
Abstract
Given the irreversible nature of nephron loss, aging of the kidney is of special interest to diagnostic and toxicologic pathologists. There are many similarities among histologic lesions in aged human and canine kidneys, including increased frequency of glomerulosclerosis, interstitial fibrosis, and tubular atrophy. Unfortunately, there are few studies in which renal tissue from aged healthy dogs was adequately examined with advanced diagnostics—namely, transmission electron microscopy and immunofluorescence—so age-associated changes in canine podocytes and glomerular basement membranes are poorly characterized. An age-associated decrease in the glomerular filtration rate in humans and dogs (specifically small breed dogs) has been documented. Although lesions in aged rats and mice differ somewhat from those of aged dogs and humans, the knowledge gained from rodent models is still vital to elucidating the pathogenesis of age-associated renal disease. Many novel molecules implicated in renal aging have been identified through genetically modified rodent models and transcriptomic and proteomic analysis of human kidneys. These molecules represent intriguing therapeutic targets and diagnostic biomarkers. Likewise, influencing critical pathways of cellular aging, such as telomere shortening, cellular senescence, and autophagy, could improve renal function in the elderly.
The concept of aging as a driving force for disease is especially relevant in organs with limited ability to regenerate, such as the kidneys, heart, and central nervous system. With respect to the kidney, normal wear and tear can lead to cellular senescence as well as loss of structural and/or functional units, specifically nephrons. Over time, renal architecture is modified, and renal reserve is diminished. Investigation of the mechanisms of remodeling that occur in aged kidneys has been of intense interest in humans and in rodents, with numerous recently published reviews.* Most of the focus on age-associated nephropathy in humans stems from the fact that the percentage of the global population ≥60 years is projected to increase from 8.6% in 1980 to 21% in 2050. 1 Costs associated with treatment of chronic kidney disease rose 53.6% between 2008 and 2012 in American patients ≥65 years old, whereas general medical costs increased only 11.5% during the same time frame. 2 Given the prospect of increasing health care costs in the growing geriatric population, slowing the progression of renal aging is expected to save billions of dollars.
In rodents, the investigation of renal aging has 2 aims. First, research of the molecular mechanisms of age-associated renal disease in rodents might shed light on similar processes in humans. Second, aged rodents are commonly examined in chronic toxicity studies. As such, age-associated lesions should not be attributed to exposure to the compound of interest.
Although canine epidemiologic data are not as easily accessible or as accurate as they are for humans, geriatric patients are a growing age group at tertiary care veterinary hospitals. Furthermore, aged dogs submitted for autopsy might have limited clinical information, which complicates the pathologist’s ability to discern the significance of renal lesions. In a similar vein, as biopsies are more frequently used to diagnose renal disease, age-associated lesions need to be interpreted correctly. In 2009, 29% of the renal biopsies submitted to the International Veterinary Renal Pathology Service were from dogs >7 years old, whereas the percentage had increased to 45% in 2014 (unpublished data, 2015).
Unfortunately, most of the literature on age-associated renal lesions in dogs is hindered by lack of adequate clinical data or misclassification of lesions. The aims of this review are (1) to describe the spectrum of renal lesions and their associated clinicopathologic findings in aged dogs and (2) to discuss the molecular basis of renal aging, with the caveat that much of the data for the latter topic have been derived from humans and rodents.
Morphologic Changes in Aged Canine Kidneys
Before we discuss specific lesions associated with renal aging, a general review of renal development and nephron maturation is in order. Early in development (week 5 in humans, embryonic day 11 in mice), the ureteric bud grows cranially from the mesonephric duct and impinges into the metanephric mesenchyme, where it then undergoes synchronous dichotomous branching to form the collecting duct system. This process induces the overlying mesenchymal tissue to condense and transition to become groups of epithelial cells called renal vesicles. These vesicles then elongate to form primitive nephron structures—S-shaped bodies—which have a layer of podocyte precursors at their proximal end. Eventually, a capillary grows into the structure to form a primitive glomerulus and contact the podocyte progenitors during the capillary-loop stage. The podocyte precursors flatten, covering the glomerular capillaries, while at the same time the glomerular capillaries branch so that the surface area of the glomerular filtration barrier is dramatically increased. 83 Polarization of gene expression patterns along the length of the neonephron occurs at the renal vesicle stage and drives the development of proximal and distal tubular segments. 42 Given the intricate nature of renal development, delays or interruptions of any of these steps can affect overall nephron number and functional renal reserve.
Unlike humans, the canine kidney continues to develop postnatally. Until approximately 8 days of age, there is a distinct subcapsular zone in which new nephrons are formed. Beneath this nephrogenic zone, the nephrons (specifically the glomeruli) become increasingly more mature (Fig. 1) toward the corticomedullary junction. Although it is not uncommon to detect fetal or immature glomeruli until 21 days of age, these should dwindle in number after that time. 20 Notably, infection, stress, or toxin exposure during this period of postnatal development might interrupt the maturation process and lead to increased frequency of fetal glomeruli. Identifying a few scattered fetal glomeruli in normal canine renal specimens is possible, with smaller dogs being more commonly affected than larger dogs (personal observation). In renal biopsy specimens, the fraction of glomeruli that are fetal and their distribution should always be reported because these nephrons are assumed to be nonfunctional. Even if the primary disease process is not developmental in nature (eg, immune complex–mediated glomerulonephritis), reporting the percentage of fetal glomeruli will provide important information to the clinician regarding functional nephron mass and renal reserve.
Nephron number in Beagles was estimated to range from 445 000 to 589 000 per kidney 20,30 ; however, other breeds have not been reported. A wider range of nephron number would not be surprising, given the marked variation in other breed-associated phenotypes. In humans, nephron number has been estimated to range >9-fold, from 210 000 to 2 000 000. 31,32 Low nephron endowment (so-called nephron paucity) can limit functional renal reserve and exacerbate renal injury in humans and rodents. 8,51,65 A similar estimate of nephron number and its relationship to progression of renal disease have not been reported in dogs. Last, there are numerous familial and hereditary diseases of the canine kidney (Suppl. Table 1) that can affect functioning nephron mass.
Age-associated renal lesions are discussed by compartment, starting with the glomerulus (Figures 1–3) and moving to the tubulointerstitium (Figure 4) and vasculature. Lesions reported in dogs are described first, and relevant comparisons to humans and rodents follow.
Glomerular Lesions
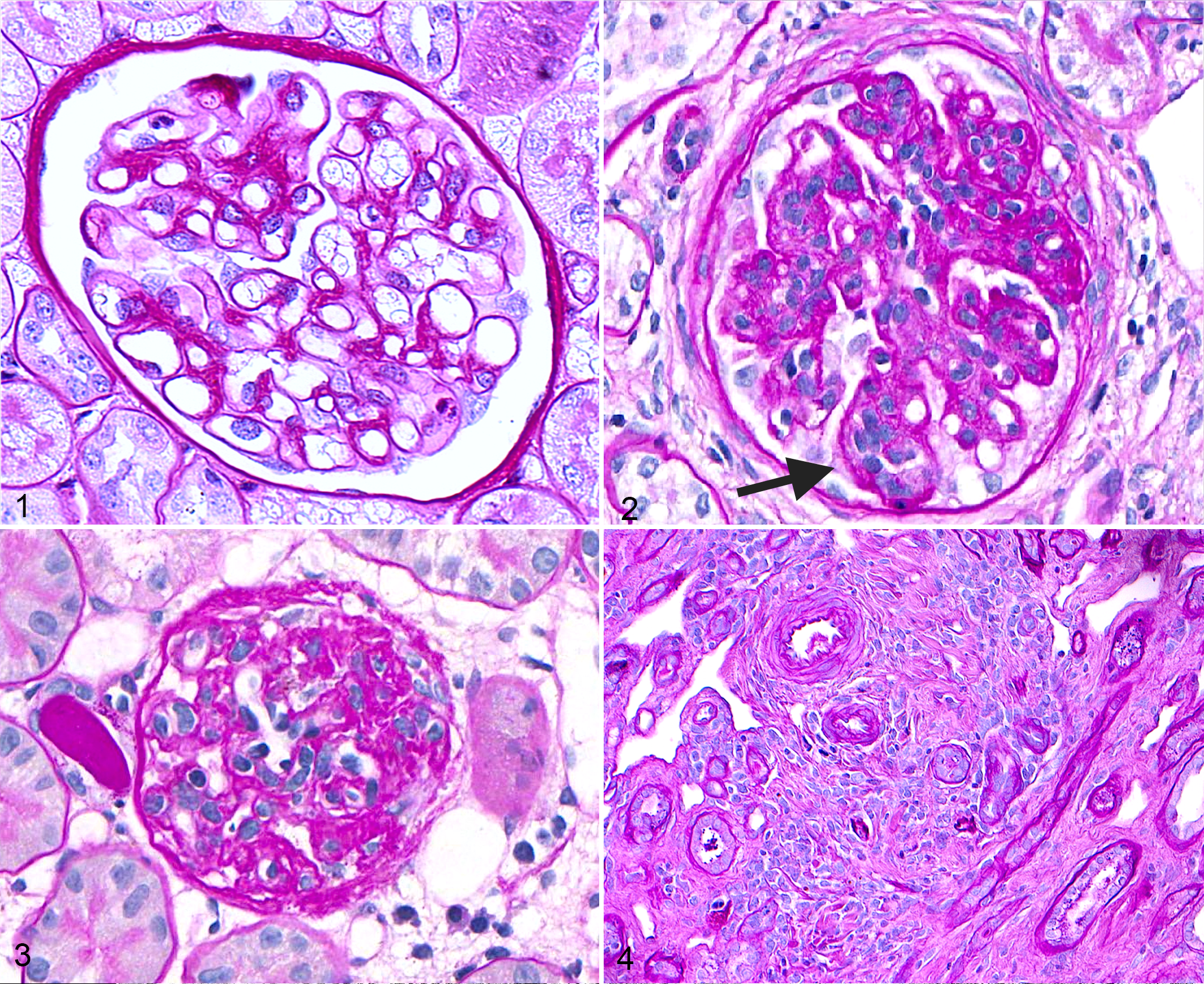
Glomerular lesions reported in older dogs include increased frequency of glomerulosclerosis (GS; Fig. 2) and obsolescent glomeruli (Fig. 3). Terminology and definitions were provided by the recent World Small Animal Veterinary Association Renal Standardization Study Group’s (WSAVA-RSSG’s) classification of glomerular diseases. GS (Fig. 2) is “increased extracellular matrix leading to obliteration of capillary lumens and consolidation of part or most of the tuft,” whereas obsolescent glomeruli (Fig. 3) are “small glomerular remnants composed of residual matrix and few cells.” 13
Studies have reported a wide range in the prevalence of dogs with GS (Table 1), and this variability is likely due to inconsistency in terminology and definitions. One autopsy study identified GS in 7 of 13 pet dogs >7 years old; however, the investigators separated GS from glomerular fibrosis, which was identified in 9 of the dogs. 29 This distinction is unwarranted; all glomerular scarring should be categorized as GS because it is extracellular matrix synthesis predominantly by mesangial cells, whereas fibrosis implies collagen synthesis by fibroblasts. 53 Another autopsy study of 159 purpose-bred laboratory Beagles reported that all dogs >2 years old had GS. This study included the presence of synechiae (adhesions between the glomerular tuft and Bowman capsule) as indicative of GS, which likely led to overestimation of affected glomeruli. 57 An autopsy study of 145 Portuguese Water Dogs identified GS in 63% of cases. Although specific ages were not provided, GS was significantly associated with the dog’s age at death. Diagnostic criteria for GS were not reported. 11 One study examined serial renal biopsies from 5 Beagles (>8 years old) that were used as controls to investigate the effects of hydrocortisone therapy. Global GS, ranging from mild to severe, was identified in 1 control dog at the start of the study and in all control dogs 16 and 24 weeks later. Again, definitions of terminology were not provided. 74 Interestingly, neither GS nor glomerular obsolescence (as defined by the WSAVA-RSSG) was identified in renal wedge biopsies from 2 healthy Greyhounds >8 years old. 13 It is clear that future studies that examine kidneys of aged dogs need to use the standardized terminology noted above so that cross-study comparisons can be made.
Published Studies Describing the Pathologic Evaluation of Kidneys From Aged Dogs.

Abbreviation: WSAVA-RSSG, World Small Animal Veterinary Association Renal Standardization Study Group.
aVascular lesions not reported.
bVascular lesions not observed.
Distribution of GS and other glomerular lesions have also been reported. In the study of 159 laboratory Beagles, sclerotic glomeruli were more commonly identified near medullary rays and were infrequent in the cortical labyrinth. Immunohistochemistry did not identify increased amounts of type III, IV, or V collagen in glomeruli of older dogs. 57 Two of the 5 control Beagles in the serial renal biopsy study had asymmetrical thickening and splitting of Bowman capsule at the end of the study. 74 One healthy aged Greyhound had synechiae and plasma insudation into the capillary walls in <15% of glomeruli, but these lesions were not present in the other Greyhound. Thickening of the basement membrane of Bowman capsule was identified in less than half of glomeruli in both aged Greyhounds. 13
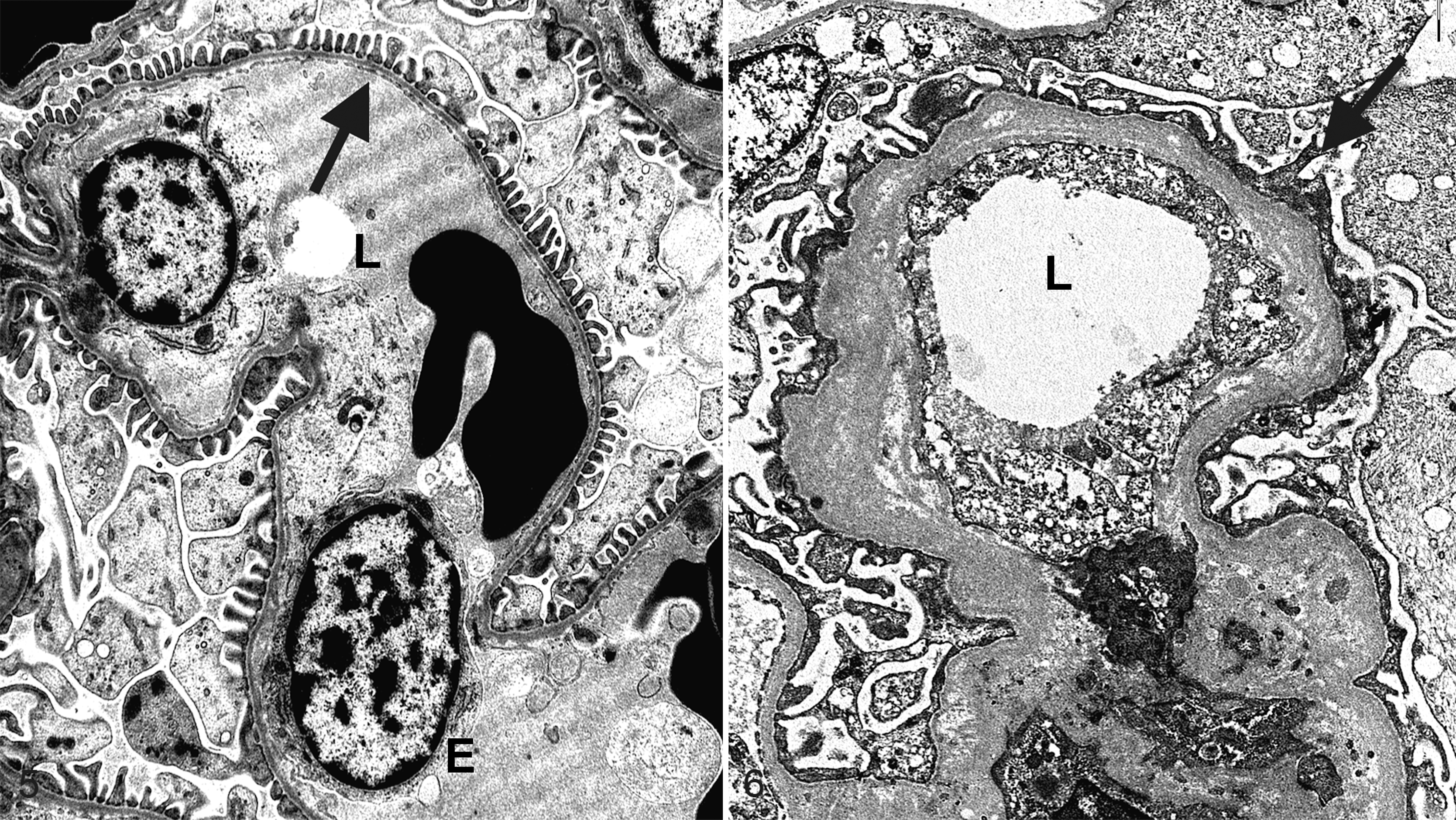
Ultrastructurally, there is an age-related increase in the thickness of the glomerular basement membrane (GBM) in dogs, 45 humans, 60 and rodents. 54 In 1 experimental study, 10 unilaterally nephrectomized 8-year-old dogs were followed for 4 years. Renal tissue from these nephrectomy samples were evaluated with transmission electron microscopy (TEM) and compared with the autopsy specimens from the dogs at 12 years of age. These dogs had a significant increase in mesangial area and GBM thickness in autopsy specimens compared with nephrectomy samples, indicating that the mesangium expands and the GBM becomes thicker with age (Figs. 5, 6). 45 The 2 healthy aged Greyhounds had varying degrees of rarefaction, wrinkling, and multilamination of the GBM, noted on TEM. Podocyte foot processes were segmentally effaced, but electron densities indicative of immune complex deposition were not identified. 13 Even though there are limited studies in which glomeruli from healthy aged dogs have been examined with TEM, it appears that the thickness and ultrastructural appearances of the GBM and mesangium change with age.
GS is also commonly identified in aged humans and rodents. Autopsy data indicate that <10% of all glomeruli should be globally sclerotic in people <40 years old; a higher frequency of global GS is assumed to be disease related. There is a much broader range of globally sclerotic glomeruli in people >40 years old, especially in people >50 years old. 33 One way to evaluate normal renal tissue from living healthy humans is to examine biopsies harvested from donor kidneys immediately posttransplantation, so-called 0-hour biopsies. Studies using these samples have revealed that there is age-associated global GS with secondary compensatory hypertrophy of the nonsclerotic glomeruli even in healthy kidney donors. 76 Glomerular density (number of glomeruli per unit area) can also be used as an index of renal aging. Evaluation of 0-hour biopsies has revealed that glomerular density decreases with age, possibly because nephrons enlarge to compensate for age-related nephron loss; however, if >10% of the glomeruli are globally sclerotic, then glomerular density tends to increase because GS and tubular atrophy are “volume losing” lesions, leading to crowding of the remaining glomeruli. 68 In rats, GS is a component of chronic progressive nephropathy, a disease that is influenced by strain, hormones, and diet. 27 There is age-associated glomerular enlargement in ad libitum–fed 20-week-old Sprague-Dawley rats, followed by GS at 2 years of age. 34
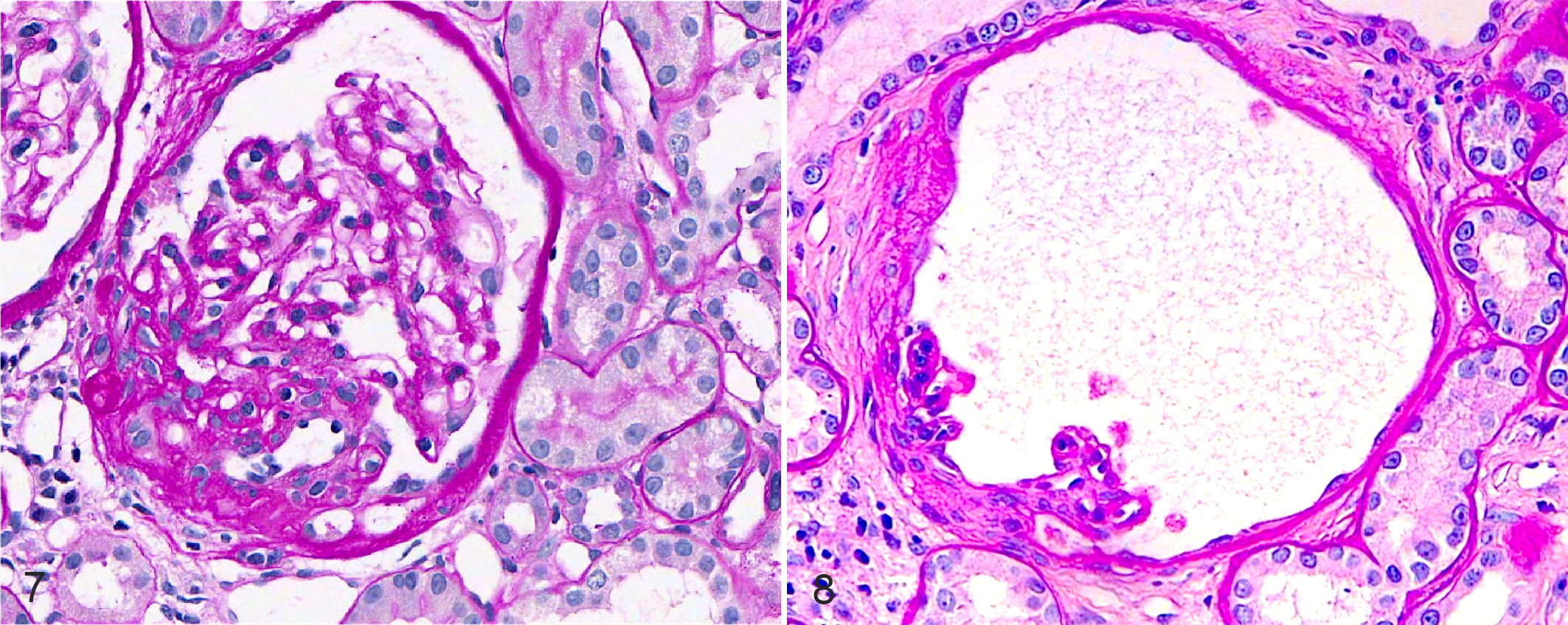
With the recent recognition of focal segmental GS (FSGS) as a disease in dogs, 72 this lesion should be identified and reported separately from global GS (Fig. 7). The WSAVA-RSSG defines FSGS as sclerosis that effaces <50% of a glomerular tuft and involves <50% of glomeruli of the sample. 13 Given that FSGS is associated with proteinuria in humans 22 and dogs, 72 its presence might indicate clinically relevant disease. Notably, FSGS was observed (but not quantified) in the 2 of the canine studies discussed above 29,57 ; however, either it was not mentioned, 11,74 or it was reported as being absent in the others. 13 One additional glomerular lesion, glomerulocystic atrophy (Fig. 8), is characterized by Bowman capsule dilation with concurrent compression of the glomerular tuft. Although many veterinary pathologists consider this an incidental finding, in some scenarios it might be indicative of renal disease, such as glomerulocystic kidney disease, atubular glomeruli, and obstructive nephropathy. 9,12,37
A discussion about the development of FSGS is warranted. Podocyte injury is central to the pathogenesis of this lesion. Podocytes are well-differentiated intricate cells with limited ability to replicate. Although it was once assumed that podocytes could not undergo mitosis, it is possible that in certain disease states (eg, collapsing FSGS), podocytes or their precursors replicate uncontrollably. 15,69 Suffice it to say that in most circumstances, this cell type will hypertrophy rather than divide to cover an enlarged glomerular tuft. In ad libitum–fed Fischer 344 rats, the podocytes hypertrophied, which first appeared to be adaptive, but eventually the enlarged podocytes could not cope with the increased workload. There was overt podocyte loss and eventual FSGS. Interpretation of the stages of podocyte injury was based on decreased expression of normal podocyte proteins, ultrastructural evidence of podocyte foot process effacement, and development of proteinuria. 80 One elegant study developed a transgenic rat model expressing human diphtheria toxin receptor on podocytes and identified a dose-dependent podocyte loss in rats given the toxin. If a rat lost 20% to 40% of its podocytes, it developed FSGS, synechiae, and mild proteinuria. Loss of >40% of podocytes resulted in segmental to global GS, heavy proteinuria, and decreased renal function. 79
Taken together, the literature indicates that a small percentage of global GS, synechiae, and Bowman capsule basement membrane thickening are likely clinically insignificant. If there is compensatory glomerular hypertrophy, FSGS, or frequent glomerulocystic atrophy, then pathogeneses other than age-related remodeling should be considered.
Tubulointerstitial and Vascular Lesions
In the tubulointerstitial compartment, lesions associated with aging have occasionally been reported in dogs, and they consist of well-delineated radiating bands of interstitial fibrosis and tubular atrophy (Fig. 4). The bands of fibrosis might end at the corticomedullary junction and therefore be attributed to ischemia. Alternatively, the tubulointerstitial scarring could extend to the medulla, in which case there is often some degree of hydronephrosis and/or ulceration of the urothelium lining the renal papilla. Inflammation varies greatly. 14
Tubulointerstitial lesions from the previously discussed studies are listed in Table 1. In the study of 159 purpose-bred laboratory Beagles, the severity of renal cortical cysts was increased in older females. The severity of interstitial inflammation and fibrosis also increased with age, but these lesions did not depend on sex. Unfortunately, the scoring schema for these lesions and data regarding neutering were not provided. 57 In the study of 13 aged pet dogs, interstitial inflammation was identified in 1, whereas fibrosis was identified in 4. 29 Serial evaluation of renal biopsies revealed interstitial fibrosis and tubular atrophy in at least 2 time points in 4 of the 5 control Beagle dogs. In the fifth dog, these lesions were identified only once, in the middle of the study. Interstitial inflammation was qualified as being rare in 3 dogs and mild in 2, but all dogs had progressed to having mild to moderate interstitial inflammation at the later time points. 74 In the autopsy evaluation of 145 Portuguese Water Dogs, fibrosis was identified in 24% and interstitial nephritis in 21%. The former but not the latter lesion was significantly correlated with age at death. 11 The 2 aged nonproteinuric, nonazotemic Greyhounds did not have interstitial fibrosis, but 1 had minimal focal interstitial inflammation. 13
Molecular analysis of the renal interstitium revealed an age-associated loss of heparan sulfate in the renal cortices of 10-year-old dogs compared with younger dogs. In the medulla, there was an age-related change in the molecular weight of the hyaluronan, shifting from high to low molecular weight forms. 78 Neither histopathology nor clinical parameters were reported; therefore, the role of renal disease in the changes of extracellular matrix is unknown.
In humans, GS, tubular atrophy, and interstitial fibrosis are components of the diagnostic category of nephrosclerosis. If arteriosclerosis is also present, then the diagnosis is modified to arterionephrosclerosis. In a large study of 0-hour biopsies from adult humans (N = 1203), there was an age-associated increase in nephrosclerosis, wherein <5% of donor kidneys from people aged 18 to 29 years had evidence of nephrosclerosis, compared with 60% of donor kidneys from people aged 60 to 69 years. This age-associated increase in the incidence of nephrosclerosis was not associated with hypertension, decreased glomerular filtration rate (GFR), or increased albuminuria and therefore might be a true age-related lesion. 67
In aged rats, there is an increase in type I collagen but not type III in homogenates of renal cortex. This is not associated with upregulation of TGF-ß transcription; there is, however, a decrease in matrix metalloproteinase 1 (MMP-1) but not MMP-2. 24 Interstitial fibrosis can be identified in 18-month-old rats, and it precedes GS and tubular atrophy. Interestingly, the fibrosis is due mainly to an increase in fibronectin and thrombospondin, with types I and III collagen being limited to regions of tubular atrophy. 3
Much attention has been directed at the rarefaction (loss) of peritubular capillaries as a driving force for tubulointerstitial remodeling in rodents and humans. In 1 study, 24-month-old rats had fewer peritubular capillaries in areas of interstitial fibrosis and tubular atrophy, as demonstrated by decreased immunohistochemical labeling with rat endothelial cell antigen 1. 77 Furthermore, there appears to be crosstalk between the renal tubular epithelial cells and the peritubular capillary. Renal tubular epithelial cells from 24-month-old male Wistar rats with peritubular capillary loss had evidence of mitochondrial dysfunction and associated oxidative stress. Analysis of cultured renal tubular epithelial cells that were depleted of mitochondrial DNA or pretreated with the mitochondrial complex III inhibitor myxothiazol indicated that mitochondrial damage impairs the ability of the cells to upregulate vascular endothelial growth factor in response to hypoxia. The authors suggested that this epithelial cell mitochondrial dysfunction might be an underlying factor in peritubular capillary loss. 70 Notably, peritubular capillary rarefaction has also been documented in humans with chronic kidney disease. 5,38,41
Overall, interstitial fibrosis and tubular atrophy with or without inflammation can be observed in nonazotemic, nonproteinuric aged dogs. The extent and distribution of these lesions might be more clinically relevant than their presence or absence.
Clinicopathologic Changes in Aged Dogs
Many structural changes in the kidney can be clinically silent. A robust study of iohexol clearance in 699 dogs demonstrated an age-associated decline in GFR in small dogs (<15 kg) but not in larger ones. Notably, approximately one-fourth of small dogs with severely decreased GFR had normal serum creatinine, and 62% had a serum creatinine <2 mg/dl. This indicates that small dogs with marked loss of renal function might not have an elevated serum creatinine, likely due to their low muscle mass. 49 Because histopathology was not performed, it is unknown whether the decreased GFR in small breed dogs is due to an increased incidence of renal disease in this population.
Longitudinal data are available from 157 Beagles studied for an average of 13 years at the Laboratory for Energy Related Health at University of California, Davis. Twenty-nine dogs (18.5%) were considered to be azotemic based on persistently elevated serum urea >2 standard deviations from normal. Of these, 8 dogs (5.1%) progressed to uremia and were euthanized. Diagnoses in these uremic dogs included single cases of amyloidosis, membranoproliferative glomerulonephritis, pyelonephritis, obstructive nephropathy secondary to urothelial carcinoma, and acute tubular necrosis due to gentamycin toxicity. The remaining 3 dogs had nephrosclerosis of undetermined etiology. Of the 29 dogs, 19 had azotemia as a feature of extrarenal disease (n = 12) or as a terminal event (n = 7). Nephrosclerosis was identified in 21 of the 29 azotemic dogs, GS in 5, and arteriosclerosis in 3. 14 Another study examined 50 aged purpose-bred laboratory Beagles at the Toxicology Research Institute. Of note, this colony was created with breeding stock from the University of California, Davis, as well as other commercial suppliers. Dogs were divided by age (3, 6, 9, 12, 14 years), and serum chemistry values from each age group were compared. Although all 50 dogs were within the reference ranges for serum urea and creatinine, both parameters were decreased in 6- and 9-year-old compared with 3-year-old dogs. Creatinine was increased in 12-year-old dogs but decreased in 14-year-old dogs. Urea was increased in 14-year-old dogs. 43 Last, a lifetime study of laboratory Beagles from the Center for Environmental Toxicology and Technology at Colorado State University reported autopsy findings for 276 control dogs, 95% of which were >10 years of age. In this cohort, chronic azotemic renal disease was the cause of death in 22 control dogs (8%). This diagnosis was defined by the presence of uremia with “end stage” kidneys. Chronic renal disease contributed to death in an additional 13 dogs, resulting in an overall prevalence of 12.7%. The authors noted that histologic evaluation in all diseased kidneys usually revealed GS, but the underlying pathogenesis of the lesions was rarely elucidated. 7
An investigation of novel urinary biomarkers of renal disease examined 10 aged versus 10 young healthy pet dogs. There was no significant age-related difference in urinary albumin:creatinine ratio, urinary C-reactive protein:creatinine ratio, urinary retinol binding protein:creatinine ratio, and urinary N-acetyl-beta-D-glucosaminidase:creatinine ratio. 75 Age-associated increases in proteinuria have not been reported.
There are abundant data regarding age-associated changes in clinical parameters in humans, with most studies reporting a decline in GFR. 21,66 One large longitudinal study, wherein creatinine clearance was measured 5 to 14 times over the course of many years, demonstrated that creatinine clearance decreased 0.75 ml/min/y in healthy human subjects. Interestingly, however, approximately one-third of the subjects had no detectable decrease over time. 40 Similar studies in rodents have revealed inconsistent results, likely due to variation in strain, sex, 10 and diet.
Taken together, these data indicate that azotemia is uncommon in aged dogs, whereas an age-associated decline in GFR is expected in small breed dogs and many healthy humans. When present, azotemia is often not due to primary renal disease.
Molecular Mechanisms of Renal Aging
Although intense research has focused on the molecular mechanisms underlying aging in the kidney, most of the studies have been in rodents and humans. Gene expression microarray of kidneys from aged humans documented decreased transcription of heat shock protein 70 and insulin-like growth factor 1, with a concurrent upregulation of extracellular matrix genes. 64 Research in rodents has provided further support for the roles of insulin-like growth factor 1 and heat shock proteins. 62,85 GS in humans was associated with upregulation of cell-cycle regulators p16INK4a and p53, whereas tubulointerstitial scarring was associated with p53, TGF-ß1, and p16INK4a expression, among others. 47
One intriguing molecule recently implicated in renal aging is Klotho, a single-pass transmembrane protein that is expressed at high levels in the parathyroid gland, distal convoluted tubule, and choroid plexus. 36 Klotho exists in soluble and membrane-bound forms. In the kidney, it facilitates the bindings of fibroblast growth factor 23 (FGF-23) to its receptors. Together, Klotho and FGF-23 promote phosphaturia and calcium reabsorption. Genetically modified mice with decreases in either Klotho or FGF-23 expression have accelerated aging phenotypes in multiple organ systems, whereas their induction has been protective in experimental models of renal disease. 48 In mice where Klotho expression was eliminated in only the kidney, serum levels of Klotho were decreased to 20% of normal values, underscoring the fact that the kidney is the main source of this molecule. The kidneys of these mice had significantly more interstitial fibrosis, nephrocalcinosis, and dysmorphic tubules when compared with age matched controls. 39 FGF-23 is upregulated in chronic kidney disease in humans 18 and cats 25 but has not been examined in dogs.
The role of the renin-angiotensin-aldosterone system in potentiating renal injury has been recognized. Long-term blockade of the renin angiotensin system with enalapril in old rats prevented age-related GS, 4 mitochondrial dysfunction, and loss of epithelial microvilli in renal tubules. 16 Although the molecular mechanisms are likely diverse, sirtuin 3 is of relevance because it is another longevity gene. Global deletion of the angiotensin type 1 receptor gene increased longevity and led to upregulation of the prosurvival transcript sirtuin 3 in renal tubular epithelial cells. There was decreased oxidative damage in many organs, including the kidneys. Ultrastructural evaluation revealed that proximal tubules had more mitochondria than wild-type mice, but renal tissues were histologically normal. 6 Sirtuin 3 has also been shown to protect renal tubular epithelial cell mitochondria in a model of cisplatin-induced acute kidney injury. 50 We are just beginning to comprehend the relationship between these longevity genes and renal health. More research is needed in all species, but the possibility of therapeutically targeting these pathways to improve renal function is intriguing.
Aside from longevity genes, estrogens and androgens strongly influence age-associated renal changes, wherein estrogens are beneficial and androgens are injurious. Murine podocytes have receptors for both hormones. Estrogen protects against podocyte apoptosis, and testosterone induces it, which can partly explain why male rodents have a higher incidence of GS. 19 Additionally, MMP levels increased with age in female and castrated male rats, but they were not increased in intact males. These MMPs levels were inversely correlated to degree and frequency of GS. 63 The role of estrogens is intriguing because proteinuric dogs diagnosed with FSGS were disproportionately female (74 of 103, 71.8%), almost all of which (93%) were spayed. 72
In aged Fisher 344 rats, the pattern of glomerular gene expression resembles changes observed in arteries with arteriosclerosis. Specifically, there is transcriptional regulation of cell adhesion molecules, chemotactic factors, oxidoreductases (specifically ceruloplasmin), MMPs, coagulation factors, and TGF-ß. These changes were demonstrated to be under the control of NF-kappa B. 82 Interestingly, these studies identified that ceruloplasmin plays a special role in preventing oxidative damage that occurs in aged kidneys. 81 Ceruloplasmin is produced by parietal epithelial cells and can be released in the urine 81 ; therefore, urinary ceruloplasmin might serve as an interesting biomarker indicative of age-related oxidative renal damage. Furthermore, increasing renal ceruloplasmin levels might also be a therapeutic approach.
Beyond the specific molecules just mentioned, three processes of cellular aging have been extensively studied in the kidney. These processes are telomere shortening, cell senescence, and autophagy. Telomere shortening has been documented in the kidneys of aged cats 59 but has not been examined in dogs. Cell senescence develops from cell cycle arrest, and there is abundant evidence that this process commonly occurs in renal tubules. Many atrophic tubules are lined by senescent cells, as demonstrated by ß-galactosidase activity. 17 In this situation, if tubular epithelial cells could be induced to leave a senescent state, it is possible that renal aging could be halted or slowed. Alternatively, induction of senescence of fibroblasts or mesangial cells would be beneficial to prevent extracellular matrix production in scenarios of fibrosis and GS, respectively. 52 Induction of cell senescence in the kidney has been shown to be mediated by mTOR-associated suppression of p21 WAF1/CIP1/SD11 in interstitial cells and mesangial cells. 86
The last major process known to influence renal aging is autophagy, which is responsible for bulk degradation of proteins and organelles. The best-studied mechanism in the kidney is macroautophagy wherein an autophagosome recruits cellular components and then fuses with a lysosome to create a cytoplasmic structure with a double membrane. Macroautophagy is a constitutive process but can be upregulated in certain scenarios of cell stress such that more autophagosomes would be observed ultrastructurally. Moreover, if autophagosome-lysosome fusion is blocked or if there is inhibition of lysosomal enzymes, more autophagosomes would be present. In these conditions, it would appear that autophagy has been upregulated, when in fact it is impeded. Therefore, it can be difficult to discern how autophagy is regulated with only a single time point. Experimental data indicate that defects in autophagy lead to increased degrees of acute tubular epithelial injury, acute kidney injury, GS, and tubulointerstitial scarring with age. 23 Autophagy is especially important in podocyte health because these cells have a constitutively high level of autophagosome flux. In mice with podocyte-specific deletion of autophagy related 5 (Atg5) gene, there was a significant increase in podocyte vacuolation and detachment, mitochondrial injury, and lipofuscin accumulation, leading to an overall increase in the degree of GS and proteinuria. 28 With regard to tubular epithelial cells, autophagy in rodents can be influenced by diet. Mice on an ad libitum diet had less autophagy, and this led to accumulation of swollen, injured mitochondria. Interestingly, mice on a calorie-restricted diet had enhanced autophagy, and this upregulation was mediated by sirtuin 1, another longevity protein. 35
Conclusion
The histologic and ultrastructural features of aging in the canine kidney are similar to those in humans, perhaps with the exception that dogs often lack significant arterial disease. Global GS, interstitial fibrosis, and tubular atrophy are common findings in aged dogs and are part of the pathway to nephrosclerosis. When FSGS is present, it should be identified separately from global GS because it is now considered an entity of glomerular disease. Importantly, future studies of canine kidney disease should use the terminology and classification scheme provided by the WSAVA-RSSG, and renal tissue should be evaluated by light microscopy, TEM, and immunofluorescence.
Clinically, smaller dogs are at risk of having decreased GFR even if the serum creatinine is normal. Other novel urinary biomarkers have not been shown to increase in healthy aged dogs. Many molecules have been identified as being important in the process of renal aging in humans and mice. Investigation of these factors at various levels of genetic sequencing, transcriptional regulation, and protein expression might shed light on the molecular mechanisms of aging. Development of therapeutics that enhance autophagy or induce senescence of fibroblasts might mitigate lesions associated with renal aging. Certainly given the similarities between canine and human renal disease, the translational aspects of any such therapy would be significant.
Footnotes
Acknowledgements
We gratefully thank Davide Trez and Enrico Gallo (Department of Comparative Biomedicine and Food Science, University of Padova) for their expert technical assistance.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.