. Infection is often transmitted from infected breeding animals to nursing pigs, and detection of subclinically infected breeding stock is imperative to reduce downstream contamination; however, testing strategies for breeding herds should account for low prevalence rates as a recent report suggests less than 2% of sows in subclinically infected herds may be shedding viable spirochetes. 29
Development of efficacious vaccines for protection against SD has been challenging as there is limited cross-protection against strains of different serogroups of B. hyodysenteriae. 42 Although numerous attempts have been made to create efficacious vaccines using both bacterins and attenuated strains, 27,36,43,54,56 commercial products are not widely available at present. Autogenous vaccines are available in some countries, but data are limited regarding the efficacy of these products. More recently, vaccine development efforts have focused on subunit and recombinant vaccines with varying levels of protection, 38,52,78 and a reverse vaccinology approach has been described for selecting potential antigens of B. hyodysenteriae. 115 The emergence of “B. hampsonii” and B. suanatina has further complicated the vaccination issue as any marketed SD vaccine would likely need to protect against at least 2 of these agents depending upon the geographic market. Given the reemergence of SD in North America, and the emergence of antimicrobial resistance to field strains in Europe and elsewhere, there is a growing need for efficacious SD vaccines globally.
Summary
Although SD has been recognized as a specific disease in swine for nearly a century, there is still much to be learned about the precise pathogenesis and the role of diet and the colonic microbiota in disease development and expression. Historically, B. hyodysenteriae was thought to be the only strongly β-hemolytic Brachyspira infecting swine and has been regarded as the sole etiologic agent of SD. However, recent identification of the strongly β-hemolytic B. suanatina and “B. hampsonii” from pigs with mucohemorrhagic colitis and the experimental reproduction of dysentery-like disease following infection with these agents suggests that the definition of SD should be expanded to encompass pigs demonstrating characteristic clinical signs and lesions along with the isolation of any strongly hemolytic Brachyspira spp from tissues or feces. Regardless of the Brachyspira spp involved, grower–finisher pigs with SD typically develop a mucohemorrhagic typhlocolitis and characteristic spirochetes are often numerous within lesions.
Diagnostic tests for Brachyspira spp detection have evolved dramatically in recent years with newer assays available that can improve the specificity of an etiologic diagnosis and frequently reveal interesting epidemiological relationships. However, this analytical specificity often comes at the cost of diagnostic sensitivity. Diagnostic approaches for SD should be broad enough to detect all potentially pathogenic spirochetes and also with sufficient analytical sensitivity to detect the low levels of spirochetes likely to be shed from subclinically infected carrier animals. Until such time that a unified molecular test is available that can identify unique virulence genes or other specific genetic targets universally present in SD-associated spirochetes, selective anaerobic culture should continue to be an integral part of Brachyspira detection and diagnosis of SD.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.