
Abstract
Systemic amyloid A (AA) amyloidosis is highly prevalent (34%) in endangered island foxes (Urocyon littoralis) and poses a risk to species recovery. Although elevated serum AA (SAA) from prolonged or recurrent inflammation predisposes to AA amyloidosis, additional risk factors are poorly understood. Here we define the severity of glomerular and medullary renal amyloid and identify risk factors for AA amyloidosis in 321 island foxes necropsied from 1987 through 2010. In affected kidneys, amyloid more commonly accumulated in the medullary interstitium than in the glomeruli (98% [n = 78 of 80] vs 56% [n = 45], respectively; P < .0001), and medullary deposition was more commonly severe (19% [n = 20 of 105]) as compared with glomeruli (7% [n = 7]; P = .01). Univariate odds ratios (ORs) of severe renal AA amyloidosis were greater for short- and long-term captive foxes as compared with free-ranging foxes (ORs = 3.2, 3.7, respectively; overall P = .05) and for females as compared with males (OR = 2.9; P = .05). Multivariable logistic regression revealed that independent risk factors for amyloid development were increasing age class (OR = 3.8; P < .0001), San Clemente Island subspecies versus San Nicolas Island subspecies (OR = 5.3; P = .0003), captivity (OR = 5.1; P = .0001), and nephritis (OR = 2.3; P = .01). The increased risk associated with the San Clemente subspecies or captivity suggests roles for genetic as well as exogenous risk factors in the development of AA amyloidosis.
Endangered island foxes (Urocyon littoralis) have lived exclusively on the Southern California Channel Islands 4,55 in genetic isolation for more than 10 000 years. 6,15,51 The foxes evolved into 6 island-specific subspecies with low intra-island genetic variability 15 attributed to historical population bottlenecks. 1 Recent catastrophic population declines due to nonnative predators 7,40 and canine distemper 47 left some islands with as few as 15 individuals. Disease surveillance through postmortem examination revealed a high prevalence of systemic amyloid A (AA) amyloidosis, 11 which poses a threat to species recovery.
AA amyloidosis is the most common systemic amyloidosis in animals and the second most common in humans, and it develops following prolonged or cyclic elevation in the acute phase protein serum AA (SAA). 2,24,43 Transcription of SAA is upregulated in hepatocytes in response to proinflammatory cytokines IL-1, IL-6, and TNF-α. 49 SAA circulates bound to high-density lipoprotein, 2 and serum levels can increase up to 1000-fold in response to infection, inflammation, or tissue damage. 27 Elevated SAA concentration, 42 SAA polymorphisms or mutations, 3,14,30,57 and proteolytic cleavage of SAA into amyloidogenic fragments 53 have been proposed as mechanisms promoting AA amyloid formation.
AA amyloidosis in humans has been associated with chronic infectious diseases, such as tuberculosis and leprosy. 35 Similarly, AA amyloidosis in captive cheetahs is associated with moderate to severe chronic lymphoplasmacytic gastritis 37 from Helicobacter sp infection. 10 Island foxes also have chronic infections, such as the endemic intestinal parasites Mesocestoides 48 and Spirocerca sp, 31,36 the latter causing chronic mural intestinal granulomas in island foxes and elevated serum SAA levels in dogs. 34 Other notable chronic parasitism in island foxes is due to the Otodectes sp ear mite, which is common on the 3 southern islands and is associated with chronic otitis externa. 31
Genetic risk factors for amyloidosis have been identified in humans and animals. In humans with familial Mediterranean fever, an autosomal recessive disease, 60 missense mutations in the pyrin gene are associated with AA amyloidosis. 48 Humans with rheumatoid arthritis have several risk factors for AA amyloidosis, including SAA1.1 (52A) and SAA1.3 (57A) 14,57 genotypes and a single-nucleotide polymorphism (SNP) in the SAA1 promoter flanking region that increases SAA transcription. 29 Similarly, in captive cheetahs, a SNP in the SAA promoter increases SAA transcription under proinflammatory conditions in vitro. 59
Familial AA amyloidosis occurs in Shar Pei dogs 9 and Abyssinian and Siamese cats 19,33,50 but by unknown mechanisms. Highly prevalent AA amyloidosis in inbred captive black-footed cats may be a heritable trait, 46 and concurrent disease and a genetic predisposition are proposed to influence AA amyloidosis development in the captive black-footed ferret population. 12
AA amyloid in island foxes has a widespread tissue distribution, being present in nearly every organ except brain—most commonly, kidney, spleen, and oral cavity. Fox AA amyloid is congophilic, birefringent under polarized light, and immunoreactive against anti-canine AA antibody, and it was identified in high abundance by mass spectrometry in amyloid-affected tissues. 11 Given the high prevalence of AA amyloidosis in genetically and geographically isolated island foxes, here we describe the severity of AA amyloid deposits and assess risk factors for disease.
Materials and Methods
Case Definition and Histologic Analysis
The study population included free-ranging and captive island foxes (N = 321) that died between 1987 and 2010 and were necropsied as part of a disease surveillance and population-monitoring program. Foxes represented all 6 island subspecies: Santa Cruz (Urocyon littoralis santacruzae), Santa Rosa (U. l. santarosea), San Miguel (U. l. littoralis), San Clemente (U. l. clementae), Santa Catalina (U. l. catalinae), and San Nicolas (U. l. dickeyi). Only carcasses with the majority of organs present and without significant autolysis were included; neonates were excluded. Animal history, gross necropsy reports, and histopathology slides were reviewed by a single board-certified veterinary pathologist (P.M.G.) to maintain consistency in the evaluation and severity scoring. Presence of amyloid and all concurrent lesions were determined by examination of formalin-fixed, paraffin-embedded tissue sections stained with hematoxylin and eosin and Congo red to confirm amyloid by green birefringence under polarized light. A case of amyloidosis was defined as a fox with confirmed amyloid in any organ. A semiquantitative grading scale adapted from Terio et al 46 was used to assess severity of amyloid deposition in kidney and spleen as absent (0%), mild (<25%), moderate (25%–50%), or severe (>50%) (Suppl. Table 1).
Immunohistochemistry
Immunohistochemistry for SAA was performed with the avidin-biotin-peroxidase method. Endogenous peroxidase was quenched with 3% hydrogen peroxide. Tissues were then blocked, sequentially incubated in polyclonal rabbit anti-canine AA antibody, 18,54 biotinylated goat anti-rabbit secondary, and streptavidin–horseradish peroxidase; and then visualized with a 3,3′-diaminobenzidine substrate. Positive controls included a Shar Pei dog kidney with amyloid and an island fox kidney with AA amyloid confirmed by mass spectrometry. 11 Additional controls included (1) rabbit IgG isotype substitution of the primary antibody on island fox tissues containing amyloid and (2) primary antibody on island fox tissues lacking congophilic deposits.
Electron Microscopy
Island fox spleen with severe nodular AA amyloid (confirmed by Congo red stain and immunohistochemistry) was fixed in Karnofsky solution, postfixed in osmium tetroxide, embedded in Epon resin, sectioned at 60 nm onto nickel grids, and negatively stained with saturated uranyl acetate in 50% ethanol and bismuth subnitrate solution. Grids were analyzed with a Zeiss EM10 transmission electron microscope (TEM).
Amyloid Fibril Extraction and Ultrastructure
AA-positive spleen tissue from an island fox with severe nodular amyloidosis was stored at –80°C and thawed immediately prior to use. A 55-mg (wet weight) piece was severed and diced with a scalpel, followed by 2 rounds of homogenization in 20mM Tris, 140mM NaCl, 10mM ethylenediaminetetraacetic acid, and 0.1% (w/v) NaN3, pH 8.0, through a Kontes Pellet Pestle (Kimble Life Science and Research Products LLC, Rockwood, TN). The sample was centrifuged at 3000 rpm and 4°C for 5 minutes, and the supernatant was carefully removed and discarded. The pellet was homogenized repeatedly with a Kontes Pellet Pestle in pure water, on ice. 38 At the end of each homogenization step, the sample was centrifuged and the fibril-containing supernatant was carefully removed and analyzed by TEM.
A 5-µl aliquot of the fibril extract was placed onto a carbon-coated Formvar 200-mesh copper grid and incubated for 5 minutes at room temperature. The excess solution was removed by blotting the grid on Whatmann filter paper, and the grids were washed 3 times with 50-µl water droplets and stained 3 times with 50 µl of 2% (w/v) uranyl acetate solution. The grid specimens were air-dried and examined in a JEM-1400Plus (Jeol, Tokyo, Japan) TEM operated at an acceleration voltage of 120 kV.
Risk Factors
Eighteen risk factors were evaluated for an association with AA amyloidosis. Stratifications and grouping of data are as follows: age classes were categorized as young of the year/juvenile, young adult, adult, and geriatric based on known age or tooth eruption and wear pattern of the first upper molar, 56 and age class was evaluated as both a categorical and a continuous variable.
Island subspecies were divided into 4 groups: San Nicolas, Santa Catalina, San Clemente, and northern islands (Santa Cruz, Santa Rosa, San Miguel). The northern island subspecies were grouped due to geographic proximity, shared management practices, and small sample size. Four managing entities were included to assess differing population-monitoring practices affecting carcass recovery and necropsy submission; these entities included the Catalina Island Conservancy, the US Navy, the National Park Service, and North American mainland zoos.
Housing type was categorized as free-ranging or captive. Free-ranging foxes had never been in a captive environment. The captive category was divided into short- and long-term captivity, where “short term” included foxes held in on-island hospitals or holding facilities for ≤30 days, and “long term” included foxes housed in on-island breeding pens or North American mainland zoos for as long as 9 years. Days in captivity were also determined for foxes within the captive strata.
The variable “chronic inflammation” was defined by the diagnosis of one or more of the individually assessed inflammatory diseases as chronic, based on histology at the time of death. Inflammatory diseases included otitis externa, nephritis, Spirocerca granuloma, abscess/cellulitis, pneumonia, septicemia, arthritis, hepatitis, endocarditis, and enteritis.
Proportions of inflammatory diseases were calculated with the denominator reflecting the availability of the organ for examination. Neoplasia included any type of neoplasm present at the time of death. Bilateral adrenal cortical hyperplasia was used as an indicator of chronic stress and was diagnosed microscopically on the basis of a cortex:medulla width ratio greater than 2:1.
Statistical Analysis
Prevalence for AA amyloidosis was calculated for the individual island subspecies; differences between prevalence estimates were determined with a 2-sample z statistic, with P ≤ .05 considered significant. Differences among proportions of mild, moderate, and severely affected kidney and spleen were also determined with a 2-sample z statistic, with P ≤ .05 considered significant.
Demographic and management factors and concurrent diseases were evaluated for their association with amyloidosis based on univariate and multivariable logistic regression using SAS 9.3 statistical software (SAS Institute Inc, Cary, NC). Univariate analyses were used to screen for risk factors to include in the multivariable model and estimate unadjusted odds ratios (ORs) and 95% confidence intervals (CIs). Factors with P ≤ .25 were considered in the multivariable model, 16 which was assembled with stepwise logistic regression and methods of model fitting and assessment of interactions, as described. 16 Residuals and Hosmer-Lemeshow goodness-of-fit test were used to assess model fit. The final model was based on statistical assessments and biological plausibility. The model included adjusted ORs and 95% CIs for cases relative to noncases as a function of the evaluated risk factors, with P ≤ .05 considered significant.
The association between the number of days in captivity—natural log (ln) transformation—and amyloidosis was examined as a continuous variable in the subset of captive foxes. To assess the association between amyloidosis and presence of certain diseases, some data analyses were limited to subsets of island subspecies where the diseases are present. Otitis secondary to Otodectes sp ear mite infection was examined in only the southern island species where this ear mite occurs (Santa Catalina, San Clemente, San Nicolas). Associations with otitis and ceruminous gland neoplasia were examined in the Santa Catalina subspecies, in which otitis is the most severe and ceruminous neoplasms were exclusively found.
The amyloid-containing kidney was chosen to assess the association between risk factors and amyloid severity because kidney is the most commonly affected organ resulting in clinical disease and mortality. The highest grade of renal amyloid—glomerular or medullary—was used in the analysis. Due to small sample size, the association between risk factors and severe amyloid deposition in kidney was examined with exact univariate logistic regression.
Results
Gross Lesions and Ultrastructure of AA Amyloid
We characterized the macroscopic lesions in 109 cases of amyloidosis in necropsied island foxes. The kidney, oral cavity, and spleen most commonly had gross lesions. In severely affected cases, the kidneys appeared diffusely pale and waxy (Fig. 1) with subcapsular stippling and white medullary streaks on the cut surface. In the oral cavity, there was macroglossia (up to 2 times normal size), often with mucosal hyperplasia of glossal papillae and submucosal amyloid nodules up to 8 mm. Within the epiglottis, there were 1- to 3-mm nodules of amyloid that distorted the normal architecture (Fig. 2). The spleen was diffusely enlarged (2 to 4 times normal size) or contained macroscopically visible white nodules of amyloid (up to 7 mm; Fig. 3).
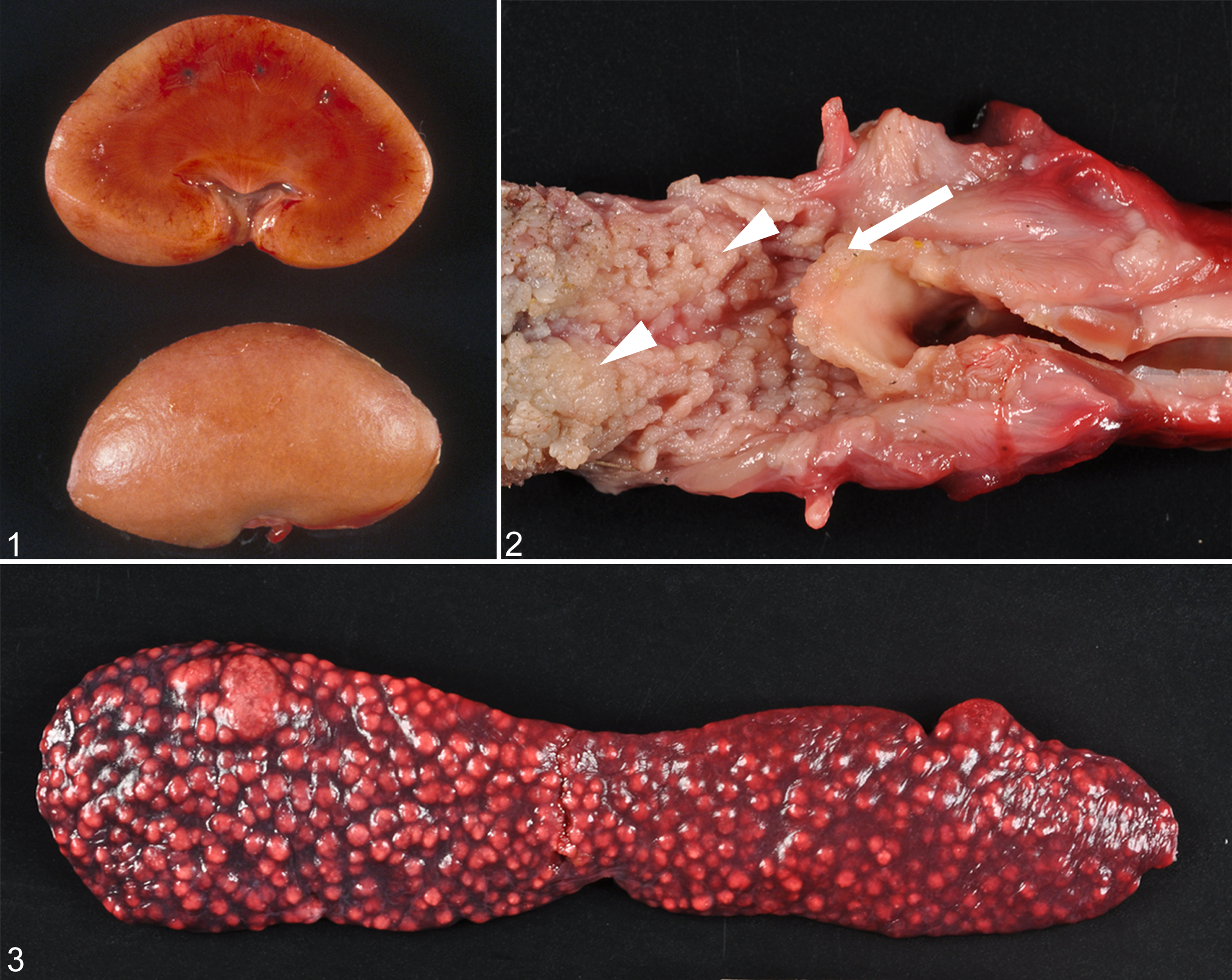
Appearance of amyloid A amyloidosis in island fox organs.
Since nodular splenic AA amyloid is unusual, amyloid fibrils were examined ultrastructurally by TEM, both in situ and in splenic extracts. Severe nodular splenic amyloidosis (Fig. 4a) was confirmed as AA by immunoreactivity to anti-canine AA antibody (Fig. 4b). In situ, nodules of splenic amyloid were composed of densely packed mats of haphazardly arranged nonbranching fibrils of approximately 10-nm diameter (Fig. 5). The extracted fibrils also possessed a linear and nonbranching structure, however, lacked high regularity and did not exhibit a uniform fibril width when measured at different positions along the same fibril (Fig. 6). Instead, fibrils often appeared broken or splintered and possessed multiple aligned protofilaments. Consequently, the fibril width differed considerably from 10 nm to almost 50 nm. Only some fibrils exhibited a well-resolved crossover structure (constriction at the site of fibril twists; Fig. 6).
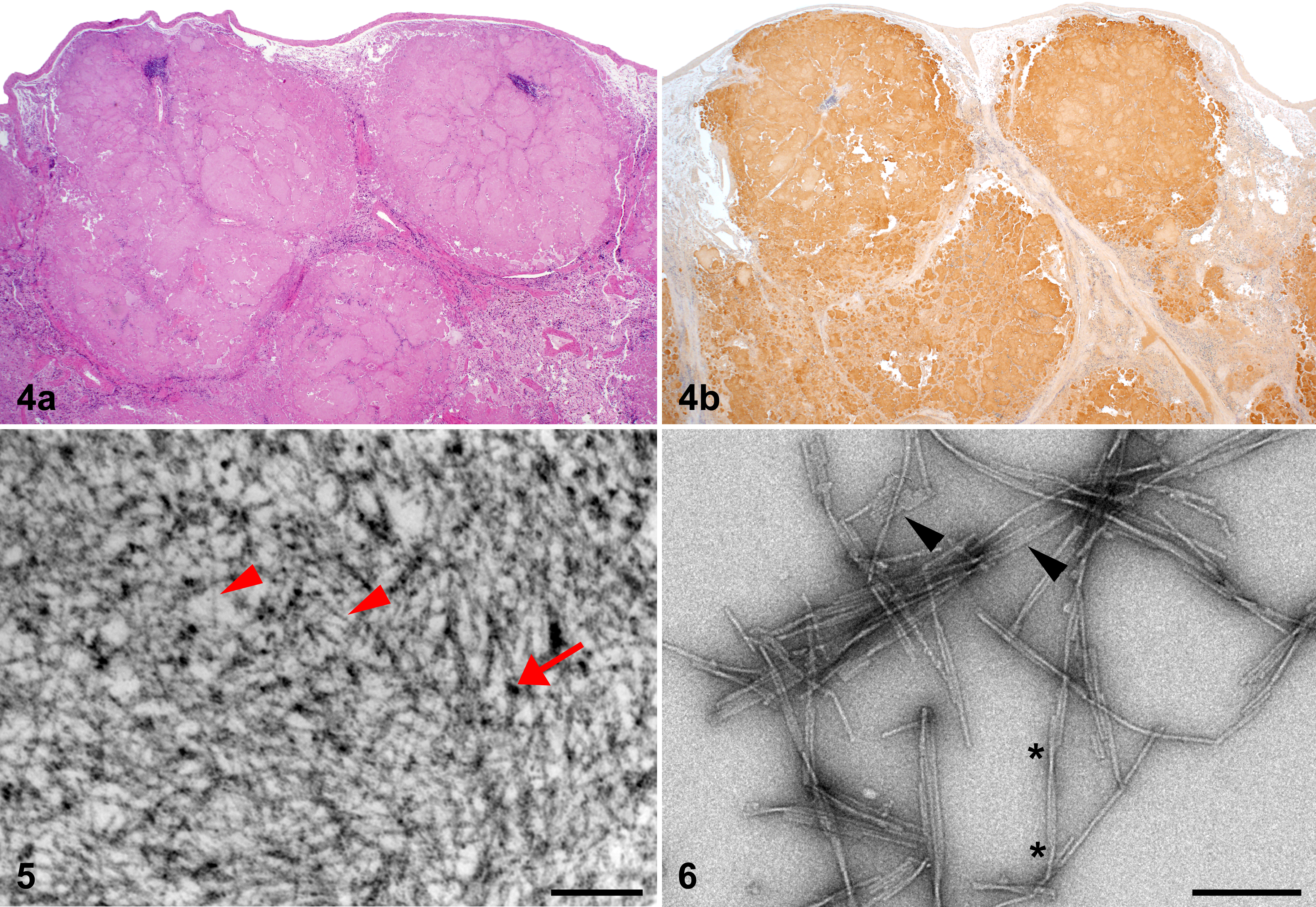
Microscopic and ultrastructural appearance of severe nodular amyloidosis in the spleen of island fox.
Severity of Renal and Splenic Amyloid
In cases with renal amyloidosis (n = 80), medullary amyloid was more common than glomerular amyloid (98% [n = 78 of 80] and 56% [n = 45], respectively; P < .0001). Because renal amyloid can lead to kidney failure, we characterized the severity of renal AA amyloid in all amyloidosis cases with available kidney (n = 105). The degree of amyloid deposition in glomeruli and medulla was designated as absent, mild, moderate, or severe, reflecting 0%, <25%, 25%–50%, or >50% amyloid deposition (Suppl. Table 1). Amyloid was absent from glomeruli and medulla in 57% and 26% of kidneys, respectively. Glomerular amyloid deposits were most often mild (30%); only 7% of amyloid cases had severe deposition. Similarly, medullary amyloid deposits were most often mild (43%) and were less frequently moderate (12%) or severe (19%; Figs. 7–16).
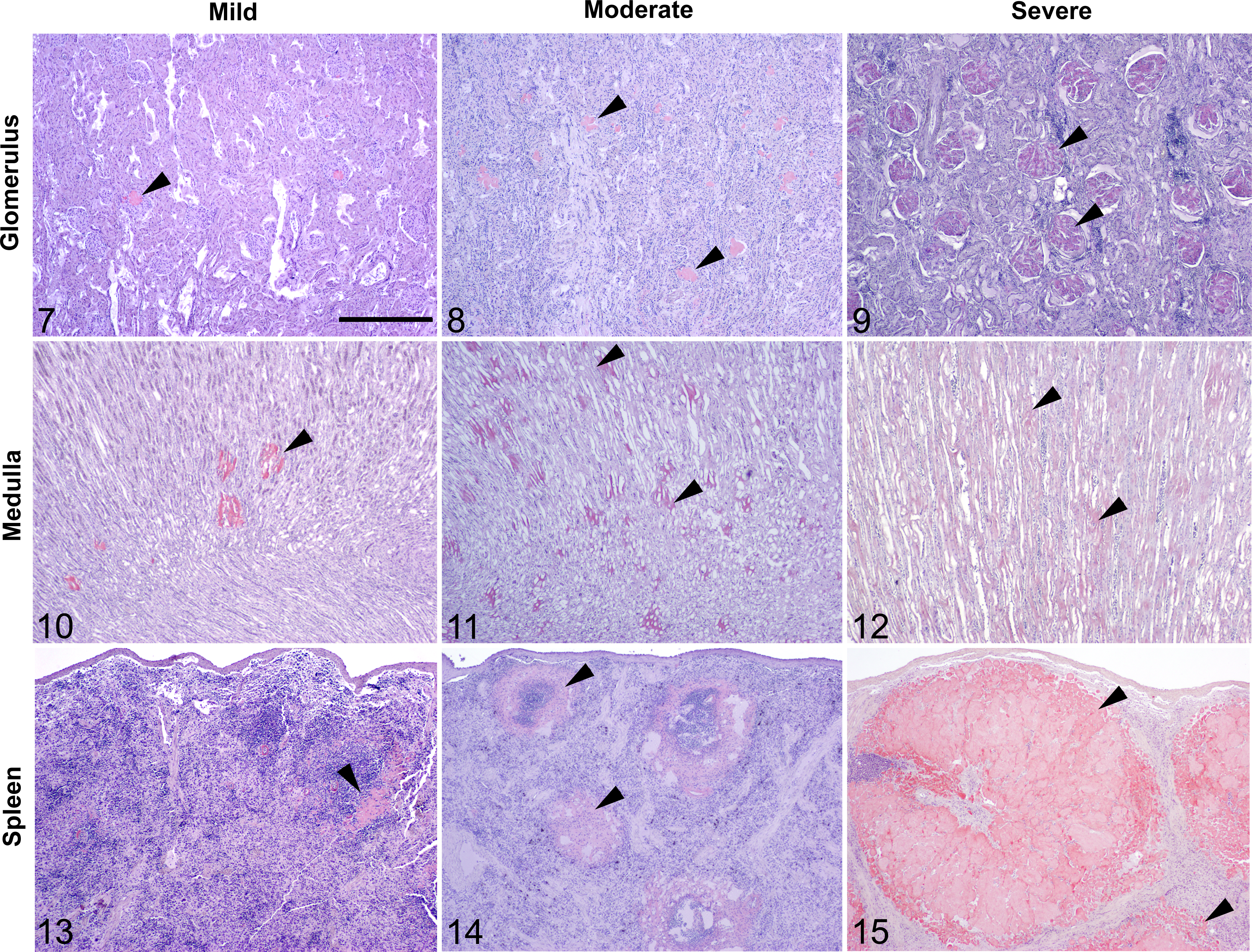
Mild, moderate, and severe amyloid A amyloidosis (black arrowheads), kidney and spleen, island fox. Scale bar = 1.0 mm. Congo red.
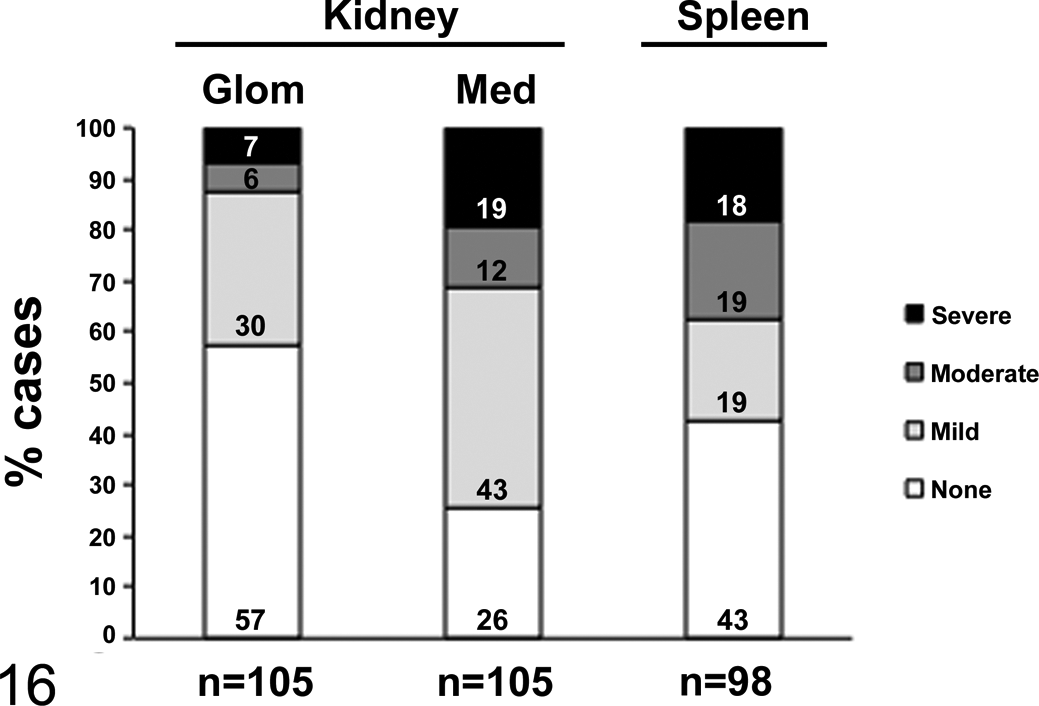
Proportions of cases with varying degrees of amyloid A amyloid deposition. When present, renal amyloid was most commonly mild for both the glomeruli (Glom) and the renal medulla (Med). Mild, moderate, and severe amyloidosis occurred in nearly equal proportions in the spleen. Proportions were calculated out of total organs available for examination in cases of amyloidosis.
In amyloid cases with kidney available for examination, renal amyloidosis caused or contributed to death in 18% of foxes (n = 19 of 105). This may underrepresent the mortality due to renal amyloidosis, as many of these amyloid cases (n = 39) died prematurely from vehicular trauma. Among nontrauma-related deaths, 29% (n = 19 of 66) of amyloid cases died from renal amyloidosis.
In the spleen (n = 98), the proportions of mild (19%), moderate (19%), and severe (18%) amyloid deposition were nearly equal, while the remaining cases (43%) had no splenic amyloid (Figs. 13–16).
Prevalence and Risk Factors for AA Amyloidosis
Prevalence of AA Amyloidosis in Island Fox Subspecies
To estimate prevalence, we examined the necropsied population of island foxes (N = 321) that died between 1987 and 2010 and were juveniles or older. The overall prevalence of amyloidosis in this population was previously estimated at 34% (n = 109 of 321). 11 Prevalence among each island subspecies was further evaluated in the present study and ranged from approximately 23% to 50%: San Nicolas (22.6%), Santa Catalina (29.5%), Santa Cruz (33.3%), Santa Rosa (36.4%), San Clemente (38.9%), and San Miguel (50.0%; Table 1). The prevalence of amyloidosis in the San Clemente subspecies was significantly higher than in the San Nicolas subspecies (P = .02). Prevalence estimates were not significantly different among the other island subspecies or between the southern or northern island groups.
Estimated Prevalence of Amyloid A Amyloidosis in Necropsied Island Foxes (N = 321).
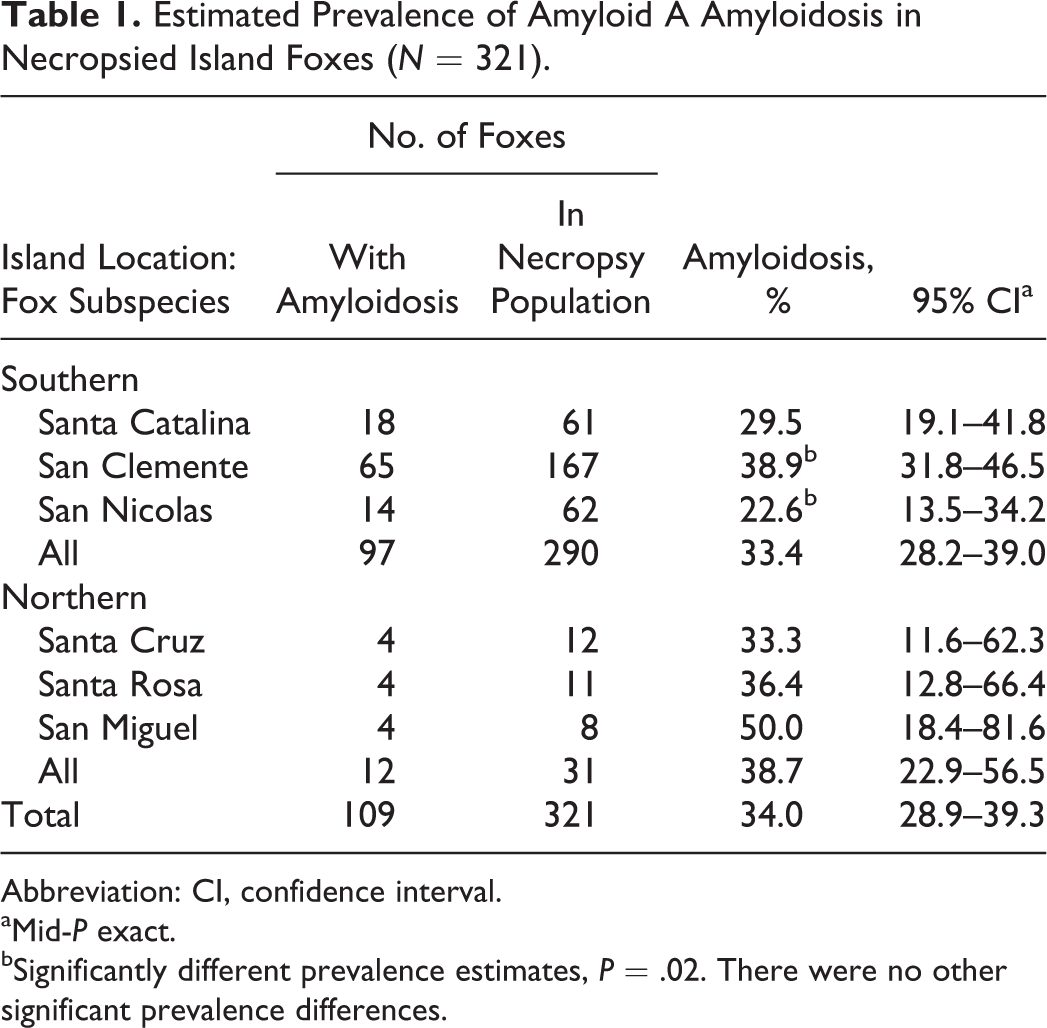
Abbreviation: CI, confidence interval.
aMid-P exact.
bSignificantly different prevalence estimates, P = .02. There were no other significant prevalence differences.
Most foxes examined were free-ranging (n = 253); fewer were housed for any time in captivity (n = 66). Prevalence of AA amyloidosis among foxes housed in captivity was significantly higher when compared with the free-ranging population (59% [n = 39 of 66] vs 27% [n = 69 of 253], respectively; P < .0001).
Concurrent Inflammatory Diseases
Comorbid inflammatory lesions at the time of death included chronic otitis externa associated with Otodectes sp ear mite infection, which was present in the majority of cases (88.8%) from the southern islands. Nephritis—including glomerulonephritis, pyelonephritis, interstitial nephritis, and pyelitis—was the next-most common group of lesions (39.6%), followed by mural intestinal or mesenteric granulomas due to Spirocerca sp infection (24.0%) and trauma-associated abscessation and cellulitis (20.2%). Less common concurrent inflammatory lesions were pneumonia (14.6%), sepsis (5.6%), arthritis (5.5%), hepatitis (5.0%), endocarditis (3.9%), and enteritis (3.9%). Animals were considered to have chronic inflammation if they had histologic evidence of chronic, active inflammation at the time of death, which was 83.5% of the amyloid cases and 90.6% of noncases (Table 2).
Univariate Logistic Regression Analyses of the Relationship Between Amyloid A Amyloidosis in Island Foxes and Evaluated Variables (N = 321).
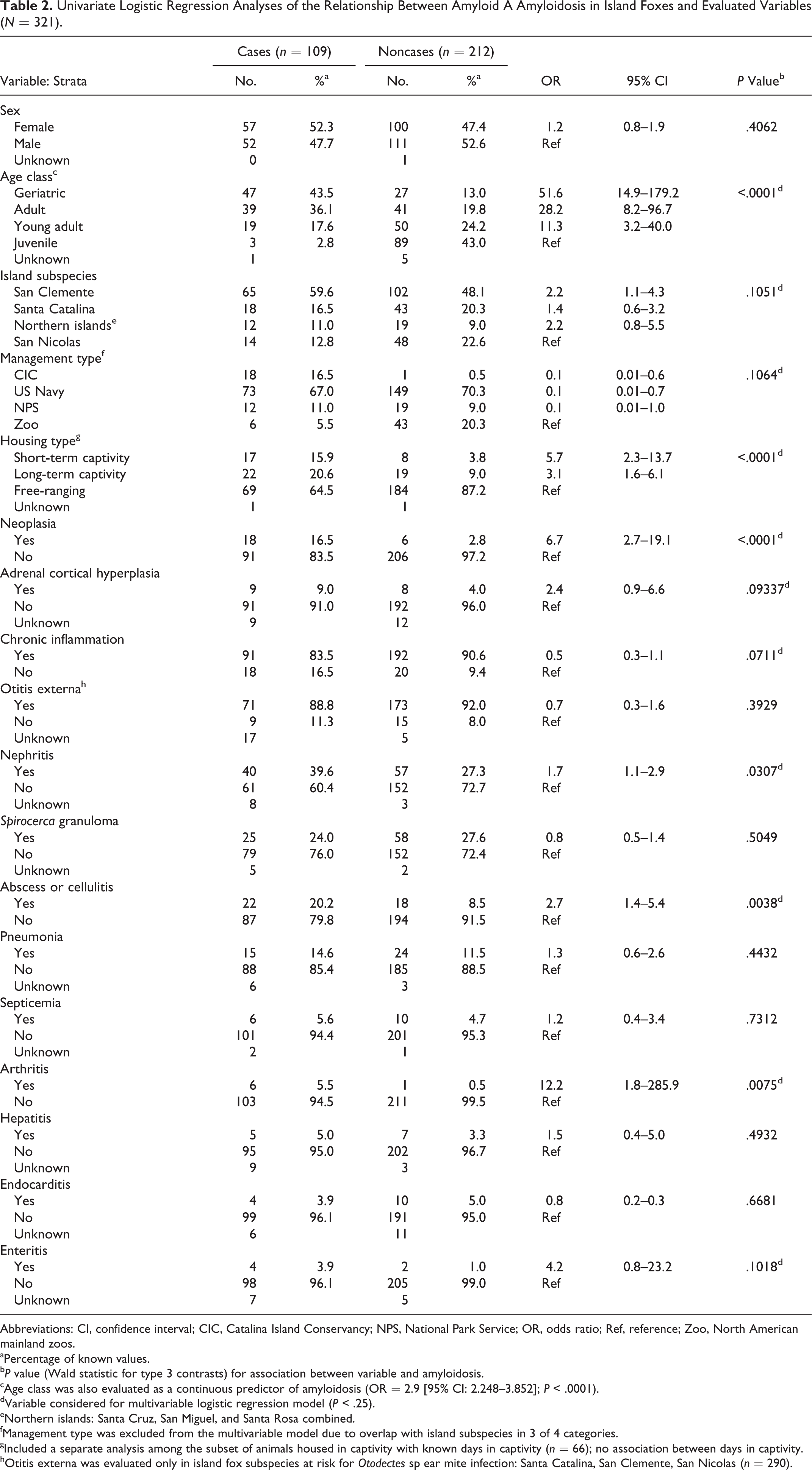
Abbreviations: CI, confidence interval; CIC, Catalina Island Conservancy; NPS, National Park Service; OR, odds ratio; Ref, reference; Zoo, North American mainland zoos.
aPercentage of known values.
b P value (Wald statistic for type 3 contrasts) for association between variable and amyloidosis.
cAge class was also evaluated as a continuous predictor of amyloidosis (OR = 2.9 [95% CI: 2.248–3.852]; P < .0001).
dVariable considered for multivariable logistic regression model (P < .25).
eNorthern islands: Santa Cruz, San Miguel, and Santa Rosa combined.
fManagement type was excluded from the multivariable model due to overlap with island subspecies in 3 of 4 categories.
gIncluded a separate analysis among the subset of animals housed in captivity with known days in captivity (n = 66); no association between days in captivity.
hOtitis externa was evaluated only in island fox subspecies at risk for Otodectes sp ear mite infection: Santa Catalina, San Clemente, San Nicolas (n = 290).
Risk Factors for AA Amyloidosis
To identify risk factors for amyloidosis in the island foxes, 18 variables were assessed for their association with the presence of AA amyloid (Table 2). Eleven variables met the criteria for consideration in the multivariable analysis (P < .25), including age class, island subspecies, island management type, housing type, neoplasia, adrenal cortical hyperplasia, chronic inflammation, and specific inflammatory diseases (otitis, nephritis, abscesses/cellulitis, arthritis, and enteritis). Based on associations in the univariate analysis and to improve statistical efficiency, the final model included age class as a continuous predictor, and short- and long-term captivity variables were combined into a single category, denoted captive. The final adjusted model included 4 independently significant variables: age class, island subspecies, captivity, and nephritis (Table 3). There was no evidence of lack of fit (Hosmer-Lemeshow goodness of fit, P = .47) nor identified confounders or interactions.
Multivariable Logistic Regression Model for the Association Between Amyloid A Amyloidosis in Island Foxes and Identified Risk Factors (n = 305).a
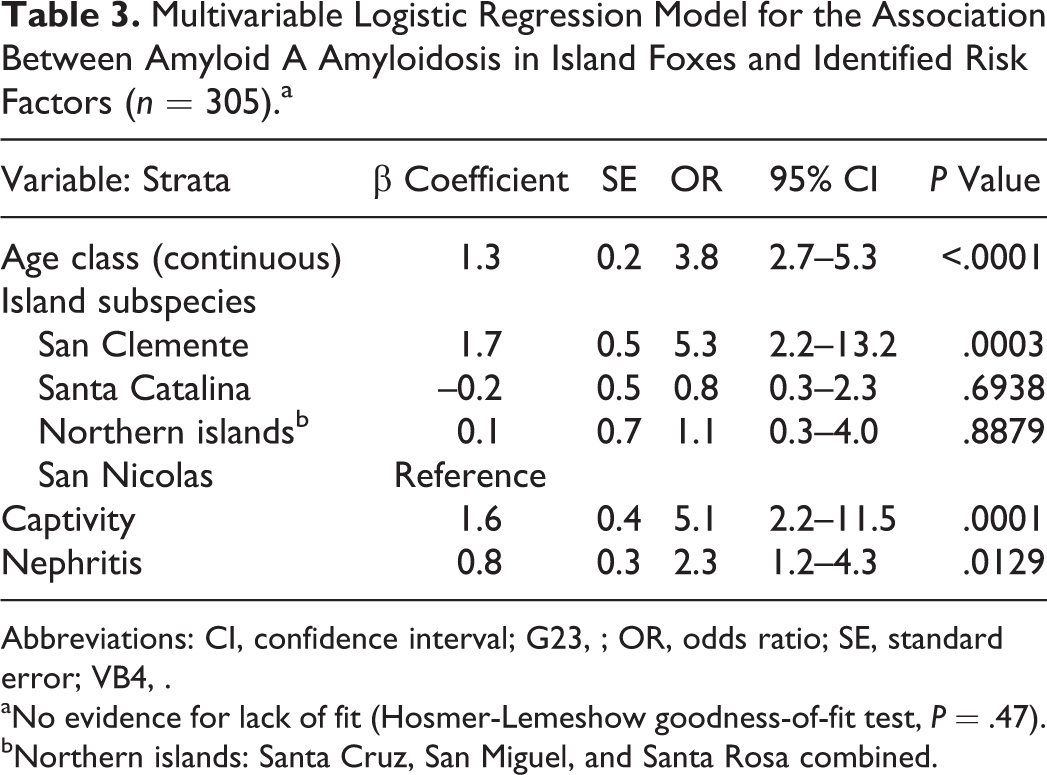
Abbreviations: CI, confidence interval; G23, ; OR, odds ratio; SE, standard error; VB4, .
aNo evidence for lack of fit (Hosmer-Lemeshow goodness-of-fit test, P = .47).
bNorthern islands: Santa Cruz, San Miguel, and Santa Rosa combined.
Our regression model showed that successively increasing age classes were associated with amyloidosis (P < .0001). The odds of disease increased 3.8-fold for each increasing age class. Adjusting for age and other variables in the model, the San Clemente Island fox subspecies had 5.3 times higher odds of having amyloidosis (P = .0003) when compared with the San Nicolas subspecies. Foxes housed in captivity were 5.1 times more likely to have amyloidosis than free-ranging foxes (P = .0001), however, the risk did not increase with longer time in captivity (P = .50) in an adjusted analyses among the captive subset. The presence of any nephritis (pyelonephritis, interstitial nephritis, glomerulonephritis) was significantly associated with amyloidosis (OR = 2.3; P = .01). Whether nephritis preceded or was a sequela of amyloidosis is not known. Because previous work showed that nephritis occurs in dogs with renal amyloidosis 41 and cheetahs with chronic renal disease have elevated circulating SAA, 8 we examined the relationship between nephritis and renal amyloidosis and found no association in island foxes (P = .3).
An association between neoplasia and AA amyloidosis was initially identified in the univariate analysis but was explained by the increased occurrence of neoplasia with increasing age and was no longer significant in the adjusted model.
Risk Factors for Severe Renal AA Amyloidosis
Among the cases of renal amyloidosis (n = 80), the same risk factors were assessed for an association with severe renal amyloidosis by exact univariate logistic regression, which revealed that female foxes and foxes in short- and long-term captivity had significantly higher odds of severe renal amyloid when compared with males and free-ranging foxes (ORs = 2.9, 3.2, 3.7, respectively; all P values < .05; Suppl. Table 2). Management type was also significantly associated with severe renal amyloidosis (P = .03) when zoo foxes were compared with those managed by the Catalina Island Conservancy. Small sample size did not allow for further assessment of this association; however, severe renal amyloid in zoo foxes may be due to longer life in captivity with clinical care. No other variables had a significant association with severe renal amyloid. Adjusted multivariable analyses were not performed owing to the small sample size and lack of statistical power.
Island-Specific Risk Factors for AA Amyloidosis
Otodectes sp–associated chronic otitis externa occurred only in the southern island foxes (Santa Catalina, San Clemente, San Nicolas) but was not associated with amyloidosis (P = .3). Ceruminous gland tumors were identified only in Santa Catalina foxes and were not significantly associated with amyloidosis (P = .3).
Discussion
The island fox is an endangered species with a high prevalence of AA amyloidosis. Here we define the degree of amyloid deposition in the most commonly affected organs—kidney and spleen—and identify the risk factors for disease. Independent risk factors for AA amyloidosis were age class, San Clemente Island subspecies, captivity, and nephritis.
In island foxes, the kidney most commonly contains AA amyloid, 11 similar to humans and many animal species with systemic AA amyloidosis. 20,28,37,39,41,46 Here we found that both glomerular and medullary amyloid deposits were common in affected fox kidneys, however, medullary deposits were nearly universal, similar to Shar Pei dogs, 9,41 Abyssinian cats, 19,33 and captive cheetahs 37 and black-footed cats. 46
The severity of renal amyloid deposition in island foxes was typically mild. In contrast, renal AA amyloidosis is typically severe in captive cheetahs, black-footed cats, and black-footed ferrets, which also have a high prevalence of systemic AA amyloidosis. 12,37,46 This difference is most likely attributable to most island foxes being free-ranging and subject to loss at younger ages from predation and vehicular trauma and lack of medical treatment. Captive island foxes were more likely to have severe renal amyloidosis.
Prolonged or cyclic elevation of circulating SAA typically precedes the deposition of AA amyloid. Chronic elevations in SAA are secondary to infectious, immune-mediated, neoplastic, and familial disease. Most island foxes died with evidence of a chronic and ongoing inflammatory disease, Otodectes sp or Spirocerca sp infection, or a tumor. Other than nephritis, there was no significant association between an acute or chronic inflammatory disease and amyloidosis when controlling for other demographic or management factors. This finding was not surprising given that 90.6% of amyloid-negative foxes also had evidence of chronic inflammation. While inflammation likely predisposes to AA amyloidosis and whereas nephritis was significantly associated with disease, additional endogenous or environmental cofactors may contribute to the high prevalence of AA amyloidosis in island foxes.
A genetic or exogenous factor may promote the development of AA amyloidosis in island foxes, particularly in consideration of the 5.3-fold increased odds of amyloidosis for the San Clemente versus San Nicolas subspecies and the absence of reported amyloidosis in mainland gray foxes, the island fox’s closest relative. 21 Possible genetic associations with amyloidosis could be due to (1) a mutation in the SAA gene(s) resulting in a highly amyloidogenic SAA sequence or (2) a mutation in the promoter, enhancing elements, or transcription factors resulting in elevated or prolonged SAA transcription. The amyloidogenic SAA1.1 isoform in humans and island fox SAA both have a high predicted aggregation potential at amino acid segment 2–7. 11,25 An SAA promoter mutation is also a possibility, as described in the cheetah. 59
Captivity was a significant risk factor for AA amyloidosis in island foxes, independent of age and subspecies, however, increased days in captivity were not associated with any increase in risk. This suggests that some aspect of the initial exposure to the captive environment may account for the increased risk for disease. One possible link between exposure to a captive environment and AA amyloidosis is the potential for transmission of AA amyloid by a prion-like mechanism through a seeding-nucleation process. 5,32 A captive environment contaminated with AA amyloid fibrils could expose an animal to an AA amyloid seed. In mice injected with a proinflammatory stimulus, such as silver nitrate, AA amyloid deposition is accelerated by inoculation of amyloid fibrils (amyloid-enhancing factor) isolated from amyloidotic organs 23,26 or from cheetah feces. 58 The potential for island fox AA amyloid to transmit horizontally from animal to animal should be considered.
Another possible explanation for captivity as a risk factor is the stress-induced production of glucocorticoids, which are known to enhance IL-1- and IL-6-induced SAA transcription. 17,44,49 Elevated fecal cortisol levels have been shown in captive versus wild cheetahs and in on- versus off-exhibit cheetahs. 45,52 Additionally, the social stress of crowded housing is associated with AA amyloidosis in Syrian hamsters and mice in the laboratory setting. 13,22 Thus, captive foxes may experience enhanced SAA transcription from stress-induced glucocorticoid production that could promote amyloidosis.
The risk factors for amyloidosis suggest that genetic and exogenous factors promote disease; however, the underlying mechanisms remain unclear. Further investigation of the molecular pathogenesis of amyloidosis is warranted to determine the role of genetic mutations, epigenetic changes, and environmental influences on AA amyloidosis development.
Footnotes
Acknowledgements
We thank the members of the island fox recovery team for more than 15 years of sample collection and valuable consultation: Tim Coonan, Winston Vickers, Dave Garcelon, Julie King, Calvin Duncan, Angel Guglielmino, Christina Boser, Brian Hudgens, Melissa Booker, Grace Smith, Mark Willet, Julie Barnes, and Karl Hill.
We gratefully acknowledge the institutions that support island fox recovery efforts: the National Park Service, the Nature Conservancy, the US Navy, the Catalina Island Conservancy, the Friends of the Island Fox, the California Department of Fish and Wildlife, and the US Fish and Wildlife Service. We thank Drs Walter Boyce and Peter Ernst for valuable discussion.
We thank Jaime Rudd and Carlitos Chen for excellent technical assistance.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by an Academic Senate Grant from the University of California, San Diego, the Karen C. Drayer Wildlife Health Center, School of Veterinary Medicine, University of California, Davis, and the ARCS Foundation of Northern California. P.M.G was funded by The Linda Munson Fellowship for Wildlife Pathology Research administered by the ACVP/STP Coalition for Veterinary Pathology Fellows and the Peter Kennedy Fellowship in Anatomic Pathology, School of Veterinary Medicine, University of California, Davis. A.K.’s work was supported through the GERAMY consortium (Bundesministerium für Bildung und Forschung, BMBF). The research of M.F is funded by the Deutsche Forschungsgemeinschaft (FA 456/15-1).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.