
Abstract
The natural transmission of vesicular stomatitis New Jersey virus (VSNJV), an arthropod-borne virus, is not completely understood. Rodents may have a role as reservoir or amplifying hosts. In this study, juvenile and nestling deer mice (Peromyscus maniculatus) were exposed to VSNJV-infected black fly (Simulium vittatum) bites followed by a second exposure to naive black flies on the nestling mice. Severe neurological signs were observed in some juvenile mice by 6 to 8 days postinoculation (DPI); viremia was not detected in 25 juvenile deer mice following exposure to VSNJV-infected fly bites. Both juvenile and nestling mice had lesions and viral antigen in the central nervous system (CNS); in juveniles, their distribution suggested that the sensory pathway was the most likely route to the CNS. In contrast, a hematogenous route was probably involved in nestling mice, since all of these mice developed viremia and had widespread antigen distribution in the CNS and other tissues on 2 DPI. VSNJV was recovered from naive flies that fed on viremic nestling mice. This is the first report of viremia in a potential natural host following infection with VSNJV via insect bite and conversely of an insect becoming infected with VSNJV by feeding on a viremic host. These results, along with histopathology and immunohistochemistry, show that nestling mice have widespread dissemination of VSNJV following VSNJV-infected black fly bite and are a potential reservoir or amplifying host for VSNJV.
Keywords
Vesicular stomatitis New Jersey virus (VSNJV) is an arthropod-borne virus in the genus Vesiculovirus, family Rhabdoviridae, that primarily affects horses, cattle, and swine, yet many other animal species are also susceptible to infection, including wildlife. 17 Infections in livestock are characterized by the development of vesicular lesions on the mouth, teats, and coronary bands, which are clinically indistinguishable from lesions of foot-and-mouth disease in cattle and pigs. 11
Infection of domestic animals with VSNJV has occurred as small endemic foci on Ossabaw Island, Georgia, as well as in parts of southern Mexico, Costa Rica, and northern South America. 1,2 In addition, VSNJV is associated with recurrent vesicular stomatitis (VS) epidemics in the western United States. 11
Transmission is associated with animal-to-animal contact and infected biting insects, such as sand flies (Lutzomyia spp) 15 , biting midges (Culicoides spp), 10 and black flies (Simulium spp). 3,6,7 However, several aspects of transmission by insects remain unclear, including how the virus is maintained between outbreaks and from what source insects are initially infected.
Serologic surveys to identify potential reservoir species involved in enzootic VSNJV maintenance have shown that mammals, including ungulates, carnivores, and rodents, may be naturally infected with VSNJV. 17 Of these susceptible species, only rodents have been shown to consistently develop viremia following experimental infection. 2 Based on serologic studies 17 and experimental infections, 2 the deer mouse (Peromyscus maniculatus) has been implicated as a potential VSNJV reservoir host. Naturally infected deer mice have been associated both temporally and spatially with the occurrence of VS epizootics. 17 Experimental infection demonstrated that deer mice can develop viremia and central nervous system (CNS) disease following intranasal or intradermal inoculation. 2 The virus also can be transmitted from VSNJV-infected black flies to naive black flies that are physically separated while cofeeding on deer mice, even though the mice do not develop detectable viremia. 7 In livestock, VSNJV can be transmitted mechanically to domestic pigs by black flies that had fed at the site of vesicular lesions in a previously infected host. 18 In addition, in cattle, VSNJV-infected black flies can transmit the virus to a naive host and cause clinical disease. 19,20
While the presence of neutralizing antibodies in animals demonstrates their susceptibility to VSNJV infection, it does not fully implicate them as reservoirs. According to the World Health Organization (WHO), a reservoir species must have virus circulating in the bloodstream at levels sufficient to infect a blood-feeding vector. 21 Detectable VSNJV viremia has been reported in rodents following direct and peripheral infection routes 2,16 but never following transmission via infected insect bite. Neither transmission accompanied by viremia in a host nor transmission to insects from a viremic host has been documented previously for VSNJV in any vector-host relationship. The present study was designed to evaluate the susceptibility of juvenile and nestling deer mice to VSNJV infection via black fly bite and also the ability of these mice to transmit the virus to naive black flies.
Materials and Methods
Two- to 3-day-old female Simulium vittatum (IS-7 cytotype) from a continuous laboratory colony (Department of Entomology, University of Georgia, Athens, Georgia) were used in this study. Peromyscus maniculatus were obtained from the Peromyscus Stock Center (Columbia, South Carolina) and housed individually in microisolation cages under controlled temperature conditions. The use of animals in this study was approved by the University of Georgia’s Institutional Animal Care and Use Committee (approval A2001-10076-ml). A 1997 Colorado equine VSNJV isolate was used in these studies. The virus was passaged and titrated in confluent Vero Middle America Research Unit (MARU, [Vero-M]) cell culture monolayers. 6
Experimental Design
Two separate experiments were conducted. In the first experiment, 20 female juvenile mice (aged 26–28 days) were separated into 2 groups (10 mice/group). The mice of group 1 (case Nos. 1–10) were anesthetized with 45 mg/kg Telazol (Zoetis, Florham Park, New Jersey) and 7.5 mg/kg xylazine and were intranasally infected with a suspension of VSNJV (10 7 pfu/ml) by placing a drop of the suspension at the opening to the nasal cavity, stimulating the mouse to inhale the droplet. On 1 or 2 days postinoculation (DPI), naive female black flies were allowed to feed on the intranasally infected mice (20–40 flies/mouse), and blood samples were collected in heparinized capillary tubes from the orbital sinus of mice and centrifuged to separate serum for virus isolation. Of these mice, 6 and 4 mice were euthanized on 1 and 2 DPI, respectively, immediately after blood sampling and fly feeding. In addition, following feeding, flies were collected for virus isolation. Group 2 mice (case Nos. 11–20) were anesthetized as described above, and the abdomen was shaved to expose skin, facilitating direct contact for insect feeding. Black flies, infected with VSNJV as previously described, 7 were allowed to feed on the shaved abdomen of each mouse until they no longer showed interest in feeding or until the anesthesia began to subside. To determine how many VSNJV-infected flies had fed on each mouse, black flies were dissected and examined for the presence of blood in the abdomen immediately after being allowed to feed on mice. Blood samples were collected in heparinized capillary tubes from the orbital sinus of all mice on 1 or 2 DPI, then centrifuged to separate serum for virus isolation. Then, 6 and 4 mice were euthanized on 1 and 2 DPI, respectively.
In a second experiment, 15 female juvenile mice (aged 26–28 days) and 8 nestling mice (aged 6–7 days) were used. Fifteen juvenile (group 3/case Nos. 21–35) deer mice were exposed to VSNJV-infected fly bite as described for group 2 above. Mice were observed daily for onset of clinical illness indicated by rapid breathing and/or paralysis in the hind legs. Mice with severe neurological signs (n = 4) were immediately euthanized. Six and 5 juvenile mice not showing clinical signs were euthanized on 10 and 11 DPI, respectively. Blood collected from the orbital sinus from these mice on 3 DPI and at euthanasia was processed for viral isolation and antibody titration. Eight nestling mice (group 4/case Nos. 36–43) were housed with the mother in a single microisolation cage. VSNJV-infected flies were allowed to feed on the anesthetized nestlings without shaving the abdomen since they had not developed a significant amount of fur. Subsequently, these mice were divided in 2 subgroups of 4 animals each, and naive flies were allowed to feed on anesthetized nestlings on 1 and 2 DPI, respectively. Immediately following feeding, the mice were euthanized as described above, and the flies were submitted for dissection and viral isolation. Flies with obvious swelling of the abdomen indicating blood feeding were homogenized individually.
All mice were euthanized by subcutaneous injection of barbiturate. Necropsy was performed in all euthanized mice and mice that died from both experiments, and samples of head, brain, spinal cord, skin, lung, and heart were collected and fixed in 10% neutral buffered formalin. In addition, samples of brain of nestling mice were collected for viral isolation.
Virus Isolation, RT-PCR, and Antibody Titration
Black flies were homogenized in groups of 1 to 3 black flies in 1.8-ml vials each containing 2 copperhead ball-bearing pellets and 0.5 ml virus isolation media (minimum essential media [MEM] with 5% fetal bovine serum and 2% antibiotics). For virus isolation, either 10 μl of serum or black fly samples was inoculated onto individual wells of a 12-well plate with confluent Vero-M cell culture monolayers and incubated at 37°C in a 5% CO2 atmosphere for 1 hour. Cell cultures were examined daily for 3 days for evidence of cytopathic effects (CPE). Before inoculation, black fly samples were centrifuged to settle debris, and 100 μl of the resulting supernatant was used for virus isolation. Virus isolates were confirmed by reverse transcriptase polymerase chain reaction (RT-PCR) as described previously. 12
A previously described plaque assay 6 was used to quantify VSNJV in virus isolation-positive black fly and serum samples. Briefly, 10-fold serial dilutions of positive serum or black fly samples were used as the cell culture inoculum. Two 0.5-ml portions of each dilution were aliquoted onto each of 2 wells of 2-day-old confluent Vero-M cell culture monolayers in 24-well cell culture plates and incubated at 37°C for 1 hour. After removal of the inoculum, the cells were overlaid with a 1% gum tragacanth/1× MEM solution supplemented with 3% heat-inactivated fetal bovine serum and 2× antibiotic/antimycotic (200 U/ml penicillin, 200 μg/ml streptomycin, and 500 ng/ml amphotericin B). After incubating for 3 days at 37°C, cells were fixed by adding 20% formalin to the overlay and then stained with 5% crystal violet. Virus titers are expressed as plaque-forming units (pfu) per black fly or milliliters of serum.
Antibody titration was performed using a microtiter serum neutralization test. 9 Briefly, for microtiter serum neutralization tests, 2-fold dilutions (maximum 1:256) of heat-inactivated serum samples were challenged with a virus solution (1000 TCID50/25 μl) to identify an increase in VSNJV neutralizing antibodies. A 4-fold or greater increase in neutralizing antibodies was considered evidence of seroconversion.
Histology and Immunohistochemistry
Formalin-fixed tissues were embedded in paraffin, sectioned at 4 μm, and stained with hematoxylin and eosin. Tissue sections from juvenile mice from the first experiment (case Nos. 1 and 2/intranasal infection; case Nos. 11, 13–17, and 20/infection by fly bite), as well as from mice of the second experiment (case Nos. 21, 23, 25, 26, 31, 32, and 34/juveniles; case Nos. 35–43/nestlings), were also stained for VSNJV antigen by immunohistochemistry (IHC).
Immunohistochemistry was performed using an alkaline phosphatase–conjugated streptavidin-biotin technique similar to what has been described and validated previously. 2 Tissues were sectioned at 4 μm, mounted on slides, and deparaffinized, and antigen was retrieved by boiling in 0.01 M citrate buffer (pH 6) for 7 minutes in a microwave oven. Sections were blocked with Power Block (Biogenex, Fremont, California) for 7 minutes at room temperature and then incubated with a 1:1500 dilution of anti-VSNJV hyperimmune mouse ascitic fluid kindly provided by Dr. R. B. Tesh. Sections were washed, and biotinylated anti-mouse and alkaline phosphatase–conjugated streptavidin label (Biogenex, San Ramon, California) were used sequentially. Fast red (Biogenex) was used as the chromagen/substrate, and sections were counterstained with hematoxylin. CNS sections of mice previously infected with VSNJV were used as positive controls. 2 The primary antibody was substituted by homologous nonimmune sera as a negative control.
Results
Clinical Outcome
One juvenile mouse (case No. 10) from group 1 was found deceased on 1 DPI, probably due to trauma during the manipulation process. Clinical disease, characterized by neurological signs including rapid breathing and/or paralysis originating in the hind legs, developed in 5 juvenile mice that were exposed to VSNJV via fly bite (group 3/case Nos. 21, 23, 25, 26, and 31). One mouse was found deceased (case No. 26) and the other mice were euthanized on 6 to 8 DPI due to severe neurological signs. Juvenile mice from groups 1 and 2 did not exhibit clinical signs since they were euthanized on 1 or 2 DPI. Clinical signs similar to group 3 were observed in the nestling mice (group 4).
Virus Isolation and Seroconversion
Virus was detected in the serum on 1 or 2 DPI in 9 of 10 intranasally infected group 1 mice by virus isolation and confirmed by RT-PCR. Virus titers in the serum of these mice ranged from 7.5 × 102 to 7.5 × 104 pfu/ml. However, virus was not recovered from any naive black flies that fed on the viremic mice (0/266 flies).
Poor feeding by VSNJV-infected black flies was observed for group 2 mice. Based on observation during dissection, the number of flies containing discernible amounts of blood in the abdomen ranged from 0 to 3 flies per mouse. Viremia was not detected in the serum of any of these mice on 1 or 2 DPI.
Virus was not detected in the serum of any juvenile mice (group 3) exposed to VSNJV via infected fly bite on 3, 10, or 11 DPI. Although viremia was not detected, transmission was confirmed by seroconversion in 7 mice (case Nos. 21, 23, 25, 27, 31, 32, and 34). Neutralizing antibody levels for all mice were <1:8 on 3 DPI, and a 4-fold or greater increase in neutralizing antibody titer was considered seroconversion. Neutralizing antibody titers ranged from 1:16 to 1:≥256, and titers in the 5 mice that developed clinical illness included both extremes of this range. The 3 mice that seroconverted but did not demonstrate clinical signs of illness (Nos. 27, 32, and 34) had neutralizing antibody titers of 1:32, 1:64, and 1:≥256, respectively.
In group 4, virus was detected in the serum of 8 of 8 nestling mice on 1 and 2 DPI with virus titers ranging from 1.0 × 104 to 9.5 × 106 pfu/ml. In addition, VSNJV was detected in the brain of all 8 nestling mice by virus isolation and confirmed by RT-PCR. Virus was recovered from 12% (15/126) of the total number of naive flies allowed to feed on all viremic nestling mice; of these, virus was recovered from 24% (15/63) of those flies that were suspected of having had a blood feeding. The nestling mouse (case No. 42) with the highest virus titer produced the highest percentage (60%) of VSNJV-infected flies, and the lowest virus titer was correlated to the lowest percentage of VSNJV-infected flies (Table 1). One nestling (case No. 40) was an exception to this observation since only 3 flies fed on this mouse. Virus titers of flies that fed on viremic nestlings were determined immediately after feeding and varied from <50 to 1.6 × 103 pfu/ml. As shown in Table 1, virus titers in VSNJV-infected flies were not directly proportionate to virus titers of nestling mice. The flies with the highest quantity of virus detected were not infected from the mouse with the highest virus titer. Overall, virus was recovered from a higher percentage of flies after feeding on nestlings with virus titers ≥105 pfu/ml.
Transmission of Vesicular Stomatitis New Jersey Virus (VSNJV) Between Viremic Nestling Deer Mice and Black Flies.a
Abbreviations: NA, not applicable.
aThe data show virus titers in serum of nestling mice 1 or 2 days after exposure to feeding by black flies infected with VSNJV, and the titers and prevalence of virus recovery from a different cohort of previously uninfected flies after feeding on the nestling mice.
bNaive black flies were allowed to feed on VSNJV-infected nestling mice on day 1c or day 2d after infection of the mouse.
Histopathology and Immunohistochemistry
Group 1
All 10 mice inoculated intranasally had lesions in the nasal cavity that ranged from multifocal necrosis of the olfactory mucosa on 1 DPI to extensive necrosis with a mild to moderate infiltration of neutrophils in the submucosa in mice on 2 DPI. Three of the 6 mice euthanized on 1 DPI had dermatitis at the site of fly bite characterized by focal, mild infiltrations of neutrophils, mononuclear cells, and mast cells in the superficial dermis associated with edema in 1 mouse. On 1 DPI, 1 mouse developed a severe, diffuse purulent bronchiolitis and bronchopneumonia. On 2 DPI, mice had multifocal pyknotic cellular debris in multiple olfactory nerve fibers as they entered the olfactory bulb. IHC was performed on 2 mice (case Nos. 1 and 2), and both had multifocal staining for VSNJV in the olfactory epithelium and in Bowman’s glands. Both of these mice also had staining of olfactory nerve fibers from the nasal mucosa to the level of the olfactory bulb. Skin, lung, and brain were immunonegative for VSNJV in these 2 animals.
Group 2
Of the first group of 10 juvenile mice exposed to VSNJV-infected fly bites, 3 of 6 mice on 1 DPI and 2 of 4 mice on 2 DPI had dermatitis at the area of feeding. This was characterized by multifocal serocellular crusting (5/5), mild epidermal hyperplasia (2/5), and diffuse and mild infiltration of a mixed population of inflammatory cells within the superficial dermis that was composed mainly of macrophages, plasma cells, and neutrophils, as well as fewer lymphocytes and mast cells (5/5). Two of these mice had staining for VSNJV by IHC. One (case No. 13) had staining within adipocytes and hair follicles, and another mouse (case No. 20) had staining in the cytoplasm of epidermal keratinocytes and in inflammatory cells in the superficial dermis. In group 2, lesions were not present in brain, spinal cord, nasal mucosa, lung, and heart. Viral antigen was not visualized in these tissues.
Group 3
In the other 15 juvenile mice exposed to VSNJV-infected fly bites, dermatitis at the site of feeding occurred in 5 of the mice, 4 of which (case Nos. 21, 23, 25, and 26) became sick or died. The dermatitis visualized in group 3 was similar as described for group 2. However, viral antigen was not detected in the skin of mice of group 3 by IHC. Only the 4 mice that became sick or died had lesions in the CNS.
In the spinal cord, there was moderate to severe, multifocal to coalescing infiltration of lymphocytes and plasma cells admixed with rare neutrophils in the leptomeninges, severe necrosis of ependymal cells lining the central canal, and multifocal neuronal necrosis affecting both dorsal and ventral horns (Fig. 1). Neuronal necrosis associated with neutrophilic infiltration was also observed in the spinal ganglia (Fig. 2).
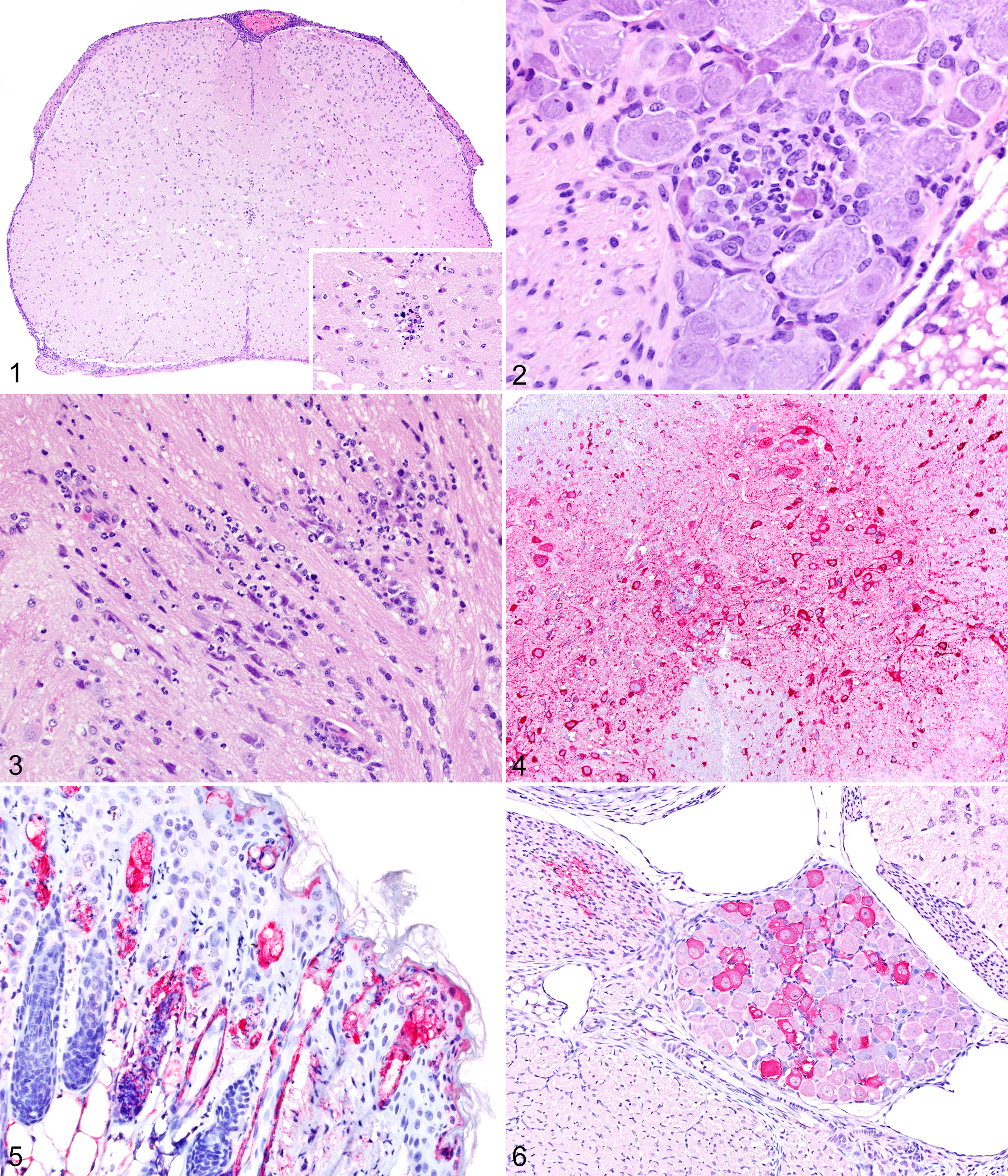
In the brain, lymphoplasmacytic leptomeningitis was most severe in the brainstem but affected the entire brain, including olfactory bulbs, and was most severe ventrally. The lesions were characterized by diffuse infiltration of a large number of neutrophils, severe multifocal neuronal necrosis (Fig. 3) in which necrotic neurons were often surrounded by neutrophils, and multifocal areas of hemorrhage. Necrosis of ependymal cells lining the fourth ventricle was also observed.
By immunohistochemistry, strong cytoplasmic immunolabeling for VSNJV antigen was detected in necrotic and viable neurons and their processes throughout the spinal cord (Fig. 4), in brainstem nuclei, and in dorsal root ganglia. VSNJV antigen was also detected in a large number of necrotic and viable ependymal cells lining the central canal of the spinal cord and the fourth ventricle. One mouse (case No. 26) had immunostaining for VSNJV in olfactory nerves, but there were no histopathologic changes in these nerves. Lesions and/or viral antigen were not detected in the olfactory bulb, cranial nerve ganglia, nasal mucosa, heart, lung, skeletal muscle, brown fat, or tongue epithelium of juvenile mice.
Group 4
In the group of nestling mice exposed to VSNJV-infected fly bites, skin lesions were seen in 5 (case Nos. 36, 37, 41, 42, and 43). On 1 DPI, in 2 mice (case Nos. 36 and 37), multifocal areas of hemorrhage were observed in the skin, but viral antigen was not detected. Three of 4 mice euthanized on 2 DPI had skin lesions. One (case No. 42) had a diffuse and mild lymphoplasmacytic and neutrophilic dermatitis without detectable viral antigen, and 1 had a moderate lymphoplasmacytic dermatitis and subcutaneous edema with a small amount of detectable viral antigen (case No. 43). One mouse (case No. 41) had extensive serocellular crusting and severe dermal and subcutaneous edema with severe neutrophilic infiltration and exocytosis. In this nestling, massive amounts of viral antigen were observed multifocally within hair follicles and adipocytes (Fig. 5), and underlying panniculus muscle fibers were positive for VSNJV by IHC.
All nestling mice in group 4 euthanized on 2 DPI had microscopic lesions in the CNS characterized by necrosis of ependymal cells lining the central canal of the spinal cord, the fourth ventricle, and the lateral ventricles. The capillaries throughout the brain were diffusely and markedly congested with swollen endothelial cells. Occasionally, the capillaries were surrounded by a small number of neutrophils, and there were multifocal areas of hemorrhage. There were a small number of neutrophils and lymphocytes within the leptomeninges.
By IHC, all nestling mice in group 4 euthanized at 1 DPI had a VSNJV immunolabeling of few neurons in dorsal root ganglia, in axons of the spinal nerve (Fig. 6), and in few scattered positive neurons in the cerebral cortex, cerebellum, brainstem, and olfactory bulb. On 2 DPI, massive amounts of viral antigen were detected in many neurons throughout the brain, including the cerebral cortex, hippocampus, olfactory bulb, brainstem, and thalamus. Immunolabeling was also present in many neurons in dorsal and ventral horns of spinal cord and dorsal root ganglia and trigeminal ganglia, as well as in olfactory nerves.
Lesions were not observed in the nasal respiratory mucosa, but 6 of 7 nestling mice had detectable viral antigen in the olfactory epithelium and Bowman’s gland. Multifocal areas of myocardial necrosis containing massive amounts of viral antigen were seen in 5 of 7 nestling mice. On 2 DPI, 3 mice had multifocal, moderate necrosis of striated muscles around the spinal column, muscles of the head, and tongue; these muscles contained a large amount of viral antigen. VSNJV antigen was also observed in areas of striated or myocardial muscle without lesions. Immunolabeling was also seen in brown fat of 2 mice (case Nos. 40 and 43) associated with or without foci of necrosis. Although lung lesions were not observed, there was multifocal immunolabeling for VSNJV in alveolar septa in 2 of 7 nestling mice (case Nos. 39 and 43).
Discussion
In this study, both juvenile and nestling deer mice developed CNS disease after exposure to VSNJV-infected black fly bites. The morphologic and immunohistochemical findings suggest that VSNJV in juvenile deer mice reached the CNS via sensory nerves, whereas in nestling mice, the hematogenous route was also involved since these mice developed viremia. Importantly, these viremic nestling mice were able to transmit the virus to naive black flies, establishing 2 essential aspects of arboviral transmission. Although viremia was also detected in juvenile deer mice intranasally infected with VSNJV, the virus titers were not sufficient to infect blood-feeding flies, which suggests that specific host and vector factors may enhance virus replication and transmission in nestling mice.
In the present study, juvenile and nestling deer mice exposed to VSNJV-infected flies developed severe lesions in the CNS. Juvenile mice (group 3) exposed to VSNJV-infected flies were euthanized on 6 to 8 DPI due to severe neurological signs. Lesions in these mice were characterized by a diffuse lymphoplasmacytic leptomeningitis, necrosis of ependymal cells lining the central canal in the spinal cord, and multifocal neuronal necrosis associated with an infiltration of neutrophils in dorsal root ganglia, in both ventral and dorsal horns of the spinal cord and in a large number of neurons of the brainstem. Viral antigen was detected by IHC in the cytoplasm of viable and necrotic neurons of all these areas, including the axons of spinal nerves. No other lesions, or VSNJV antigen, were observed in other tissues. Despite dermatitis visualized in some mice, VSVNJ antigen was not detected in skin on 6 to 8 DPI. However, on 1 and 2 DPI, juvenile mice exposed to VSNJV-infected fly bites (group 2) had detectable VSNJV antigen in epithelial cells within the epidermis, in inflammatory cells, and in the dermis.
The presence of VSNJV antigen in the skin supports the hypothesis that when juvenile mice are exposed to VSNJV-infected flies, VSNJV reaches the spinal cord through peripheral nerves via the sensory pathway, whose receptors are located on or near the body surface. 4 In this way, the virus may first reach the dorsal root ganglia as suggested by the presence of VSNJV antigen on neurons and extending axons. Projections of neurons of the dorsal root ganglia enter the spinal cord through the dorsolateral sulcus and synapse with interneurons involved in reflex activity or with neurons that project cranially to the brainstem. 4 Large amounts of VSNJV antigen were found in neurons throughout the brainstem, consistent with involvement of the sensory pathway in VSNJV pathogenesis. In addition, the presence of viral antigen in ependymal cells lining the fourth ventricle and central canal of spinal cord suggests that dissemination within the ventricular system may occur in juvenile deer mice exposed to VSNJV-infected black fly bites. In contrast, in intradermally VSNJV-infected juvenile deer mice, viral antigen was widespread in many regions in the brain and also in the heart. 2 In this case, viral dissemination was attributed to hematogenous infection. 2
Unlike juvenile mice, all nestling mice from the present study developed viremia on 1 and 2 DPI after being exposed to VSNJV-infected flies. The lesions in the CNS of nestling mice on 2 DPI were widespread throughout the brain, and VSNJV antigen was detected in a large number of neurons in many regions of brain and spinal cord, as well as in myocardium, striated muscles around the spinal column, tongue, lungs, olfactory epithelium, brown fat, and skin. Although the sensory pathway was also involved, viremia associated with widespread viral antigen in many organs, including the CNS, suggests that VSNJV spreads by a hematogenous route in nestling mice, which differed from the infection in juvenile mice.
The pathogenesis of VSNJV, when inoculated intradermally in a previous study, was similar between juvenile and nestling mice, with both age classes developing viremia. 2 This could be attributed to the amount of inoculum, which is most likely higher in intradermal inoculation compared with fly bite. However, in nestling mice of the present study, virus dissemination was more widespread and prominent compared with intradermally inoculated nestlings. 2 Since delivery of VSNJV by mosquitoes has been shown to induce a neutralizing antibody response more efficiently than injection, 5 it is possible that specific factors of the vector are important for infection of deer mice with VSNJV from blood-feeding flies. A previous study has shown that specific factors of the vector may play an important role in VSNJV transmission to rodents by blood-feeding flies. 10
Following intranasal VSNJV inoculation of juvenile deer mice, virus travels through the olfactory nerves to the olfactory bulb and subsequently to other parts of the brain. 2 In juvenile mice inoculated intranasally in the current study, lesions and viral antigen were found primarily in the olfactory epithelium and were similar to those described previously. 2
The presence of virus in the serum of intranasally infected juvenile deer mice in this study is consistent with previous experimental infections using intranasal inoculation. 2 However, because intranasal infection is not likely a naturally occurring mode of transmission, this may not reflect the pathogenesis of the natural disease. In addition, the inability to recover virus from naive flies that fed on these viremic juveniles suggests that the virus titers in these mice were not sufficient to infect blood-feeding flies. Although undetectable immediately after feeding, virus may have replicated to detectable and infective levels during the extrinsic incubation period. Unlike the intranasally infected juveniles, no juvenile mice exposed to VSNJV-infected fly bite (group 2) developed detectable viremia. It cannot be excluded that poor feeding by these flies could result in low amounts of viral inoculum resulting in no detectable viremia on 1 and 2 DPI. However, viral antigen was observed in the skin of these juvenile mice, demonstrating that viral transmission occurred in this group.
In contrast to juvenile mice exposed to VSNJV-infected fly bite, all nestling mice developed detectable viremia and, unlike the viremic intranasally infected juveniles, had virus titers sufficient to infect naive flies. The virus titers of nestling mice were generally higher than those of the intranasally infected juvenile mice and may be attributable to specific host factors that allow for enhanced virus transmission and replication in nestling mice following infection by infected insect bite. Thus, the results demonstrate that nestling deer mice have an increased susceptibility to VSNJV infection when inoculated through black fly bite. Similarly, in other rodent species, young mice are more susceptible to VSNJV infection. 2,8,13,14,16 Based on the titers obtained from nestling mice, it is likely that a minimum of approximately 104 pfu/ml of virus in the serum of deer mice is necessary for the infection of black flies. It is important to note here that the presence of VSNJV in black fly homogenate indicates transmission only and, for our purposes, was assumed to lead to disseminated infection in at least a portion of these flies, making them capable of transmitting virus. Virus titers in the nestling mice varied proportionately with recovery of virus in flies, indicating that more flies can potentially become infected by feeding on a host with a higher amount of virus in the blood.
In the absence of viremia, such as for the juveniles in the second experiment, transmission was also confirmed by seroconversion. Three juvenile mice developed relatively high neutralizing antibody levels without conspicuous clinical illness. This suggests that juvenile deer mice may be infected with VSNJV without displaying clinical signs and that mice of this age are capable of mounting a sufficient antibody response to limit development of disease and to survive VSNJV infection.
The detection of viremia in nestling mice is the first demonstration of viremia in a host following transmission by VSVNJ-infected insect bite. Although previous experimental infections have suggested the potential of deer mice to serve as reservoir or amplifying hosts for VSNJV because of their susceptibility to systemic infection, 2,16 especially development of relatively high virus titers in serum, these studies used intradermal injection or unnatural routes of infection rather than infection by black fly bites, the natural route of arboviral transmission. In the present study, the development of detectable viremia following infective insect bite satisfies one essential criterion for arboviruses. 22 Conversely, in this study, the infection of black flies by feeding on viremic nestling mice is the first demonstration of VSNJV infection of an insect by feeding on a viremic host, further fulfilling the definition of an arbovirus. Together, these 2 demonstrations, viremia following VSNJV-infected insect bite and infection of insects via blood feeding on a viremic host, were able to reproduce principal components of what may occur in nature with the enzootic VSNJV transmission cycle. Besides the involvement of a viremic host in VSNJV transmission, a nonviremic host may also exert an important role in VS epidemiology. VSNJV can be transmitted horizontally by black flies to uninfected flies by cofeeding on the same nonviremic deer mouse. 7 In addition, mechanical transmission of VSNJV by black flies has been demonstrated in domestic swine. 18
In summary, the present study demonstrates that VSNJV-infected black flies were able to transmit the virus to juvenile and nestling mice, which were susceptible to infection developing significant CNS lesions. Naive flies were infected after feeding on viremic nestling deer mice, which could be potential reservoir hosts of VSNJV and may play an important role in the enzootic maintenance of the virus.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The project was supported by the National Research Initiative of the USDA Cooperative State Research, Education and Extension Service, grant number 2001 35204 10069. Supplemental support was provided through Cooperative Agreement 2002-9613-0032-CA, Veterinary Services, APHIS, USDA. L. P. Mesquita was supported by a doctoral and research internship abroad grant from São Paulo Research Foundation (FAPESP), process no. 2014/20631-8.